
Abstract
The global clean energy supply has drawn tremendous attention toward alternative resources for biofuel. Microalgae are proving a potential source due to their high oil content and rapid biomasss production, but it still challenging to find potential species and produce biomasss at low-cost. The present work describes the sucessful collection, isolation, and identification of Mexican native microalgae species from eutrophic water. Morphological studies were conducted by analyzing the size and shape of pyrenoids, cell walls, nuclei, and mucilages with the help of light microscopy, and molecular analysis was performed using the internal transcribed spacer (ITS, 18–28S rRNA) gene partial sequence followed by phylogenetic analysis. Three species isolated from eutrophic water (Chlorella vulgaris, Chlorella saccharophila and Scenedesmus dimorphus), two standard culture media (Bold's Basal Medium (BBM) and Murashige and Skoog), and partially treated wastewater (TEC) were used in this study to select native potential microalgae for low-cost biomass and lipid production. The biomasss and lipid accumulation was nearly similar between BBM and TEC. It was concluded that it is feasible to use wastewater (TEC) to cultivate these native algae to treat wastewater for removal of nutrients and produce lipid at a lower cost than standard medium. Furthermore, the native species can be used without seasonal acclimatization. It is necessary to conduct further experiments to optimize the culture condition and maximize biomasss production in order to scale the production to an industrial level.
Introduction
Since the oil crisis of the 1970s, research on microalgae—especially those which accumulate lipids—has been increasing. Lipids from microalgae can be extracted and transformed into biodiesel, which can be used in conventional diesel engines 1 –3 and with some additives. 4,5 Biodiesel can be defined as fatty acid monoesters produced from animal fat or vegetable oil (edible or not) through a transesterification process. Microalgae may play an important role in biodiesel production due to their high growth rate and their ability to accumulate high lipid quantities. 6 However, Sheehnan et al. 7 concluded that scaling microalgae culture for biodiesel production was not economically profitable due to culture medium cost and recommended culturing microalgae in combination with wastewater treatment. Since then, many studies have been focused on comparing biomass production and lipid accumulation in microalgae using different types of wastewater (urban, dairy, and crop effluents) 8,9 with different types of treatment and concentration. 10 –12 Kong et al. 13 cultivated Chlamydomonas reinhardtii in municipal wastewater with the productivity of 2 g/L/d and 25.25% of lipids (dry weight). This work suggests that when using treated wastewater, even the nitrogen and phosphorus concentration in raw wastewater can inhibit growth. Yang et al. 14 used municipal wastewater treated by activated sludge to culture Chlorella ellipsoidea YJ1 and produced 425 mg/d of biomass (dry weight), with 43% lipid content. Bhatnagar et al. 10 also affirm that, in order to lower costs in the microalgal biodiesel process, it is necessary to use wastewater as a free source of nutrients like N and P, and CO2 from bacterial activity. For this, they cultured Chlorella minutissima in Diluted Domestic Wastewater (DDW) and concluded that C. minutissima increased biomass production by 50% with DDW compared to BG11 artificial medium, cutting down the production expenses.
Farm wastewaters are also full of nutrients, which in many cases trigger eutrophication in water bodies if not treated. This excess of nutrients can be used to cultivate microalgae. Woertz 12 used dairy wastewater to cultivate microalgae species Chlorella, Scenedesmus, Micractinium, and Actinastrum, getting up to 915 mg/d of biomass (dry weight) and 29% of lipids in mixed cultures. Olguin et al. 15 cultured Spirulina platensis in pork wastewater diluted with sea water under different light intensity and found that highest biomass productivity (38 mg more than artificial medium) was at 144 μmol photon/m2/s with a biomass production of 0.77 g/L by using sea water. However, the highest percentage of lipids (28.8%) was found at 66 μmol photon/m2/s.
Many species are closely related morphologically, which can be observed among Chlorella species due to Brachionus–factors 16 and among Scenedesmus due to Daphnia–factors 17,18 and Brachionus–factors. 3 We also found the same problem with our previous algal identification. 19,20 It is therefore necessary to identify the species using genetic analysis, and molecular characterizations based on internal transcribed spacer (ITS) region used for taxonomic studies is an effective tool for identification up to the species level. 21 –24
In this work, we have attempted to isolate, identify taxonomically and molecularly, and characterize local algal strains from eutophic water and evaluate their potential for lipid production by growing them directly on partially treated wastewater in order to make an economically profitable process at an industrial scale for wastewater treatment and biomass and lipid production.
