
Abstract
This study evaluates the effect of lignocellulosic substrate obtained after hydrogen dark fermentation of food waste on plant growth using the radish Raphanus sativus L as an example. Addition of substrate at concentrations of 66 and 132 g/m2 into plain sand caused a strong inhibitory effect on seed germination and root and shoot growth, while a concentration of 33 g/m2 showed significant positive effect with 40% higher seed germination and 90% higher survival after drought stress compared to plain sand. Application of lignocellulosic substrate increased the amount of ammonifying and free-living heterotrophic diazotrophic bacteria that may act as plant probiotics supplying mineral nitrogen for plant nutrition in arid conditions. Preliminary results suggest that lignocellulosic substrate may be used as biofertilizer.
Introduction
Increasing supply of food and energy crops is an important challenge facing modern agriculture. Efficient solutions are required to maintain soil fertility, avoid soil depletion due to intensive use, and develop new acreages from low fertility, dry, and marginal lands. 1,2 Recent approaches in this area are focused on application of eco-friendly biological methods, 3,4 with significant contribution from microorganism-based solutions. These include enriching soil and sand with biomass of microbial origin, 5 microbial-derived biofertilizers, 5,6 plant probiotics, 6 –8 and the use of microbial metabolites that act as plant biostimulants and protect crops from pathogens. 9
Using excess microbial biomass, side products, and undigested residues from microbial methods of organic waste recycling as biofertilizers is promising for several reasons. The method uses cheap and abundant organic substances as raw material, enables the development of zero-waste environmentally safe recycling processes, and allows for the manufacture of valuable products from waste. 10 At the moment, biofertilizers based on treated sewage sludge 6,9 and digestate from biogas production from agricultural and food waste, 11 –13 compost, 14,15 and manure 16 have proven effective in improving the quality of soils and increasing crop yield. Such complex biofertilizers supply soil with a mixture of micro- and macronutrients and also enrich soil with microbial communities. 17,18
Along with recognized microbial methods, some authors mention biohydrogen dark fermentation as a possible alternative to agricultural and food waste recycling. 19,20 Despite a number of challenges related to development of a sustainable fermentation process and increase in biohydrogen yield, scaling-up of hydrogen fermentation of food waste is considered promising because of clean energy production, reduced use of chemicals and carbon emissions, lower energy consumption, 21 and the ability to produce a number of valuable products. 22 –24 Our previous experiments showed that hydrogen-producing microbial communities can be extremely efficient for decomposition of multicomponent food waste. Fermentation by heat-pretreated soil microbial communities in mesophilic conditions results in a 92–98% decrease of weight and volume of solid food waste. 25 In different fermentation cycles, 2–8% of waste is left undigested, depending on waste composition. Dark fermentation provided by hydrogen-producing bacteria significantly differs from methane fermentation by composition of microbial communities and final products. Firstly, heat pretreatment enriches microbial communities with spore-forming species 26 and eliminates methanogenic, sulfate-reducing bacteria and pathogenic species, 27 which are undesirable for introduction in soil. Secondly, hydrogen-producing bacteria do not provide fermentation of lignocellulose under mesophilic conditions. 28 Since lignocellulose is an important structure-forming soil component, undigested residues might be used for bioremediation of deteriorated soils. 29 These considerations support the hypothesis that undigested residues can be used as biofertilizer and a source of microorganisms for depleted soils. To the best of our knowledge, there were no previous studies on the possibility of applying undigested residues following fermentation with hydrogen-producing microorganisms. Therefore, the objective of this study was to assess the effect of lignocellulosic substrate (LCS) and microbial communities resulting from dark hydrogen fermentation of food waste on plant growth in a mixture with sand as a model of low fertility and exhausted soil.
Materials and Methods
Microbial Inoculum
Seed inoculum for dark fermentation process was prepared by heat-pretreatment of soil to select spore-forming aerobic destructors of waste polymers and anaerobic hydrogen-producing bacteria. 100 g of soil was mixed with an equal amount of water and boiled for 30 min at 100°C.
