
Abstract
Spontaneous settling of certain aggregating microalgae or cyanobacteria could potentially alleviate the energy requirement in biomass harvesting. A locally isolated floc–forming marine cyanobacteria, Chroococcidiopsis sp., was initially grown indoor to study its self-settling efficiency; 97% of the biomass settled spontaneously in 1 h. Later, the strain was grown in outdoor raceway tanks (1 square meter) for 70 d. Every alternative day, 50% of the culture was harvested by self-settling, and the supernatant was recycled back to the tank. Average biomass productivities for the strain was 16.08 g/m2/d. The biomass had an average 2.26% water extractable phycobiliproteins that comprised of both phycocyanin (11.4 mg/g) and phycoerythrin (10.6 mg/g). Since Chroococcidiopsis sp. biomass can be harvested very efficiently, it would reduce the energy and cost of biomass production which are deemed necessary for microalgal animal feed and biofuel applications. Furthermore, its ability to produce high-value pigments will also make it economically very attractive from biorefinery approach.
Introduction
Microalgal biomass is considered one of the potential sustainable feedstocks for biofuel, feed, and bulk chemicals because microalgae have very high areal biomass productivity while utilizing non-arable land and non-potable water. 1 –3 Depending on the strain and metabolites of interest, microalgae can be cultivated in batch, semi-continuous, or even continuous mode. Batch mode of cultivation is often preferred for starving some of the microalgae to accumulate high amount of lipid. 4 However, for low-value feedstocks, in most other cases, semi-continuous and continuous cultivation are adopted. Furthermore, lipid-rich microalgae biomass is not essential for biocrude oil production using hydrothermal liquefaction (HTL), as other major metabolites (i.e., proteins and carbohydrates) also get converted into biocrude in HTL process. 5,6 Similarly, protein-rich microalgae/cyanobacteria biomass (e.g., Spirulina sp.) can be used as fish-feed supplement. 7
After harvesting the biomass, the culture growth media is usually recycled back into the cultivation system; this not only minimizes the water footprint but also allows recycling of the excess nutrients in the media 8,9 However, depending on the harvesting method, the recycled media can introduce additional unwanted compounds in the culture; for example, coagulation-flocculation, and electrocoagulation will increase the concentration of specific metals whereas, centrifugation can release intracellular organic matter. 10,11 Additionally, microalgae biomass harvesting can be very energy demanding. 12,13 To overcome these issues, auto-flocculating or self-settling microalgae or cyanobacteria strains could be used. Adjusting the culture pH, either by acid or base, were also studied to initiate settling for several microalgae. However, the requirement of chemicals in both adjusting the culture pH for inducing the flocculation, and to adjust the culture pH after flocculation will incur additional cost. On the contrary, few microalgae (green unicellular and diatom) and cyanobacteria precipitate spontaneously, in the absence of any external mixing.
Chroococcidiopsis sp. is a thermophilic, desiccation-tolerant, and ionization-radiation tolerant cyanobacteria. 14 Therefore, the growth of this strain was studied under extremely harsh environmental conditions similar to Mars. 15 The strain Chroococcidiopsis sp. has the ability to produce exo-polysaccharides which initiate the formation of cubical cell aggregates; once the cell aggregates become heavy, these undergo gravity sedimentation (or, self-settling). 16 Cyanobacteria biomass could be a rich source of essential amino acids and pigments for feed application. Phycocyanin (PC) and Phycoerythrin (PE) are two of the water extractable phycobiliproteins that are found in some cyanobacteria and microalgae. 16,17 The presence of high-value metabolites could reduce the cost and energy burden of the biomass production, especially for biofuel and bulk chemical feedstocks. In an indoor experiment, Hayashi et al. showed that Chroococcidiopsis sp. was capable of producing a high concentration of PC. 18 Considering its ability to grow in extreme conditions, accumulate high-value pigments, and precipitate spontaneously, this strain could be grown in large scale for commercial use. There are limited number of indoor small-scale growth studies on this microorganism, and hence, the objective of this study was to investigate its long-term outdoor growth performance under the dynamic environment.
Materials and Methods
Strain and the Growth Medium
Chroococcidiopsis sp. is a self settling marine cyanobacteria which was isolated from Qatar's environment. 19 Modified Guillard f/2 nutrients media was used to grow this cyanobacteria; urea was used as a nitrogen source to replace sodium nitrate. The strain was grown in natural seawater (4.2% NaCl) which was collected from AL-Khor shore area, and transported by a tanker. For indoor cultivation, the seawater was initially filtered through a 0.45 μm filter paper under vacuum; later the filtered seawater was autoclaved before inoculation. For outdoor largescale cultivation, seawater was sterilized using commercial sodium hypochlorite.