Materials and Methods
Microalgae Isolation
Microalgae was collected and isolated from two points near Guadalajara, Mexico in March 2010. The first location, an irrigation canal that carries water from the Santiago river, (20°32'48.84”N; 103° 9'9.75”W) is known for its high pollution levels caused by industrial and farming activities—so the treatment plant collects water from several sources of household and industrial waste. The second location was a water-treatment plant near the small town of Ixtlahuacán (20°24'34.38”N; 103°07'56.95”W). Samples were then concentrated by centrifuge and cultured in liquid and solid media. In order to isolate microalgae single colonies using solid medium, a series of dilutions were made. 100 μL of each dilution flask was inoculated in Petri dishes with solid Combo 25 and Murashige and Skoog (MS) 20 medium. Cultures were kept at 27°C under white fluorescent lamps (150 μE/m2) for periods of 16 h light:8 h dark. After 20 d, visible isolated colonies were transferred into separate flasks with liquid medium and cultured under the same conditions. Microscopic observations were made in order to assure mono algal cultures. If two or more species were observed, the isolation process was repeated until mono algal cultures were obtained.
Taxonomic Determination
To determine the taxonomic division of the isolated strains, LucidCentral (Stafford Heights, Australia) keys were used to preliminarily compare morphological characteristics. Then, the samples were stained with Lugol, Cresil Blue, and Bright Green to observe starch in pyrenoids, cell walls, nuclei, and mucilages under an optical Olympus (Tokyo, Japan) BH-2 microscope coupled to a digital Leica (Wetzlar, Germany) DFC 450C camera. The number of cells per coenobium, cell shape, and cell arrangement were documented, and taxonomic identifications were performed. 5,26
Molecular Identification
Genomic DNA from three algal strains was extracted according to a previously described method, with minor modifications.
27
The 18–28S ribosomal RNA internal transcribed spacer (rRNA ITS) region of all three algal strains was amplified using specific primer pairs ITS1 (
Amplified PCR products were subjected to agarose gel electrophoresis and visualized using a UV transilluminator. The specific products of PCR directly as well as the products of PCR from the agarose gel were gel purified using kits according to the manufacturer's instructions (Qiagen, Hilden, Germany) and sequenced. The DNA sequences were viewed and edited using Sequence Scanner, (Applied Bioscience). A global BLAST search was then performed to identify the most closely related sequences from the NCBI database using PUBMED nucleotide Blast. The local blast and multiple sequence alignments were compiled and analyzed using BioEdit v7.2.5 (Ibis Biosciences, Carlsbad, CA), and the phylogenetic analysis was performed using software MEGA 6. 29
Microalgal Growth Measurement
For growth measurement experiements, three culture media were used: Bold Basal Medium (BBM), 30 MS, and treated water from the Tecnologico de Monterrey, Campus Guadalajara domestic wastewater treatment plant (TEC). The water was taken after a biological treatment consisting of an active sludge system followed by a clarifier before chlorination. The composition of the two media is described in Table 1. BBM medium was prepared with distilled water and, along with TEC water, adjusted to 6.6 pH before sterilization in an autoclave. For microalgae cultures, the flask was adjusted to 0.05 OD in 250 mL flasks with 100 mL of culture medium. Cultures were kept under the same conditions as cultures in the solid medium, plus agitation at 150 rpm. Growth measurements were taken daily at the same time (680 nm) and the correlation between the optical density and dry weight was calculated. The biomas was also collected from the same culture for lipid measurements.
Culture Media Components (mg/L)
BBM: Bold Basal Medium; bTEC: Water from Tecnologico de Monterrey, Campus Guadalajara municipal wastewater treatment plant.
Lipid Contents Measurements by Red Nile Fluorescence
Red Nile fluorescence measurement was used to determine the lipid content of above algal liquid cultures, and only BBM and TEC media were compared at this stage. 2-mL samples were taken from each strain and each medium and stored at −80°C for further lipid analysis. Then, 8μL of Red Nile solution (250 mg/L) was added to previously defrosted samples. After 30 sec of vortex agitation and 10 min rest, fluorescence was measured at 480 nm excitation/580 nm de emission. 31,32 All measurements were carried out in triplicate. In order to have enough biomass for lipid extraction, cultures were established under the same conditions as described above. Once lipid accumulation reached the highest peak, determined by the fluorescence analysis, algal biomass was collected and lyophilized.