Biohydrogen Dark Fermentation
The fermentation was conducted in a self-designed batch reactor with a transparent chamber and built-in mechanical mixer, as described by Tashyreva et al. 25 Typical kitchen food waste, consisting of raw vegetable and fruit peels (50%), bread and cereals (30%), and raw and cooked meat residues (20%) were grinded to the size of solid particles (1.0–1.5 cm3) and pasteurized for 10 min before fermentation. For the digestion cycle, 2 kg of waste, 6 L of tap water, 100 mL of soil bacteria inoculum, pH indicator bromthymol blue, and 100g of CaCO3 powder were loaded into the reactor chamber, leaving air in a headspace as a gas phase. The fermentation was carried out at 30°C under previously established optimal mixing mode (10 min of mixing, 24 rpm/20 min of pause), and maintenance of pH value in optimal range 6.5–7.0. Acidification was rapidly assessed by changing color of the pH indicator from dark green to yellow and then measured by porous glass electrode ESК-10603/4 using pH-meter-millivoltmeter, pH-150 MA (Gomel Plant of Measuring Instruments, Republic of Belarus). The CaCO3 powder was used to increase buffering capacity of fermentation liquid, and concentrated NaHCO3 solution was added to adjust sharp pH fluctuations during the most intensive phase of waste decomposition. Total gas volume and chromatographic analysis of gas samples were measured twice a day. Biogas composition was analyzed using the chromatograph LHM-8-MD (Moscow Experimental Plant Chromatograph, Russia) equipped with thermal conductivity detector and two stainless-steel columns packed 13X (NaX) Porapak-Q carrier with argon as carrier gas: first (I–3 m/3 mm) for H2, O2, N2 and CH4 analysis and second (II–2 m/3 mm) for CO2 analysis; column and detector temperature 60°C; evaporator temperature 75°C; detector current 50 mA; and gas flow rate 30 cm3/min. Duration of the fermentation cycle was determined by termination of gas production and visual observations of waste degradation. Residues were obtained for further experiments from more than 50 batch fermentations to establish optimal fermentation mode during 2015–2017.
Processing of Lignocellulosic Residues
After the end of the digestion cycle, the undigested substrate was collected from the reactor and transferred into a flask. It was then mixed with water (the weight ratio of the undigested substrate and water was 1:10) and incubated in aerobic conditions at 30°C (shaking for 6 h, 60 rpm). The substrate was then rinsed in tap water, weighed, and dried at room temperature. This was done to promote aerobic decomposition of fatty acids that resulted from hydrogen fermentation of food waste since these compounds may have an inhibitory effect on microbial and plant growth.
Assesment Effect of Lignocellulosic Residues on Plant Growth
Plain sand and commercial soil mixture produced by local manufacturer Agrosvit (Rivne, Ukraine) were used as controls. Sand is considered a poor substrate due to its low nutrient content, microbial diversity, and water-retaining capacity. Using it as an agricultural substrate results in weak plant growth and low resistance to drought stress. A commercial soil mixture consisting of peat and sand was used as a model fertile substrate, enriched with humus and microelements. This also ensured optimal balance between drainage and water retention. Lignocellulosic substrate (LCS) enriched with microbial communities added to plain sand was used to explore whether these undigested residues may improve sand quality as an agricultural substrate. Radish Raphanus sativus L. seeds of Mila cultivar (Nasinnya Ukrainy) were sown in plastic pots (0.14 × 0.14 × 0.68 m, 20 seeds/pot). For the experimental setup, LCS was mixed with sand at a depth 0.1 m in proportions of 132 g LCS/m2 of sand (sand/LCS-1), 66 g LCS/m2 (sand/LCS-2), and 33 g LCS/m2 (sand/LCS-3). These proportions (with diminishing LCS content) were used based on the results of our previous experiment which demonstrated the toxic effect of LCS addition to sand in high concentration. 30
R. sativus L. were cultivated indoors in the photobox for 9 days (at 20°C, air humidity 60–65%, photoperiod 10 h). Light intensity was not measured. The plants were carried out once a day (1 L/pot) during seven days of cultivation. After that, the plants were subjected to drought stress (2 days without watering). The effect of LCS additive on plant growth was assessed by the measuring seed germination, length of main root, length of shoots, and weight of plants after drought stress. The experiment was carried out in triplicate. Statistical analysis (ANOVA, Tukey's test) was performed using built-in functions of R software environment. 31
Microbiological Analysis
The initial estimation of microbial communities' activity was conducted by counts of ammonifying and free-living diazotrophic bacteria in lignocellulosic undigested residues and in a mix with sand after 9 days of plant growth. Bacterial suspension was washed with isotonic solution from 1 g of dry milled LCS, LCS/sand mixtures, and controls, seeded from the tenfold dilutions in triplicates and incubated during 7 days at 28°C on Ashby Agar (HiMedia®) for detection and counts of diazotrophic aerobic bacteria 32,33 and peptone-based Nutrient Agar (HiMedia®) with addition of sterile phenol red (HiMedia®) solution as pH indicator for ammonifying bacteria. A change in agar color from yellow (pH = 6.8) to red (pH ≥8.2) around growing colonies indicated cleavage of NH2 group from amino acids and formation of NH4 + ion. 34 Morphology and ability to produce spores of isolated strains was studied by Gram staining and endospore-staining with Malachite green. 35
Results and Discussion
Results of Biohydrogen Dark Fermentation
Three batch fermentations of multicomponent kitchen waste showed similar results: fermentation cycle duration of 4 days; average biogas hydrogen content of 40%; and biohydrogen yield in range 51–58 L/2 kg of raw waste. Since fermentation process is not a focus of this study, additional details are not provided. Advanced data about dynamics of the fermentation process, digestion, and biogas yield parameters can be found in the previous paper. 25 The fermentation process was extremely efficient and resulted in digestion of 99% of solid waste particles into liquid products, the weight of undigested residues after drying made 4.8–6.2 g from two kg of raw waste. Kitchen waste normally contains a small amount of lignocellulose because inedible parts are removed at early processing stages before reaching a customer. 36 The content of cellulose fibers in grain, fruit, and vegetable waste at the manufacturing stage is higher and may comprise up to 50%. 37 The residues were easily separated from fermentation liquid and consisted of solid dark and light brown fibers and thin peels, which are assumed to be lignin-cellulose (Fig. 1).

