
Abstract
Energy security, climate change, and economic development are the driving forces behind the increasing focus on biofuel production worldwide. Lignocellulose can provide ample raw materials for biofuel production, via its hydrolysis and the fermentation of the released sugars. Currently, lignocellulases are often used to catalyze this step, which hydrolyze the β-1, 4 glycosidic linkages in lignocellulose. These enzymes offer a good starting point for enhancing the overall economics of biofuel production. To reduce cost in biofuel production, there is a need for enzymes with improved activity and broader substrate range. The rapid development of a variety of protein engineering techniques has facilitated the development of novel biocatalysts with improved properties. A range of specific properties can be enhanced in enzymes, including the catalytic activity, thermostability, and specificity, under industrial conditions.
Introduction
Global demand for energy has grown with development of new industries, requiring constant improvement and the search for new sources of energy. The need for renewable energy generated an expectation for use of biomass as energy source. The current challenge is to unlock the glucose that nature has smartly wrapped into lignocellulosic biomass.
Lignocellulose—the most abundant component of renewable biomass produced by photosynthesis—is synthesized at an estimated rate of some 200 billion tons per year. 1 This biopolymer is composed of cellulose, hemicelluloses, and lignin. Cellulose is the most abundant polymer; in nature, it is found mainly as a structural component of plant and algal cell walls. Biomass is a primary source of food, fodder, and fiber, and as a bioenergy source provides about 10.2% (50.3 EJ/y) of the annual global total primary energy supply (TPES) from a wide variety of biomass sources. 2 More than 80% of biomass feedstocks used for energy is derived from wood and shrubs. The remaining bioenergy feedstocks come from the agricultural sector (energy crops, residues, and byproducts). An International Energy Agency report assessing available residues in 2030 concluded that 10% of global residues could yield around 5.2 EJ lignocellulosic ethanol, or roughly 4.1% of the projected transport fuel demand in 2030, and 25% of global residues converted to ethanol, diesel, or syngas could contribute 13–23.3 EJ globally. 3,4 It is well documented that cellulosic ethanol offers greater environmental benefits and sustainability.
An analysis of US Environmental Protection Agency shows that lignocellulosic ethanol generates 91% less greenhouse gases than fossil-based petrol or diesel in transport applications, compared with just 22% for corn-based ethanol. 5 However, the conversion process relies heavily on major technological innovations centered on effective and low-cost enzymes, feedstocks, and efficient process design.
Lignocellulosic biomass (plant biomass), is a great potential resource because it is inexpensive, largely abundant, and environmentally sound to produce. Agricultural resources (like leaves, stems, and stalks from corn fiber; sugarcane bagasse; rice hulls; woody crops; and forest residues) are a great source of lignocellulosic biomass—which is renewable, chiefly unexploited, and inexpensive. Also, there are multiple sources of lignocellulosic waste from industrial and agricultural processes, e.g., citrus peel waste, sawdust, paper pulp, industrial waste, municipal solid waste, and paper mill sludge. Approximately 70% of plant biomass is locked up in 5- and 6-carbon sugars. In naturally occurring cellulose, the degree of crystallinity varies between 40% and 90% and the rest of the cellulose is said to be “amorphous.” This amorphous cellulose is a very tough substrate that often occurs embedded and entangled with hemicelluloses and lignin—forcing cellulose-degrading organisms to develop more or less typical sets of enzymes, including both endoglucanases and cellobiohydrolases.