Settling Study of Chroococcidiopsis sp.
Self-settling study of Chroococcidiopsis sp. was conducted in a 1-L glass container. 1 L dense culture of Chroococcidiopsis sp. (around 1.0 g/L biomass density) was poured into the container and 1 mL sample was collected, 5 cm above the bottom of the container, at different time intervals (0–60 minutes). Biomass density of the sample was measured by taking its optical density (OD) at 750nm. The self-settling efficiency of the strain was calculated using the following formula:
where AFE = self-settling efficiency at time t; Wi = Initial biomass density (g/L); Wt = Biomass density (g/L) at time t.
Outdoor Culture Condition and Harvesting
Chroococcidiopsis sp. inoculums for the outdoor cultivation was prepared indoor in 10-L plastic photobioreactors. 20 L indoor-grown culture was inoculated into 180 L of sterilized seawater in a 1-m2 raceway tank. Two identical raceway tanks were used as replicates for the growth trials. 100% pure CO2 was sparged at the bottom of the raceway tanks. pH value of the culture was checked multiple times in a day to ensure that the culture pH remained in between 7.5–8.5. A modified Guillard f/2 nutrients were used to grow this strain (Table S1, Supplementary Data are available online at
Metabolite Analysis
Six harvested biomass samples were selected, from each raceway tank, for understanding the variation in intracellular metabolites production. At first, the phycobiliproteins were extracted using a phosphate buffer (pH = 7.2). Next, the supernatant was separated using centrifuge and concentrations of phycocyanin and phycoerythrin, in the extracted liquid sample, were determined using empirical equations (Equations 2 and 3) as developed by Sampath-Wiley and Neefus.
20
A564, A618, and A730 are the absorbance values at 564 nm, 618 nm, and 730 nm, respectively.
Cellular total lipid content was estimated by a colorimetric sulfo-phospho-vanillin method. 21 Intracellular protein, and carbohydrate contents were determined by Lowry and Phenol-Sulfuric methods respectively; the details of these methods were presented elsewhere. 22 Amino acid profile of Chroococcidiopsis sp. biomass was determined (by Eurofins-India Limited) using liquid chromatography and fluorescence detector.
Bioassay with Chroococcidiopsis sp. Biomass
Every alternative day, 1 mL culture from the tank was added in a 1 L artemia culture, which continued until the end of the outdoor growth experiment. Chroococcidiopsis sp. biomass was mixed with grinded commercial fish feed (sera gold) at a ratio 1:4, and the necessary amount of water was added in the mixture to make a paste. The paste was then passed through a needle connected with a 10-mL plastic syringe. A noodle shape feed (0.1–0.2 cm in diameter) was then collected in a steel tray. The tray was kept under the sun for 6 h to obtain crispy noodle shape (0.1–0.2 cm in diameter) feed. Two fish bioassays were conducted to study (i) the toxicity of the Chroococcidiopsis sp. Biomass; and (ii) the use of the formulated feed. The dried noodle feed (DNF) was then manually made to smaller parts and fed to 10 Zebra fishes (in 10-L glass container) for 7 d, and 5 Koi fishes (in 80-L glass aquarium) for 30 d. As a control, another set of experiments were conducted with commercial fish feed. For the Koi fishes, the feeding rates in both aquariums were 0.5 g feed/d/fish (0–15 d) and 0.75 g feed/d/fish (16–30 d).
Results and Discussion
Self-Settling Study of Chroococcidiopsis sp.