Lignocellulosic residues obtained after hydrogen dark fermentation.
Effect of Lignocellulosic Substrate on Plant Growth
The impact of lignocellulosic addition on sand properties was assessed by two main criteria: 1) stimulation of plant growth (by evaluating the percentage of germinated seeds, length of main root, and length of shoots; and 2) capacity of plants to survive under short-term drought stress conditions (by evaluating plant weight after the two-day drought stress).
Lignocellulosic residues saturated with hydrogen-producing bacteria showed different effects on the radish, depending on the concentration in a sand mixture. The highest seed germination (100%) was detected after 3 days in mixture containing 33 g LCS
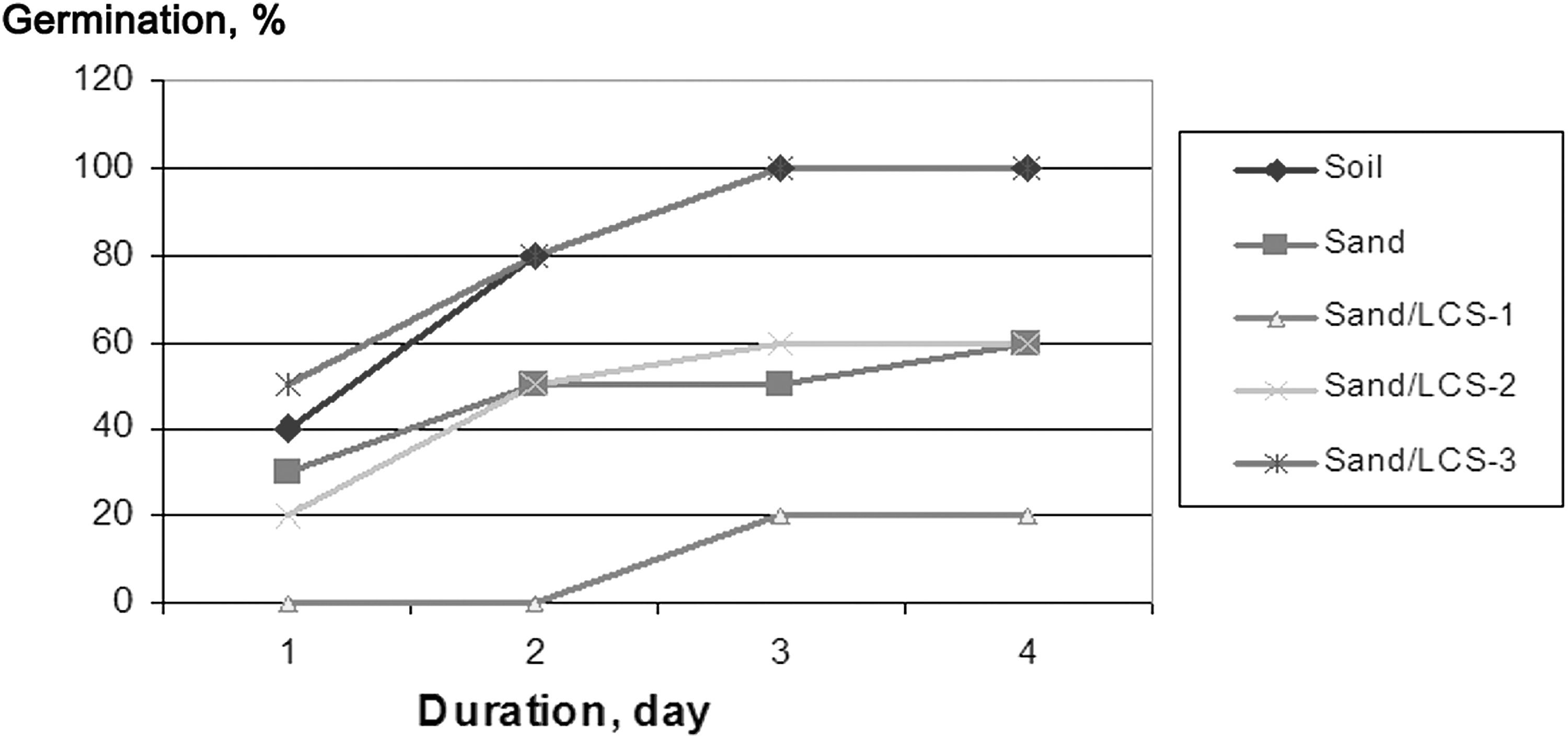
Percentage of germinated Raphanus sativus seeds grown on different substrates.