The efficiency of cellobiohydrolases, in particular, is enhanced by their strongly progressive action, where a substrate-binding tunnel enhances the probability for the enzyme to remain bound after a catalytic cycle. Cellulose is primarily crystalline in nature. Cellulose chains in the crystals are stiffened by inter- and intra-chain hydrogen bonds, and the adjacent sheets that overlie one another are held together by weak Van-der Waals forces. During biosynthesis, 50–100 cellulose molecules associate to form an elementary fibril with a diameter of 4 nm. About 20 such elementary fibrils form a microfibril, which is readily visible with an electron microscope. Another major component present in the lignocellulosic biomass is xylan, the substrate of xylanases and a major structural polysaccharide in plant cells. Xylan accounts for approximately one-third of all renewable organic carbon on earth. 6 Hemicellulose is a complex of polymeric carbohydrates including xylan, xyloglucan (heteropolymer of D-xylose and D-glucose), glucomannan (heteropolymer of D-glucose and D-mannose), galactoglucomannan (heteropolymer of D-galactose, D-glucose, and D-mannose), and arabinogalactan (heteropolymer of D-galactose and arabinose). 7
This, together with cellulose (1,4-β-glucan) and lignin (a complex polyphenolic compound) make up the major polymeric constituents of plant cell walls. 8 Within the cell wall structure, all three constituents interact via covalent and non-covalent linkages, with the xylan being found at the interface between the lignin and cellulose (where it is believed to be important for fiber cohesion and plant cell wall integrity). 9
Degradation of the polymer is performed by two methods: acid hydrolysis and enzyme hydrolysis. Acid hydrolysis is a widely used method but has disadvantages, namely the generation of hazardous acidic wastes and the technical difficulties in recovering sugar from the acid. Enzymatic hydrolysis is developing much faster, with improvements in cost and efficiency, and the enzymatic method can be employed without generation of any toxic waste. Physicochemical pretreatment of biomass is a prerequisite step before enzymatic degradation for industrial applications, at least. There are myriad of different methods of pretreatment and the optimal method depends on the nature of the biomass (softwood or hardwood).
Cellulose and Hemicellulose Degrading Enzymes
Cellulases include endo-1,4-β–glucanase, exo-cellulase and β-glucosidases. Hemicellulases include endo-β-xylanase and β–xylosidase, acetyl xylan esterase, feruloyl esterase, arabinofuranosidase glucuronoyl esterase, glucuronidase, glucanase, mannanase, xyloglucan hydrolase; monooxygenases. These enzymes are compulsory components of the state-of-the-art enzyme cocktails used in industrial lignocellulose degradation.
Mechanism of Lignocellulose Degrading Enzymes
Endocellulase (EC 3.2.1.4) hydrolyzes internal glycosidic linkages present in the structure of cellulose at random positions. Exocellulase (EC 3.2.1.91) depolymerizes and hydrolyzes cellulose chains typically into disaccharide units, down the length of a chain. β-glucosidase (EC 3.2.1.21) hydrolyzes soluble cello-oligosaccharides into glucose. Xylanase includes β-1,4-endoxylanases (xylanases; EC 3.2.1.8), which cleave internal glycosidic bonds within the xylan backbone. Arabinofuranosidase (EC 3.2.1.55) hydrolyzes arabinose side-chains. α-glucuronidase removes glucuronic acid side-chains from the xylosyl units. Xylan esterase (EC 3.1.1.6) releases acetate groups, and β -xylosidase (EC 3.2.1.37) hydrolyzes xylobiose to xylose.
Basic Research on Cellulase and Xylanase
Although there has been a plethora of research on xylanases and cellulases since the end of World War II, some major gaps still exist in the understanding of the mechanism by which they catalyze the crystalline cellulose. One notable gap is information on the mechanism by which a cellulase binds a segment of a cellulose chain from a microfibril into its active site. This is possibly the rate-limiting step for crystalline cellulose degradation. Therefore, understanding the mechanism of this step is very significant. Another knowledge gap is how cellulosomes are able to efficiently catalyze the cellulose despite their large size restricting their ability to access much of the cellulose surface area that is available to smaller free cellulases. A third gap is an understanding of the way in which certain free, cellulose-binding modules (CBM) stimulate cellulase hydrolysis. It is possible that these domains modify the cellulose, but exactly how this modification done is not known. There are some plausible mechanisms for cellulase synergism. Cellulases, hemicellulases, and pectinases have proved their biotechnological potential in various industries, including animal feed, food, textile and laundry, brewery and wine, pulp and paper, and agriculture, as well as in research and development.