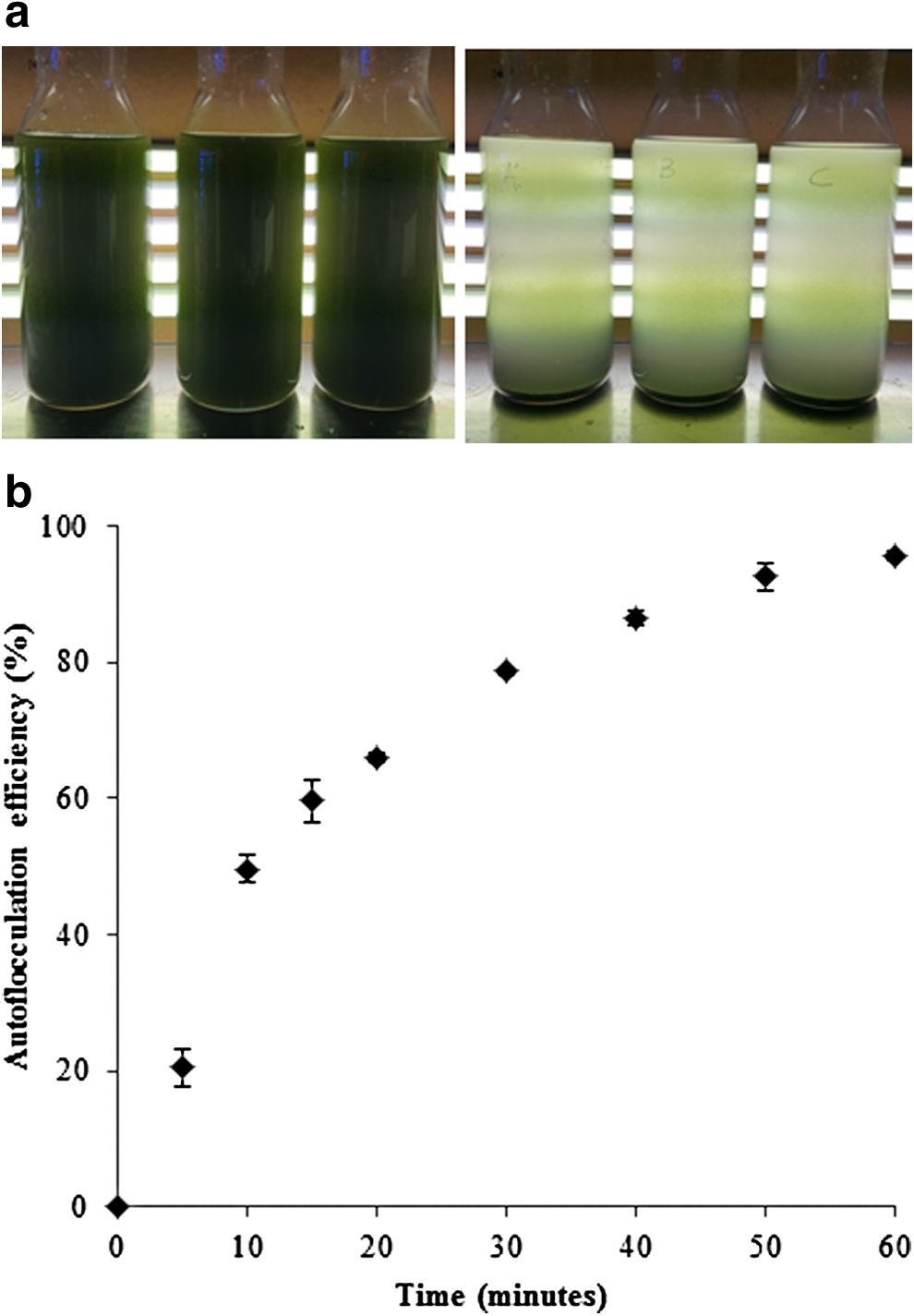
Under low light condition (<100 μmol E/m2/s), the culture is usually greenish black. However, with higher light intensity it turns into reddish brown. In the indoor experiment, the strain had an average specific growth rate of 0.44/d (data not shown). The strain has an average diameter of 2–6 μm; however, the cells are not found solitary. On the other hand, Chroococcidiopsis sp. forms regular and irregular shapes large agglomerations (Fig. 1). At any point of the growth cycle, in the absence of mixing, the biomass spontaneously settles to the bottom of the container; this phenomenon could be exploited in large scale biomass production as it would significantly reduce the energy and cost associated with the biomass production.

100x magnification image of the Chroococcidiopsis sp. culture (before settling). Color images available online at
In the small-scale settling study, 50% of the biomass precipitated within first 10 min of the experiment; after 1 h, 97% of the biomass settled at the bottom (Fig. 2a–b). The culture, used for the settling study, had a pH value of 8.5; however, culture pH value (7.5–9.5) had no effect on the settling efficiency of Chroococcidiopsis sp. (data not shown). The strain showed similar settling efficiency for a culture salinity range of 4–8% NaCl (data not shown). Taking advantages of the s aggregate forming and self-settling properties, the strain was studied to coprecipitate other small potential microalgae of interest (i.e., Nannochloropsis sp., Tetraselmis sp. and Scenedesmus sp.). The strain did not coprecipitate other microalgae after mixing at equal volume, providing utility for separation of this cyanobacterium of interest from potential contaminating microalgae. After 1 h of mixing, only a major fraction of Chroococcidiopsis sp. settled at the bottom, whereas the supernatant contained the other microalgae.