Growth of radish seedlings greatly differed in control substrates and experimental mixtures (Fig. 3 –5). LCS-1 and LCS-2 (Fig. 5b) showed strong negative effect, completely suppressing root growth, and shoot length was the shortest among all variants, while LCS-3 did not show inhibition effect (Fig. 5d). It is evidence of the toxic effect of using of high concentrations of LCS. Difference in the root length (P ≤ 0.01) was detected for sand (128.5 ± 14.2 mm) and soil (74.0 ± 29.8 mm) controls. Mean value of root length in the sand mixture LCS-3 was 31.9 mm lower than in the plain sand (96.6 ± 14.0 mm). However, this difference was not statistically proved (Fig. 3).
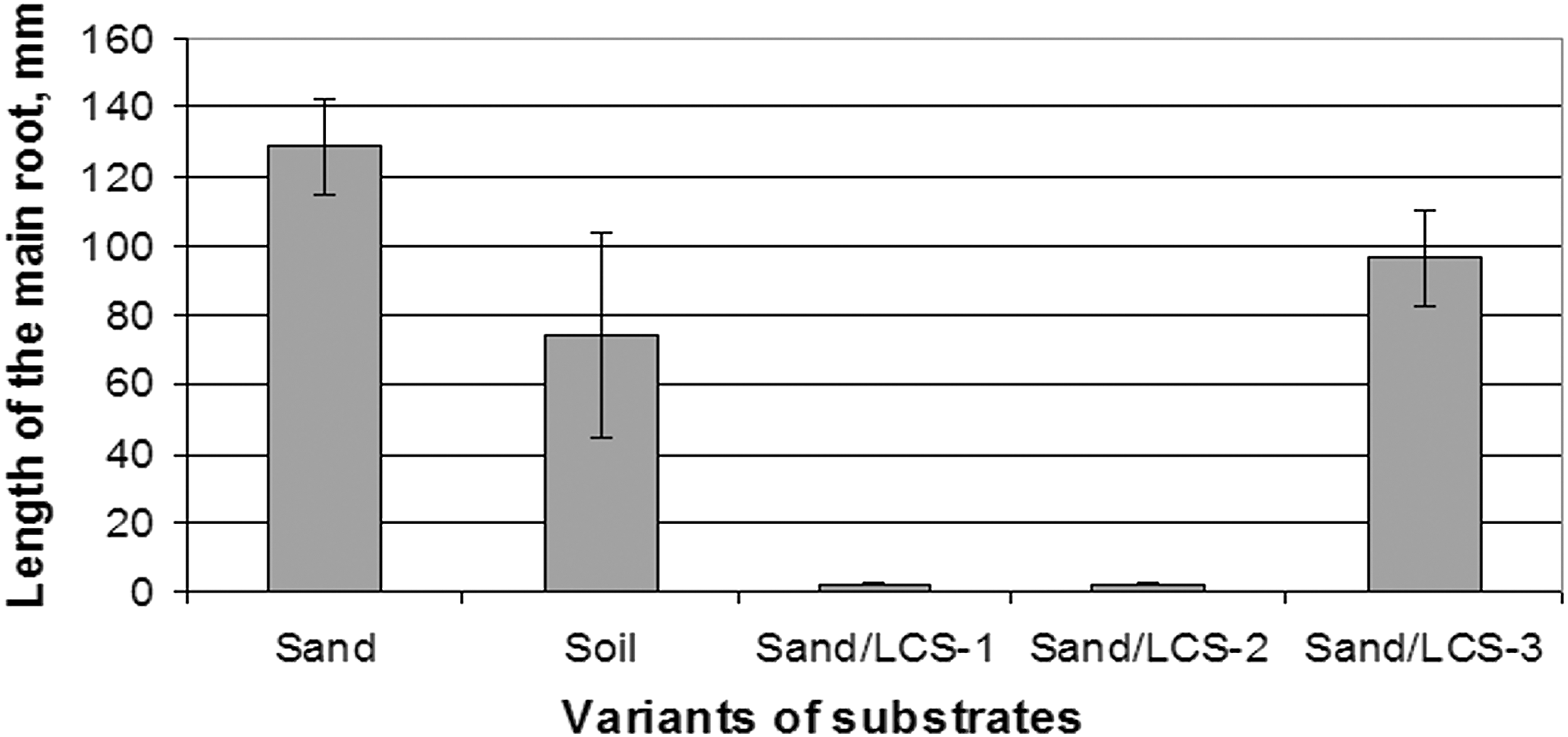
The length of the main root of plants grown on different substrates.