Extraction and Estimation of Lipids
The lipids were extracted following Feng et al. 33 To 150 mg of dry algal biomass, 12 mL of chloroform and 6 mL of methanol was added. After vortex agitation for 1 min, 6 mL of methanol and 10.8 mL of distilled water were added to a final ratio of 2:2:1.8, ending with vortex agitation for 1 more min and centrifugation at 5,000 rpm for 10 min. Three layers resulted: an upper layer containing water and methanol, a middle layer containing chloroform with lipids, and a lower layer containing pelletized biomass. The upper layer was discarded, and the chloroform layer was transferred to a clean, pre-weighed vial. The procedure was repeated twice to the lower, pelletized biomass layer for a complete lipid extraction. The three chloroform layers containing lipids were mixed, and the solvent was evaporated in order to obtain pure lipids. The percentage (w/w) of lipids was quantity and calculated.
Results
Microalgae Isolation and Taxonomic Determination
During isolation, several multicellular and unicellular algae were found in the sampled eutrophic water. Only three specices were considered for further study on the basis of growth and prevous studies.
C. vulgaris was identified based on the morphological characters,
5,26
such as unicellular, spherical cells measuring between 2.6 and 9.8 μm in diameter and bearing a thin cell wall; cup-shaped chloroplast; and 2–4 starch grains in the pyrenoid (Supplementary Fig. S1A,B Supplementary Data are available online at
Molecular Identification
The internal transcribed spacer of the 18–28S rRNA gene (ITS) region was successfully amplified from three species C. vulgaris, C. saccharophila, and S. dimorphus. The resulting PCR products of the ITS from C. vulgaris and C. saccharophila were found to be nearly equal in size (760 bp), whereas the ITS region of S. dimorphus was smaller (700 bp, data not shown).
Phylogenetic analysis was conducted using the sequences of ITS regions obtained from all three strains under study. The sequences were compared with several reference sequences (including the ITS region of several Chlorella and Scenedesmus species) obtained from the NCBI database. Phylogenetic tree analysis of Chlorella and Scenedesmus strains fell into four distinct clusters and several sub-clusters. The C. saccharophila was found to be very close sequentially and formed clusters along with all other C. saccharophila; S. dimorphus formed clusters with S. dimorphus and S. obliquus; and C. vulgaris forms a separate sub-cluster a short distance from all other C. vulgaris. The C. saccharophila of this study was very close to SAG (Germany) and UTEX (USA) algal collections, while S. dimorphus most closely met with UTEX. C. vulgaris was different from UTEX collections but close to CBS 15–2015, of CCAP (UK), as seen in Fig. 1.

Molecular phylogenetic analysis; the tree with the highest log likelihood (−7467.8402) is shown. Initial tree(s) for the heuristic search were obtained by applying the Neighbor-Joining method to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 37 nucleotide sequences. There was a total of 867 positions in the final dataset. Evolutionary analyses were conducted in MEGA6.
Microalgal Growth Measurements
In order to measure biomass and lipid content, a correlation between optical density and dry weight was performed (Supplementary Table S1). The simple regression analysis were performed to evaluate the correlation using Statgraphics. The accumulation of biomass was monitored in Chlorella vulgaris in three different culture media up to 11 days (TEC, MS and BBM) (Fig. 2A). The maximum biomass acumulation was achieved on the seventh day, and thereafter there was no significant increase in biomass in all the media studied. The highest biomass concentrations were registered in the TEC and BBM culture media (0.252 and 0.257 g/L, respectively) without any significant difference between them (P = 0.4671, α = 0.05), whereas biomass acumulation in MS medium was minimal (0.221 g/L) (Fig. 2A). The biomass accumulation was monitored in C. saccharophila in different culture mediums (TEC, MS, and BBM) up to 11 days. In Fig. 2B, the accumulation of biomass of C. saccharophila showed that there were no significant (p = 00-05) differences among the three studied media except on day 7 in which the growth was slightly less in MS medium. The highest accumulation of biomass was observed in the TEC medium, with 0.412 g/L followed by BBM and MS, with 0.393 and 0.319 g/L, respectively. It may be possible due to the types of nutritional requirement and differences in the structure of the cell wall in C. saccharophila, with respect to C. vulgaris. The biomass accumulation of S. dimorphus is shown in Fig. 2C. The stationary phase of this algal culture was reached on day 4 in both the TEC and MS media and on day 5 for the BBM medium. The culture medium TEC reached the stationary phase with a biomass accumulation of 0.107 g/L, with MS medium at 0.046 g/L and 0.156/g L for BBM. A clear and significante difference was observed in biomass between TEC and MS medium (P = 0.0109-value, α = 0.05). On the other hand, growth in BBM medium reached a higher biomass accumulation on day 5 (P = 0.0337, α = 0.05). However, on day 4, there was no significant difference with the TEC medium (P = 0.3196, α = 0.05).