Outdoor Growth Study of Chroococcidiopsis sp.
In the first phase (i.e., initial 10 d) of growth study, areal biomass density of Chroococcidiopsis sp., in both the tanks, are shown in Fig. 3a. The maximum biomass productivities were 21.5 and 23.4 g/m2/d, on day 5, for tank 1 and tank 2, respectively. However, as the density of the culture reached above 0.4g/L, biomass productivities were reduced drastically. Additionally, the growth rate was low in the lag phase as the strain was acclimating to the outside environment. Therefore, the average biomass productivities were 8.16 and 8.8 g/m2/d for tank 1 and tank 2, respectively.

Biomass density of Chroococcidiopsis sp., in both tanks, exceeded 0.45g/L on the 10th day of cultivation. From days 10–70, 50% of the culture was harvested every alternative day, regardless of the biomass density. Daily total irradiated photoactive radiation (PAR) and air temperature are shown in Fig. 3b and 3c, respectively. Average values of daily total PAR and air temperature were 77.2 MJ/m2/d and 21.2°C respectively. There were huge fluctuations in both daily irradiated PAR values (3–10 MJ/m2/d), and the differences in daily maximum and minimum air temperature values (5.7–16.1°C). From the 28th to 31st day of the experiment, there were intermittent rain and occasional cloud cover. Although the growth media was recycled, fresh water was added to compensate for evaporation loss. In the first phase (10 d), average evaporation water loss was 1.08 cm/d. However, the average evaporation water loss in the second phase (60 d) was 0.72 cm/d, which was mainly due to the decrease in air temperature.
Throughout the entire growth period, both the cultures had similar brown color appearance (Fig. 3d), and self-settling efficiency of the strain was 90% or above for the cultures in both tanks, although the actual amount of biomass harvest varied (Fig. 3e). After decanting 95 L of the growth media back to the cultivation tank, 5 L slurry was then further concentrated using cloth filter (50 μm mesh). Filtered growth media (over 4.5 L) was also recycled back to the culture. Final biomass paste had a solid content of 7–10%. The average biomass harvest rates (ash free dry weight basis) were 15.97 g/m2/d and 16.2 g/m2/d for tank 1 and tank 2, respectively. Areal biomass productivities for some other reported microalgae, in outdoor continuous cultivation mode, are listed in Table 1. 25 –35 A large variation in areal biomass productivities are observed among these strains, which could be due to the characteristics of the microalgae (average specific growth rate) and growth conditions (temperature, light intensity, the design of the cultivation pond, etc.). While most other microalgae were harvested using energy-intensive harvesting methods, Chroococcidiopsis sp. was harvested by simple self-settling. Very similar to this strain, some diatoms (e.g., Staurosira sp.) are also known to aggregate and settle. 23 Therefore, these strains will not only reduce the cost of biomass harvesting but also allow recycling of culture media.
Comparison of Outdoor Biomass Productivities, Duration of Cultivation, and Pond Size for Some of the Selected Strains with Chroococcidiopsis sp.
NA, not available.
No other microalgal or cyanobacterial contamination was observed, in both tanks, throughout the growth period. It was possible that larger size flocs of Chroococcidiopsis sp. in the culture entrapped any incoming microalgal contamination and prevented propagation of such contamination. Similarly, it would have been difficult for other grazing microorganisms (amoeba, zooplanktons) to consume large size floc of Chroococcidiopsis sp., and hence, these microorganisms were not observed in the culture. It was shown that the effect of grazers contamination was lesser in the cultures of colonial cyanobacteria as compared to filamentous and other types of cyanobacteria. 24 However, the average ash content in outdoor grown Chroococcidiopsis sp. biomass (28%) was much higher than ash content of indoor grown biomass, (12.3%) which could be due to incoming sand, and dust particles that were carried over by occasional sand storms. The strain was grown, in the same scale, in the summer for 1 month and no contamination was observed. Three more strains (Dunaliella sp., Synechococcus sp. and Tetraselmsis sp.) were grown simultaneously in 3 other identical tanks. Similar to Chroococcidiopsis sp., Tetraselmis sp. culture was not contaminated; however, the other two species got contaminated by Tetraselmis sp.