Research work on cellulase and xylanase has been focused mainly on fungi; however, of late, higher-growth rate, alkali-tolerant and thermostable properties have directed attention toward bacterial enzymes. For complete degradation of lignocellulose, multi-enzymes need to be used in large quantities. For example, cellulase enzymes access the glucose polymer cellulose while xylanase acts on xylose polymer xylan; both the enzymes share a common mechanism of action. This is why lignocellulases are found to be bifunctional in nature. The conserved amino acid residues among the glycoside hydrolase family also indicate the structural and functional similarity of the cellulose and xylanase enzymes
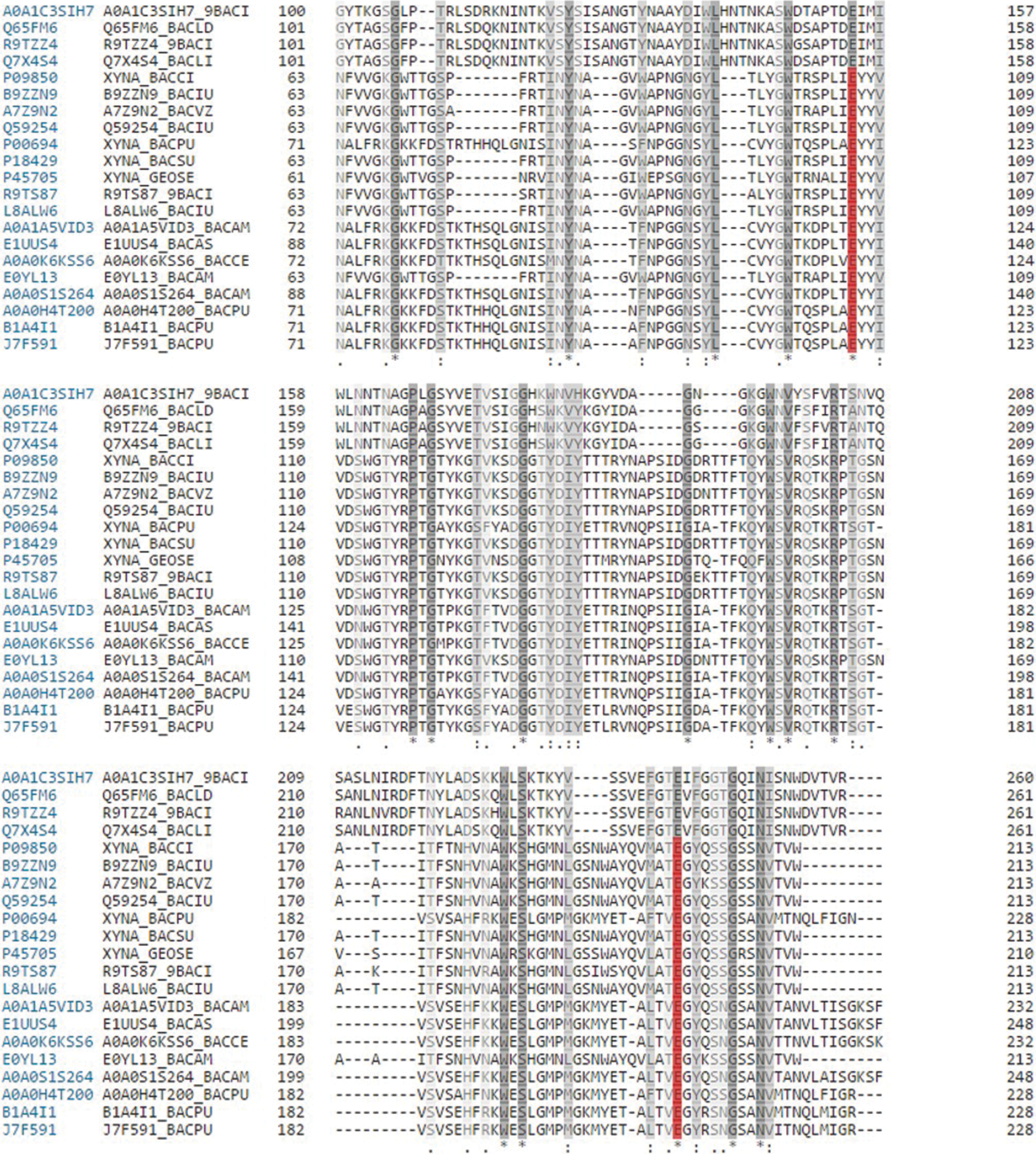
Sequence alignment among cellulases and xylanases. Red color shows active site residues while gray color displays the similarity and conserved residues among glycoside hydrolase family. Color images available online at
A number of studies have been carried out on bacterial and fungal cellulases and xylanases, including enhanced cellulase production by a mutant of Trichoderma reesei; 10 improvement of fungal cellulase production by mutation and optimization of solid state fermentation; 11 fungal strain improvement for cellulase production using repeated and sequential mutagenesis; 11 exploration of cellulose surface-binding properties of acidothermus cellulolyticus cel5a by site-specific mutagenesis; 12 engineering better biomass-degrading ability into a GH11 xylanase using a directed evolution strategy; 13 effect of single active-site cleft mutation on product specificity in a thermostable bacterial cellulase; 14 crystal structure of celm2, a bifunctional glucanase–xylanase protein from a metagenome library; 15 and many more. But still, many modifications to bacterial cellulases and xylanases through protein engineering are required to make more efficient and suitable enzymes for complete hydrolysis of lignocellulose. Natural enzymes typically are quite specific for their specific substrate, and it is often needed to modify or broaden their substrate selectivity for industrial or biotechnology applications. 16
Improvement of Cellulase and Xylanase
In spite of a broad spectrum of these enzymes being isolated, not one is completely suitable as is for the hydrolysis of lignocellulose. Nonetheless, these enzymes offer a good starting point for enhancing the overall economics of biofuel production. Typically, the use of protein engineering technology has been directed toward the study of the catalytic function of cellulase and xylanase. Mutagenesis has provided a means for studying the role of different amino acids within the catalytic domain. Recently, modifications to bacterial enzymes through the use of protein engineering are playing a significant role in the production of efficient hydrolytic enzymes used in a broad range of industries. This includes targeting structural amino acids, other than those amino acids of the catalytic site. There are two major strategies for the improvement of enzymes: rational design and directed evolution.
Rational Design