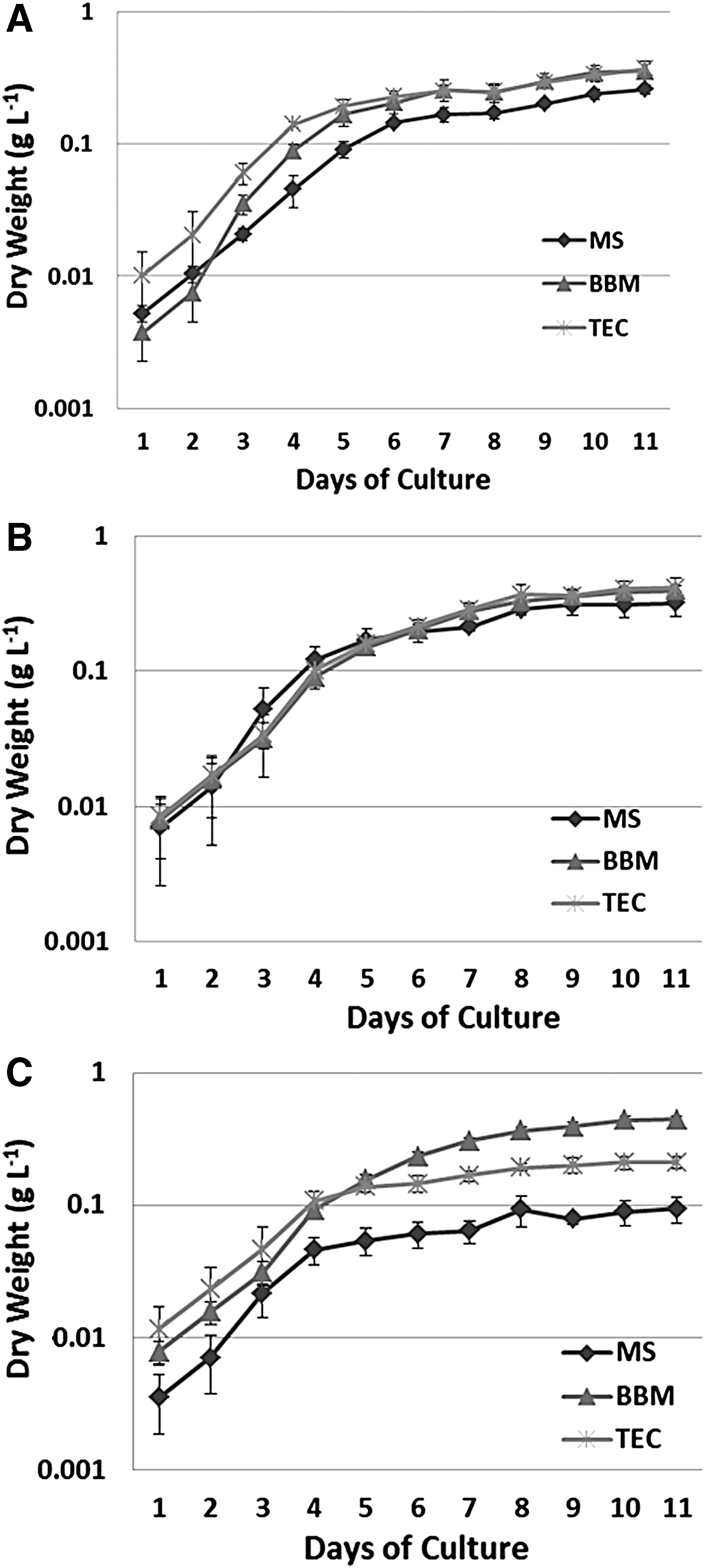
Biomass accumulation in different culture media;
Lipid Contents by Red Nile Fluorescence Measurements
Cells of C. vulgaris were stained with Nile Red and showed a slight increase of lipid production among the culture period observed by fluorescence (Fig. 3A). In the TEC medium, In the BBM medium, a decrease was observed until the second day, followed by an increase to reach a maximum at day 12. The lipid accumulation of C. saccharophila is shown in Fig. 3B. It was observed that the increase in lipid concentration starts on the second day. However, the accumulation was more prominent when the culture reaches the stationary phase on day eight. For S. dimorphus, Fig. 3C shows lipid accumulation was noticed at the end of the exponential phase. The highest accumulation of lipids in the TEC medium was observed on the 9th day, whereas in the case of BBM it was on the 12th day. The fluorescence values for this strain were lower than those of the previous strains.