Variation in Metabolites of Chroococcidiopsis sp.
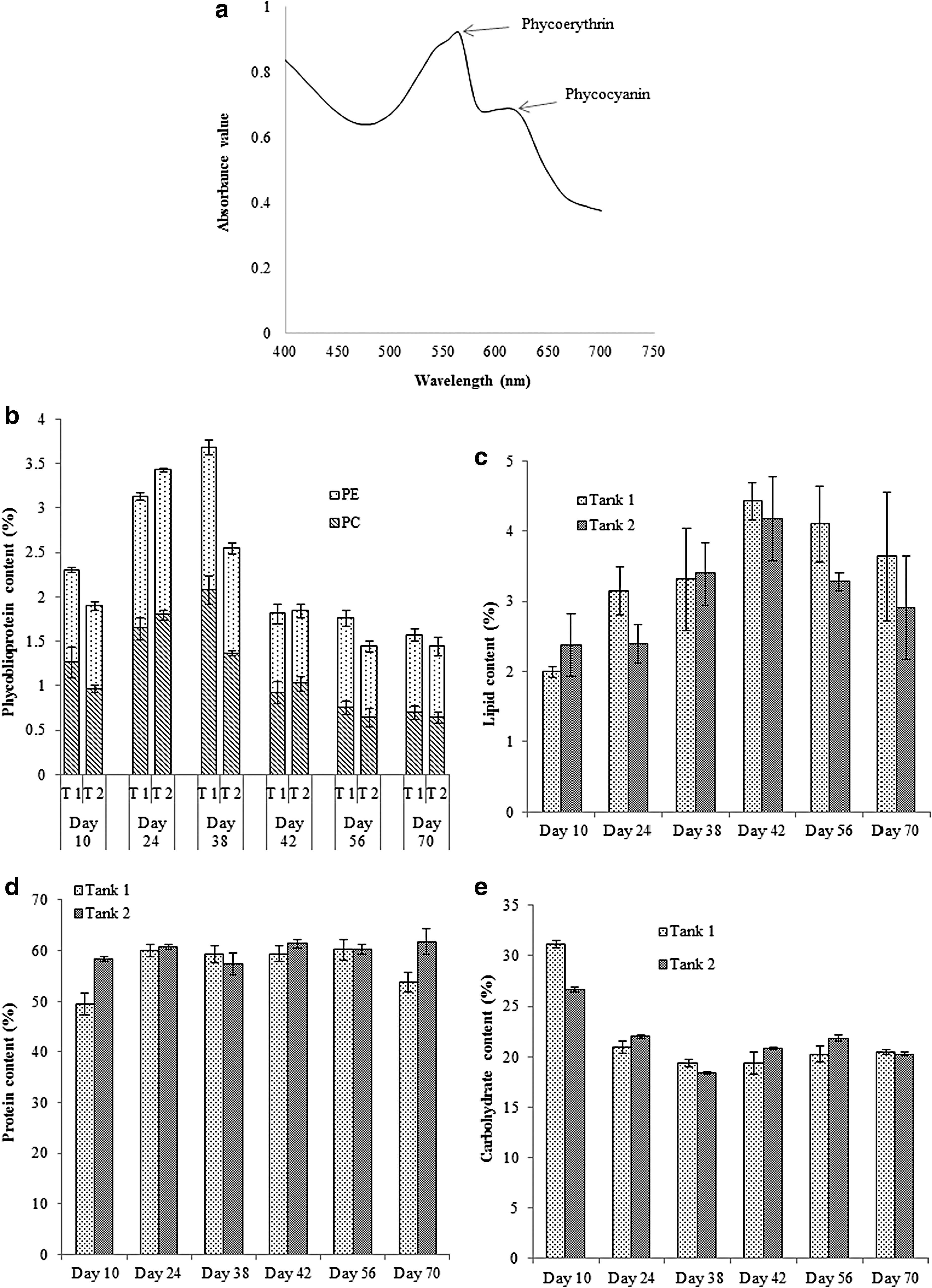
A typical UV spectrum scan of the water extracted pigment sample (d 24, tank 1) is shown in Fig. 4a. Total phycobiliprotein, lipid, protein, and carbohydrate content of Chroococcidiopsis sp., in both tank cultures, on specific day of outdoor growth study, are presented in Figs. 4b and 4e. Average phycobiliproteins contents of the strain were 2.37% and 2.16% for tank 1 and tank 2 respectively, whereas, average phycobiliprotein productivity were 1.90 and 1.73 mg/l/d for tank 1 and tank 2, respectively. Although the strain was grown in continuous mode, it was still able to accumulate high concentration of phycobiliprotein (>1.43%; see Fig. 4c). In a separate indoor batch cultivation (700 μmol E/m2/s, 10-d growth period, Guilllard 3.F/2 nutrients) it was found that the strain accumulated 2.32% of phycobiliprotein (1.18% PC and 1.14% PE). However, the strain had a maximum of 3.68% of total phycobiliprotein in tank 2 culture on day 38. The total concentrations of PC and PE of some other strains are listed in Table 2. Ishida et al. (1997) and Billi and Caiola (1996) found the presence of phycocyanin alone in Chroococcidiopsis sp., 36,37 whereas, the local strain accumulated both phycocyanin and phycoerythrin in indoor and outdoor cultures. Variation in nitrogen, light intensity, and temperature could affect the ratio of intracellular phycobiliproteins. 38 Average phycocyanin content values, in the harvested biomass, were 1.22% and 1.05% for tank 1 and tank 2, respectively. Similarly, average phycoerythrin content values were 1.15% and 1.05% for tank 1 and tank 2, respectively. Phycobiliproteins have been reported to exhibit antioxidant, anti-inflammatory, neuroprotective, hypercholesteromic, antiviral, antitumor, and hepatoprotective properties. Phycobiliproteins are also used as food colorants. 38 It was reported that presence of both PE and PC in the fish feed was beneficial to the fishes. 39 Hence, it might not be required to separate PC and PE. The easily extracted pigments of Chroococcidiopsis sp. can be included in the fish feed. Furthermore, because of high protein content and presence of necessary essential amino acids (EAAs), whole biomass of Chroococcidiopsis sp., can be used as fish feed ingredient.
Concentrations of PC and PE in Other Strains