Rational design is the earliest approach to enzyme engineering. It was introduced after the development of recombinant DNA technology and site-directed mutagenesis more than 20 years ago and is still widely used. This strategy requires detailed information of the protein structure, of the structural causes of biological catalysis, or structure-based molecular modeling and ideally structure-function relationship. Rational design involves (1) selection of a suitable enzyme, (2) identification of the amino acid sites to be altered, and (3) characterization of the mutants (Fig 2). The availability of data on the protein structure of an enzyme or of homologous proteins typically governs the choice of a suitable enzyme for modification. The modification of amino acid sequence can be achieved through site-directed mutagenesis, exchange of elements of secondary structure, or even exchange of whole domains and/or generation of fusion proteins. The faith in the power of rational design relies on the belief that our current scientific knowledge is sufficient to predict function from structure. But such information of structures and mechanisms is not available for the majority of enzymes. However, even if the structure and catalysis mechanism of target enzyme are well characterized, the molecular mutation basis for the desired function may not be achieved. 17
Schematic representation of rational design. Color images available online at
Rational design appears to be a logical method for researchers to examine possible amino acid sites near to the active site or the substrate binding pocket in a 3-dimensional structure. 18 Indeed, many residues distributed over large parts of the protein often confer important properties. Even when large functional changes can be obtained with a few amino acid substitutions, it will often be difficult or impossible to discern the specific mutations responsible. For example, a significant increase (106-fold) in the specificity constant (kcat/Km ) of aspartate aminotransferase favoring valine requires 17 amino-acid changes, only one of which occurs within the active site. 19 A successful computational design to convert non-active ribose binding protein to triose phosphate isomerase was based on 18–22 mutations and exhibited a 105–106-fold activity enhancement. 20
Baker and coworkers reported a 20% improvement in the activity on microcrystalline cellulose of a modified endoglucanase Cel5A from Acidothermos cellulolyticus. 21 The Cel5A endoglucanase, whose high-resolution crystallographic structure has been available, 22 was subjected to a series of mutations designed to alter the chemistry of the product-leaving side of the active site cleft. Using structural information and an assumption that end product inhibition could be relieved by a substitution of non-aromatic residue at site 245, a mutant (Y245G) was shown to increase Ki of cellobiose by 15-fold. 23 The maximum product yield of an endoglucanase from Cellulomonas biazotea mutant 51 was 1.5- to 2.5-fold more than was produced by the wild- type cells and was twice that reported by previous researchers on carboxymethylcellulose (CMC). Similarly, the highest productivity of β -glucosidase by a mutant of C. Biazotea was 2.5-fold more than that of the parent organism, and the mutation stabilized the enzyme thermophilically.