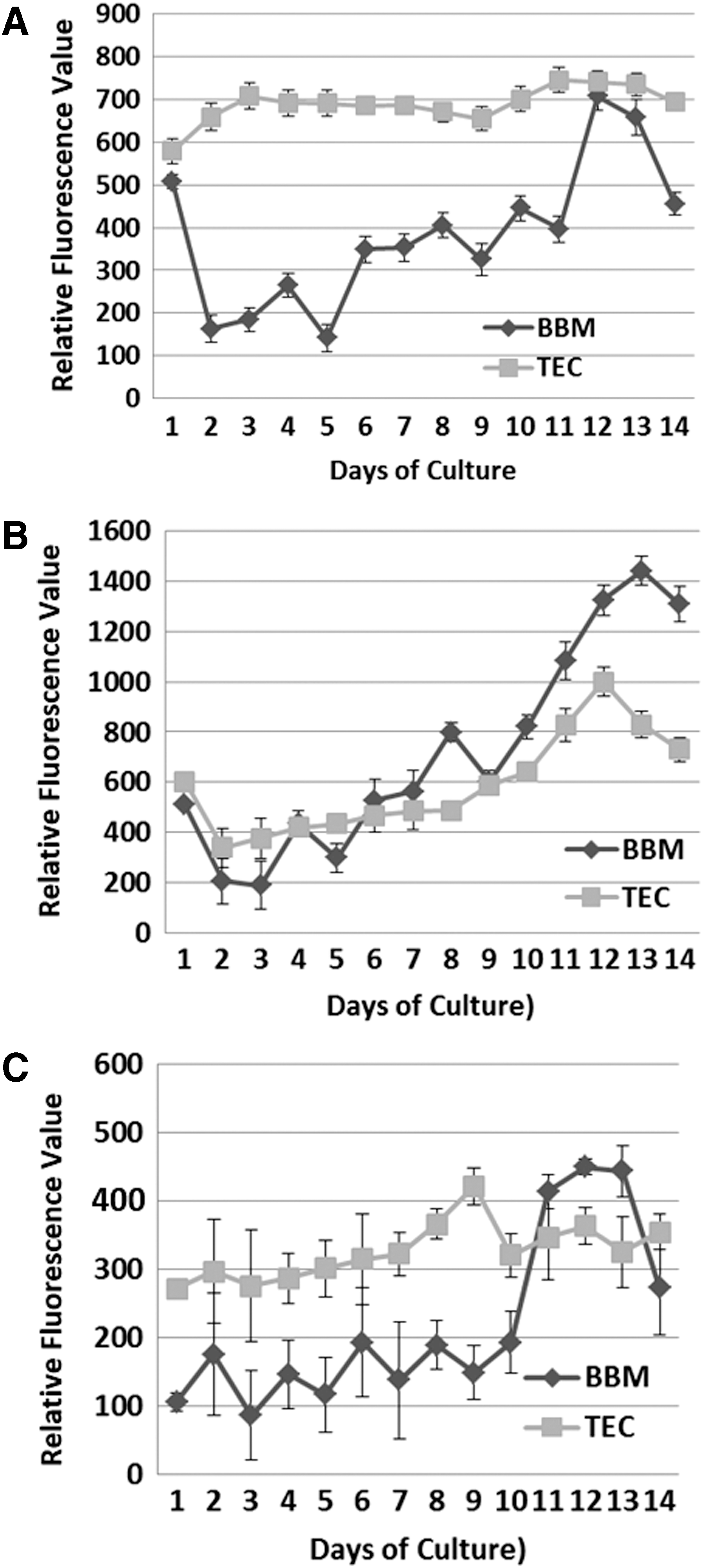
Accumulation of lipids quantified by Nile Red fluorescence;

Comparative biomass accumulation representation of Chlorella vulgaris, Chlorella saccharophila and Senedesmus dimorphus in three different culture media on the 11th day.