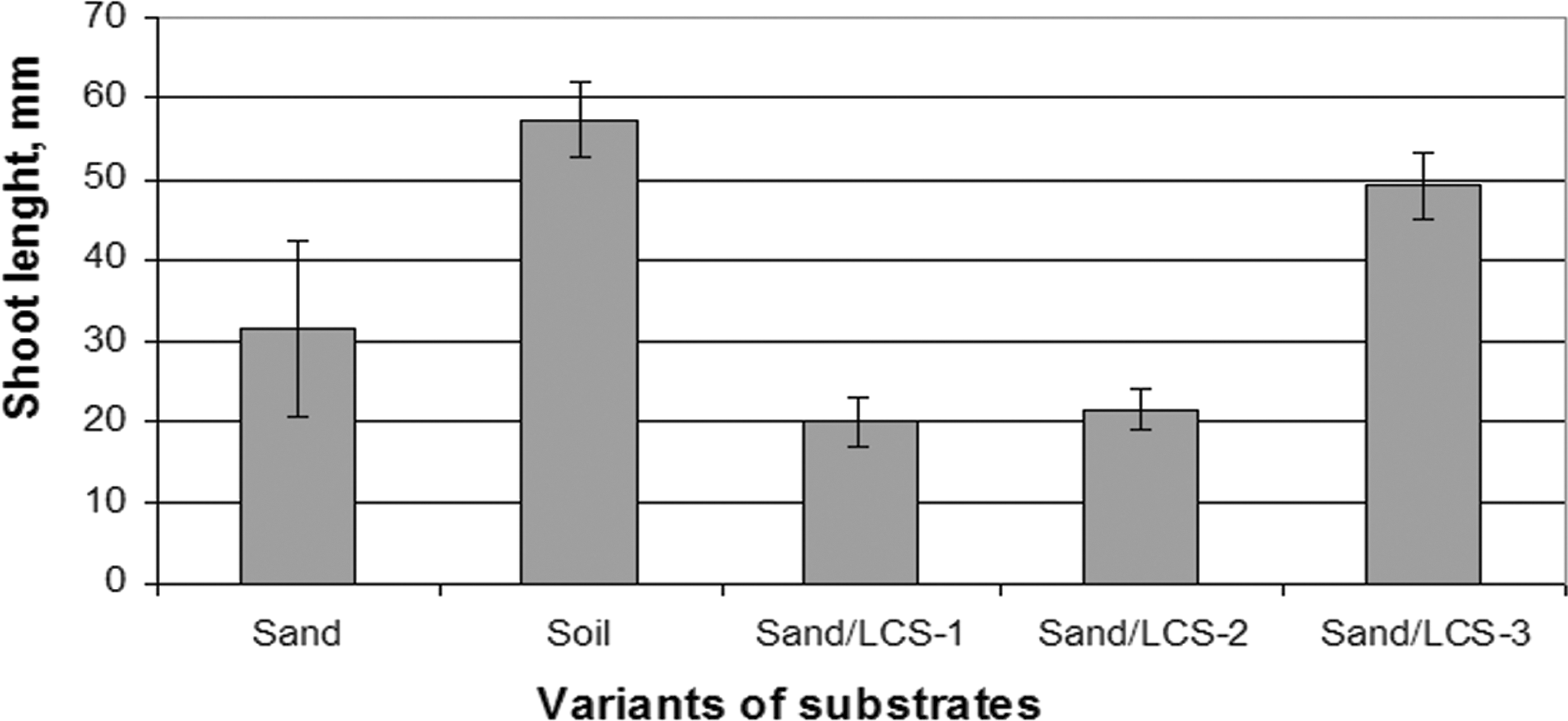
Shoot length of plants grown on different substrates.
Plants after 9 days growth in
Shoot length of the plants grown in the sand mixtures with high LCS content was 2.9 and 2.7 times less, respectively, than in control soil. Statistical analysis did not reveal differences in shoot length between the plants growing in soil, sand, and LCS-3 (Fig. 4). Short duration of the experiment might be a possible reason for this effect. At the same time, it must be noted that LCS of lowest concentration (LCS-3) did not oppress shoot growth. Thus, decrease of LCS content in soil promoted plant survival on LCS-3 substrate, and this preparation was not toxic for the plants.
Addition of lignocellulosic substrate into sand has led to a significant increase in the survival of plants in stress conditions induced by short-term lack of water. Absence of irrigation for two days caused fading of shoots, and only 10% of seedlings survived in the plain sand ( Figs. 5,6 ).

Effect of drought stress on the survival of plants in
At the same time, 100% of plants survived in LCS-3 sand mixture, as well as in soil control. (Fig. 5a-c).
Significant difference (P ≤ 0.05) was found in biomass of plants grown on plain sand and LCS-3 mixture, whereas the weight of seedlings was similar in soil and LCS-3 substrate. (Fig. 7).