Directed evolution
Contrary to rational design, irrational design is an approach to non-informational protein engineering that utilizes the power of natural selection to evolve proteins and select for those with desired traits. Specifically, it requires the use of DNA techniques such as error-prone PCR (epPCR) and DNA shuffling to randomly generate a large library of gene variants (Fig. 3). It has a great advantage over rational design because it is independent of enzyme structure and of the interactions between enzyme and substrate. However, a major challenge of this method is developing a means to accurately evaluate the performance of mutants generated by recombinant DNA techniques and select the high-performance mutants. The success of this method relies on a large library of gene variants—the larger the library, the greater the chance of mutants with desired properties. This method was used to improve the thermal stability of Clostridium cellulovorans cellulosomal endoglucanase (EngB) in vitro by DNA recombination with non-cellulosomal endoglucanase EngD. 24 Interestingly, a bacterial surface display method was used to selectively screen for variants with improved activity on CMC agar with Congo red staining. Furthermore, directed evolution using epPCR and family shuffling was used to successfully increase the thermal stability of β-D-glucosidases from Paenibacillus polymyxa; desired mutants were screened using a chromogenic substrate. 25 A problem with directed evolution, however, is that it can only be used to screen potential single or with a massive screen potential double mutation, since the mutant library size required to include most possible larger multiple mutations is too large. Even in case of directed evolution, the chances of reversion of mutants is high (likewise done by various physical mutagens).

Schematic representation of directed evolution. Color images available online at
For rational protein design, those amino acid residues that appear “logical” to the researcher examining the three-dimensional structure are usually modified to (i.e., they are close to active site, the binding pocket etc.). Continued progress in technology is making it easier to use rational design to improve cellulolytic enzymes. Hence, site-directed mutagenesis (SDM) is widely used for enzyme improvement. Table 1 lists a few examples of cellulases and xylanases that are mutated for improvisation in their properties by various strategies and methods of protein engineering. 26 –40
List of the Cellulases and Xylanases Enzymes Engineered by Protein Engineering
Future Prospects
Since the early 1950's, scientists have tried to shed light on how microorganisms manage the difficult task of degrading lignocellulose. Since there is a huge amount of lignocellulosic waste present on earth, it has the potential to be a good source of biofuel through hydrolysis and fermentation. It is also important to emphasize that in order for lignocellulose biotechnology to make meaningful impact on developing countries, suitable bioconversion processes need to be developed on a much wider scale. These countries should begin to pull their meager resources and biological science expertise in a cooperative and integrated manner toward modern. Work should focus on advanced genomics and proteomics technologies for identifying novel lignocellulolytic enzymes and engineering enzymes with improved activities suitable for industrial-scale application. Glycoside hydrolase family 7 cellobiohydrolase and Trichoderma reesei Cel7A are “work horses” and the major components of commercial cellulolytic cocktails. Engineering of such enzymes has been employed to alter the processivity, cellulose chain acquisition, product inhibition, CBM, length of the linker, and pattern of glycosylation—to the benefit of biofuel production.
The large market potential and the important role that cellulases and xylanases play in the emerging bioenergy and biobased products industries provide motivation to develop better enzyme preparations for plant cell wall hydrolysis. These improved cellulases and xylanases must also have characteristics necessary for biorefineries, such as higher catalytic efficiency on insoluble cellulosic substrates, increased stability at elevated temperature and at a certain pH, higher tolerance to end-product inhibition, and multi-functional nature higher efficiency for the catalysis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.