Lipid content of the cyanobacteria was very low as compared to lipid content of many other green unicellular microalgae. Average lipid contents of Chroococcidiopsis sp. were 3.43% and 3.09% for tank 1 and tank 2, respectively. The strain had a maximum of 4.42% of lipid on 42nd-day culture in tank 1 (Fig. 4d). In the indoor batch culture, the strain accumulated 5.83% lipid content. However, Ishida et al. (1997) found a much higher (approximately 10–15%) lipid content in Chroococcidiopsis sp. 36 While Ishida et al. used gravimetric method to quantify the lipid, in this study, colorimetric sulpho-phospho-vanilin method was used to determine the lipid content. Since the outdoor culture was maintained in continuous mode, it could have resulted lower intracellular lipid content. Although microalgal lipid content is very crucial for making biodiesel, biocrude production using HTL technique does not require lipid rich biomass.
Hydrothermal liquefaction is a promising technology as it converts the whole microalgae biomass to in its wet condition to biocrude oil which can be later fractionated into biofuels. Recent studies have indicated that biocrude conversion yields from low-lipid feedstocks (e.g., macroalgae, microalgae, cyanobacteria, municipal sewage sludge) have exceeded 40%. 40 –42 Nevertheless, the quantity and quality of the biocrude oil may be influenced by the lipid fraction of the biomass.
Similar to many other cyanobacteria, Chroococcidiopsis sp. is rich in protein. In the indoor growth study, the strain had 47.3% protein content. However, in the outdoor study, the strain had 50% or higher amount of protein throughout the entire growth period. The maximum protein content was observed as 61.8% on day 70 in tank 2. Average protein contents of the strain were 57% and 60% for tank 1 and tank 2, respectively. Nitrogen loading rate in the raceway tank was 1.75 g N/m2/d. Average protein productivities in Chroococcidiopsis sp. were 9.10 and 9.72 g/m2/d for tank 1 and tank 2, respectively. Considering the nitrogen content in protein is approximately 15%, the nitrogen utilization efficiencies by the strain were 84.3%, and 90.05% for tank 1 and tank 2 respectively. Therefore, this strain could also be considered as a potential source of feed. Ishida et al. (1997) found that Chroococcidiopsis sp. Strain TS-821 could accumulate 32–56% protein, under various growth conditions. Higher protein content, obtained in the outdoor growth study, could result due to the continuous cultivation of the local strain in nitrogen rich growth media. The amino acid profile of the strain is shown in Table 3. 43,44 The strain synthesizes all the EAAs; apart from methionine, all the EAAs are present in desirable concentrations. Therefore, the biomass of this strain can be an ideal source of protein in the formulation of feed.