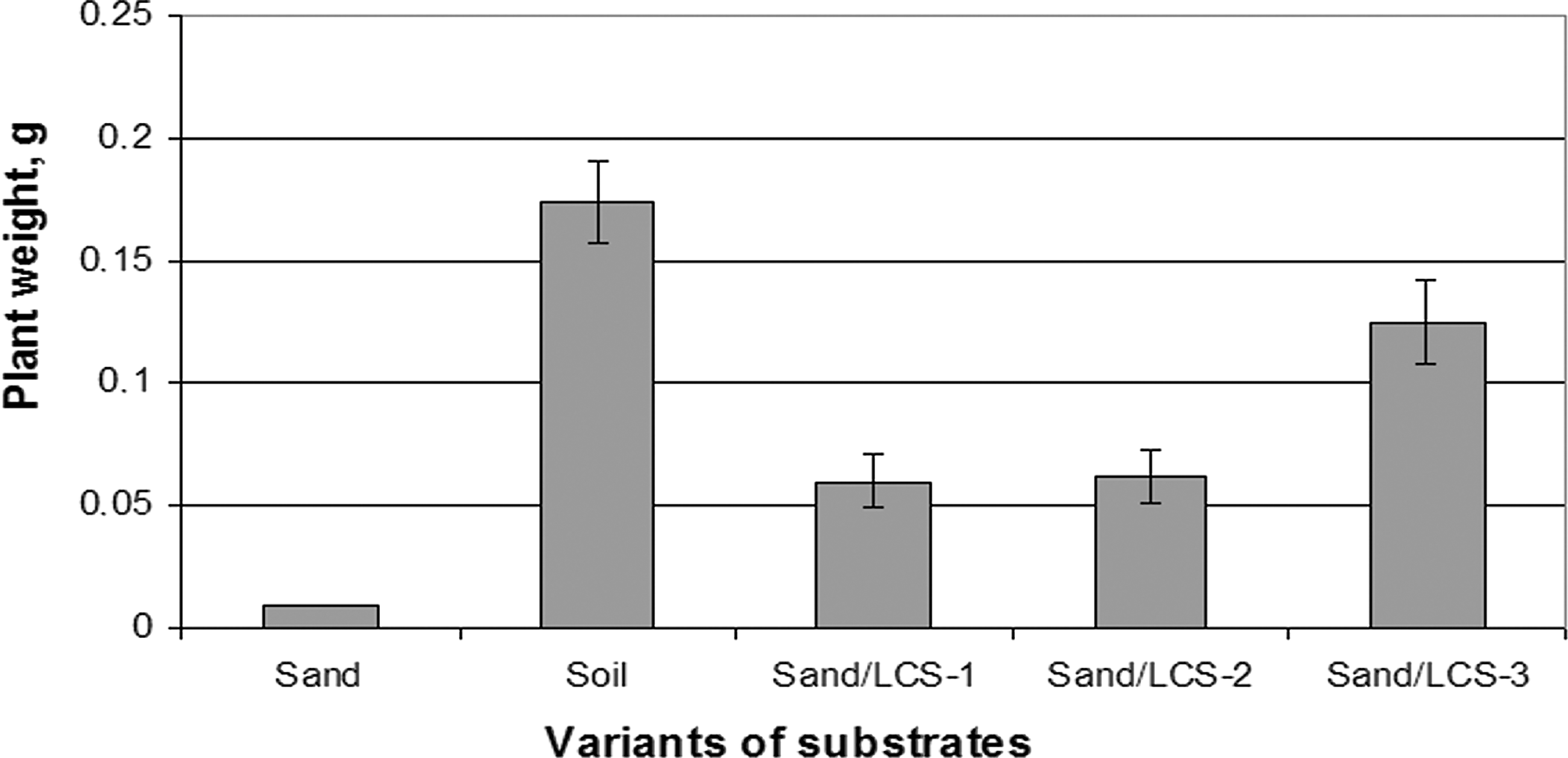
Weight of plants after two days without watering.