Extraction and Estimation of Lipids
Finally, total biomass was harvested on the 11th day, and total lipid content was estimated in both culture media. The results of lipid contents (% w/w) are shown in Fig. 5. C. vulgaris shows a higher accumulation of lipids in the BBM medium (25.2%) compared to medium TEC (19.5%), although no statistically significant difference was detected (P = 0.148195, α = 0.05). In C. saccharophila, the accumulation of lipid was slightly higher in the BBM medium by 24.37% in comparison to TEC medium with a content of 23%. However, lipid concentrations were not significantly different (P = 0.713, α = 0.05) (Fig. 5). S. dimorphus also observed the same tendency, the total lipid content in the TEC medium was slightly higher, with 24.56% lipid than that in the BBM medium with 19.79%, although there were no significant differences between them (P = 0.1424, α = 0.05) (Fig. 5).
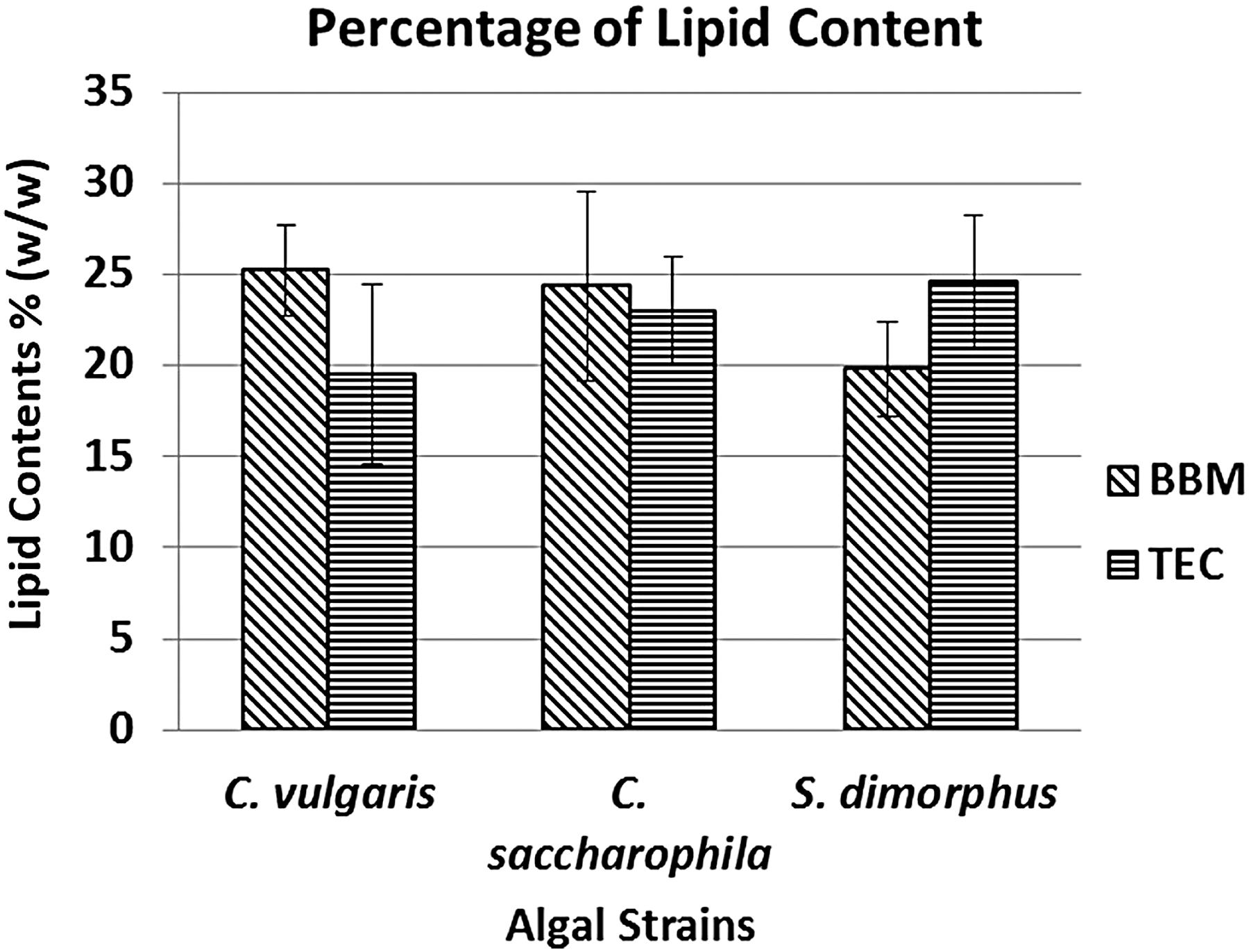
Comparative percentage of lipid content representation in Chlorella vulgaris, Chlorella saccharophila, and Senedesmus dimorphus in two different culture media on the 12th day.