Comparison of Amino Acid Profiles of Chroococcidiopsis sp. with Other Strains
Carbohydrate content of the strain, during the outdoor growth study, varied from 18.42% to 31.4%. In the indoor growth study, the intracellular carbohydrate content was 29.7%. Similarly, much higher content of carbohydrate (30–47%) was found in Chroococcidiopsis sp. TS-821 in the study by Ishida et al. 36 Maximum carbohydrate content was found on day 10, in both tanks, when cultures in both tanks were in stationery mode. As the cultures, in these tanks, were maintained in continuous mode from 10th day, the carbohydrate content varied from 19.35–20.94% and 18.42–22.0% for tank 1 and tank 2, respectively.
As the strain can be harvested by autofloccualtion, neither the metabolites quality will be affected by other chemicals (i.e., coagulant), nor there will be loss of intracellular metabolites which would otherwise occur in different harvesting techniques (i.e., electrocoagulation, centrifugation, filtration). Autoflocculation of the biomass will also allow recycling the culture growth media which, in turn, will reduce the water footprint of biomass production. From the perspective of high-value pigment production alone, Chroococcidiopsis sp. is a very promising strain. Pigment-extracted biomass, still, could be processed by HTL technology for biocrude-oil production.
After completing the growth trials in October-December (70 days), a similar continuous growth study was conducted in the summer (mid August-mid Sepetember; max temperature 48°C) for 1 month. The strain could not only survive the high temperature and light intensity but also maintained above 23g/m2/d biomass productivity. Apart from having desired nutritional properties, the strain can be grown all the year round—hence the strain is a potential candiate for large-scale feedstock production. In the near future, the strain would be grown in the existing 500 m2 (100,000 L) raceway pond, in the same facility, to determine the technoeconomic feasibility of producing biomass.
Bioassay Study
As the Chroococcidiopsis sp. culture samples were added in the artemia culture, the artemia continued to increase in size. Furthermore, in the later part of this experiment, cysts of artemia were visible in the culture. Similarly, for the DNF-fed Zebra fish culture, there was no mortality and no change in color. Therefore, Chroococcidiopsis sp. produced neither any intracellular, nor any extracellular compounds that were toxic to artemia and Zebra fishes. During the course of feed trials, there was neither any change in fish color nor any mortality for the DNF-fed koi fishes. The commercial feed floated in the water, while the DNF used settled to the bottom of the aquarium. DNF-fed Koi fishes gained weight at an average rate of 0.25g/d, while koi in the controlled experiment gained weight at an average rate of 0.28g/d. One of the major reasons behind the lower growth rate for the DNF feed could be the ash content in Chroococcidiopsis sp. In the commercial feed, ash content was 6.7%, which was much lower than the Chroococcidiopsis sp. biomass (28%)—ash content in the DNF was 10.96%.
Also, the commercial feed was comprised of Spirulina sp., a freshwater strain. Mass scale production of Spirulina sp. would depend on the availability of an adequate amount of freshwater. As the world is currently facing an acute freshwater crisis, developing marine microalgae or cyanobacteria biomass as feed ingredients could contribute to achieving sustainable food security. In this regard, Chroococcidiopsis sp. could be a promising marine strain as an ingredient of feed.
Conclusion
In this study, it was demonstrated that Chroococcidiopsis sp. could be grown continuously over a long period of time, and the phycobiliprotein-rich biomass could be harvested using its self-settling property. Self-settling microalgae or cyanobacteria can greatly reduce the cost of biomass production. Furthermore, presence of essential amino acids and phycobiliproteins in the biomass make Chroococcidiopsis sp. a potential fish feed ingredient. Any strain that produces high-value metabolites can also assist in enhancing the economic viability of biofuel production. Thus, pigment extracted Chroococcidiopsis sp. could also be explored as a potential feedstock for biofuels.
Footnotes
Acknowledgments
The authors would like to acknowledge the support of Qatar National Research Fund (QNRF, a member of Qatar Foundation) for providing the funding (under grant NPRP8-646-2-272) for this study.
Author Disclosure Statement.
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.