Other studies also reported that lignocellulosic side products of the pulp and paper industry improve soil structure and increase water retention. 38 Addition of lignin and cellulose in arid soil launched biogeochemical transformation of carbon, 39 and crystalline cellulose enhanced growth and activity of microbial communities in semi-desert soils. 40
Microbiological Analysis
Undigested lignocellulosic residues showed high content of bacteria of nitrogen cycle. Pure lignocellulosic substrate counted 7.2 × 10 7 CFU/g of ammonifying aerobic bacteria and 5.0 × 10 6 CFU/g of free-living aerobic/microaerophilic diazotrophs. After 9 days of experiment, the number of microorganisms from both groups in all sand/LCS mixtures was in the range of 107–108 CFU/g (Figs. 8,9). This indicates intensive growth of microbial population from dried residues, especially taking into account that LCS was added into the sand in a small proportion. Plain sand counted 1.0 × 103 CFU/g of ammonifying bacteria and 1.0 × 102 CFU/g of free-living diazotrophs; this was, on average, 4 and 5 orders of magnitude less compared to all sand/LCS mixtures. In all experimental mixtures, the number of studied bacterial groups was similar to the soil control. Also, we observed a positive relationship between the number of microorganisms and LCS content in the sand mixtures; the number of both groups from mixed substrate with 132 g/m2 LCS content was two times higher than from mixed substrate supplemented with 33 g/m2 LCS. Though high LCS content caused inhibition of plant growth, we suspect it is not connected to the number of studied microorganisms. The main reason plant growth was inhibited could be due to a residual amount of fatty acids on the surface of lignocellulosic particles. The distinctive feature of hydrogen fermentation is formation of significant amount of acids that may suppress plant growth. Previous study showed that hydrogen fermentation under similar conditions resulted in production of butyric and hexanoic acids in the concentration range 600–2,100 mg/L. 41 Literature data also confirm that fatty acids inhibit the growth of microorganisms and plants. 42 –45

Content (CFU) of ammonifying bacteria in substrates.

Content (CFU) of diazotrophic bacteria in substrates.
Aerobic bacteria of lignocellulose substrate were gram-positive spore-forming rods, and most probably were selected during heat pretreatment of soil microbial communities (Fig. 10). Although dark fermentation of food waste is an anaerobic process, the number of aerobic bacteria in undigested residues was extremely high. We suppose that enrichment of microbial communities with aerobic/microaerophillic species occurred at the initial stage of fermentation because we did not apply replacement of air to inert gas in reactor headspace, and also at the last stage during aerobic treatment of liquid metabolites.
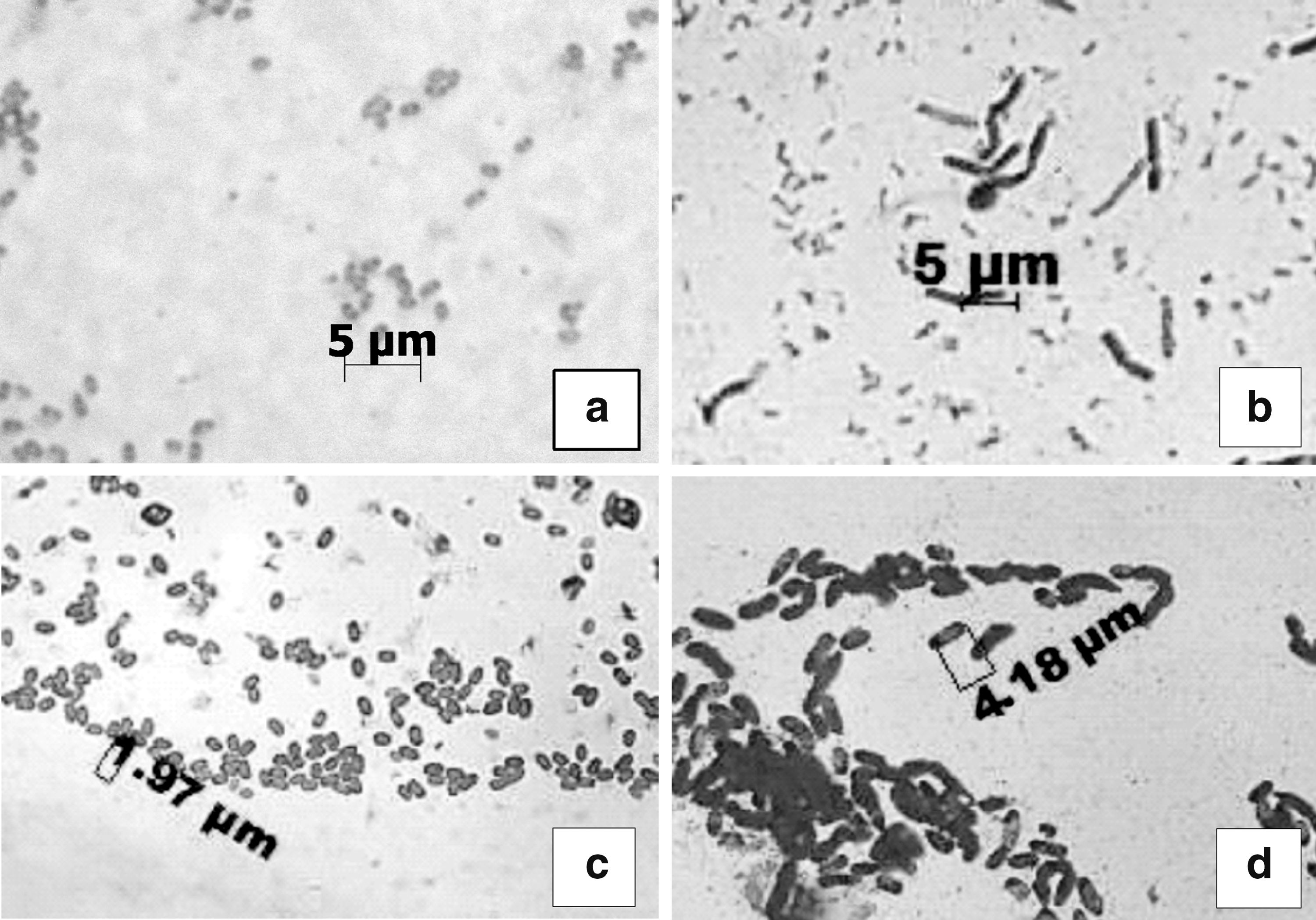
Examples of aerobic spore-forming rods isolated from lignocellulosic substrate;
Clostridia are considered the main heterotrophic hydrogen producers. 46 16S rRNA pyrosequencing of heat-pretreated anaerobic sludge of microbial communities in an active phase of hydrogen production showed that Clostridia were the most abundant group with a proportion of more than 80%, while in seed inoculum it made <1%. 47 However, advanced studies revealed that hydrogen-producing microbial communities are characterized by predominance of Firmicutes phylum in general, including high diversity within Bacilli and Clostridia classes. 48,49 Proteomic analysis confirmed that Bacilli are active during the phase of hydrogen production and their proteins accounted 38.5% of Firmicutes' bacterial proteins. 38 Additionally, efficient hydrogen production from a wide range of organic substrates was shown for strains B. amyloliquefaciens, 50 B. firmus, 51 and B. licheniformis. 52 In our previous study, two strains affiliated with B. licheniformis and B. atrophaeus were isolated from fermentation liquid after digestion of food waste containing LCS substrate. 53 Therefore, we hypothesize that aerobic bacteria of lignocellulosic substrate might be affiliated with Bacilli class and comprise a significant part of microbial communities in the experimental sand mixtures.