Conclusion
Morphological variations are very high among Chlorophytes in normal growth conditions and even higher in the influence of various factors. 16,18,35 According to the observation of the number of cells per coenobium, cell shape, and cell arrangement (Supplementary Fig. S1), the three isolated strains were identified taxonomically 5,26 and molecularly as C. vulgaris, C. saccharophila, and S. dimorphus 21,23,24,36 as as per their sequential similarity in ITS region.
In the TEC medium C. vulgaris maintained the lipid concentration throughout the culture for 12 days (Fig. 3A); this may be due to a high concentration of organic carbon 37 and/or a low amount of nitrogen in the medium. 38,39 In the case of C. saccharophila, increase in lipid concentration starts from the second day and after the 10th day for S. dimorphus. However, the accumulation was more prominent when the culture reaches the stationary phase in all three cases. This may be due to reduced nitrogen and other nutrients in the medium. 37 –41 The fluorescence values for S. dimorphus were lower than those of the other two strains (Fig. 5), perhaps because the cell wall of Scenedesmus has a more complex structure and contains, besides cellulose, sporopollenin, which inhibits the total diffusion of Nile Red. 42
The percentage of lipid extracted on the final biomass content was not significantly different (Fig. 5) among the three species under study, although the Nile red staining fluorescence values were quite different (Fig. 3A–C). The Nile red fluorescence value is sometimes species-specific due to the complexity of cell wall, 42 and it may not give a comparable fluorescence value among species, but rather be a better tool for single species studies. In Table 2 3,33,38,43 –46 the results obtained by C. vulgaris and C. saccharophila in this study are compared with data obtained from other investigations. As seen in this table, the results obtained by C. vulgaris and C. saccharophila in this work were very close to each other, in some cases higher than the findings of other researchers.6,8,33,38,47 Kim et al. 22 studied Scenedesmus sp. in fermented wastewater (swine urine water treated with bacteria through fermentation and bioreaction technologies) and found a greater biomass accumulation in the control medium (BBM) but a lower percentage of lipids, while the present study showed the opposite. This can occur because the fermented wastewater used by Kim et al. 22 have higher levels of nitrogen and phosphorus than those found in the wastewater of this study (nitrogen: 86.4 and 57.6; phosphorus: 20.2 and 4.26 mg/L, respectively), suggesting that culture media rich in these elements accumulate greater biomass but a lesser percentage of lipids. On the other hand, the 0.107 g/L of biomass obtained in TEC is not as high compared to biomass levels commonly obtained (0.3–0.5 g/L) in cultures of microalgae. 48 This could be due to the low concentration of phosphorus in the TEC medium 14 and without supplementary CO2. The supplementation of phosphorus may function like the complete synthetic medium. These local strains appear promising for testing for industrial-scale biomass and lipid production coupled with waste water treatment.
Accumulation of Biomass and Lipids Content of C. vulgaris and C. saccharophila in Different Studies Under Autotrophic Culture (Except When Othewise Indicated)
The present laboratory-scale work demonstrates that native microalgae strains isolated from eutrophic water bodies—C. vulgaris, C. saccharophila, and S. dimorphus—can accumulate the same amount of biomass using treated water from domestic Tec de Monterrey wastewater treatment plants as in artificial culture medium (BBM). Furthermore, levels of lipid accumulation showed no significant differences compared to other studies. Fatty acid profiles and their biofuel applications are well-recognized for these genera. Therefore, the treated wastewater can be used successfully in the production of biomass and lipids of these strains. Further studies are needed to show if in this same process the domestic water can be treated using these microalgae for the removal of nutrients and other contaminants. As these genera are also recognized for protein and other feed value, coupling next scale with production of raw material for the feed industry could be considered.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.