The number of ammonifying bacteria found in the sand/LCS mixtures corresponds to that of highly fertile soils, while in sandy loam and long-term cultivated soils it is significantly lower (103–10 4 CFU/g). 54 B. cereus, B. subtilis, and B. megatherium showed ammonifying activity in agricultural soils, 55 and Paenibacillus species possess a wide range of metabolic pathways for nitrogen conversion, including ammonification. 56 The high number of gram-positive spore-forming rods in the sand mixtures can be also explained by the presence of lignocellulose. Introduction of crystalline cellulose into semi-desert soil caused a shift in a microbial community, with an increase in the proportion of Paenibacillus. 40
Interpreting the number of microorganisms growing on Ashby medium can be difficult. On the one hand, bacilli are able to grow in nitrogen-free environments and do not show nitrogen fixation. 57 On the other hand, bacilli are known diazotrophic endophytes, growing in the microenvironment of the root surface and supplying plants with nitrogen compounds. 58 We can conclude that bacteria of lignocellulosic substrates are able to grow at low nitrogen content and act as primary colonizers of sandy loam. However, the number of bacteria supplying mineral nitrogen for plant nutrition can be significantly lower than that of recorded in experiment.
The noticeable impact of microbial communities on R. sativus germination and resistance to drought stress can be also explained by the presence of Bacillus species in lignocellulosic substrate. Recent studies reported similar positive effect of Bacillus strains on Oryzae sativa at the early stages of growth. Inoculation of rice seeds with Lysinibacillus sphaericus and Bacillus cereus increases seed germination due to phytohormone production, 57 and a Bacillus sp. strain caused an increase of shoot length and content of dry matter in young rice seedlings. 59 Additionally, Bacillus comprise significant proportions of agricultural soil microbial communities in dry regions. 60,61 A native strain Bacillus megatherium, isolated from soil, increased resistance of white clover plants (Trifolium repens) to water stress. 62
Conclusion
Lignocellulosic substrate obtained during dark fermentation of food waste did not oppress shoot growth in lowest concentration (LCS-3, 33 g LCS/m2). It had a positive effect on root formation and significantly increased survival of the plants in stress conditions from short-term lack of water. Cultivation of R. sativus in the sand mixture containing 33 g LCS/m2 provided 100% seed germination and survival of 100% of seedlings under the two-day drought stress, while in plain sand these indicators were significantly lower. The number of aerobic ammonifying and free-living heterotrophic diazotrophic bacteria in LCS/sand mixture was similar to fertile soil. Preliminary results also suggest that hydrogen-producing microbial communities can be a source of Bacillus strains that may act as plant probiotics and supply mineral nitrogen for plant nutrition in arid conditions. Bacteria-saturated lignocellulosic substrate can be used potentially as a biofertilizer for sandy soil. However, further research is required to analyze microbial community composition and study the mechanism for the inhibition effect of high LCS content in sand mixtures.
Footnotes
Author Disclosure Statement
No competing financial interests exist.