
Abstract
Cellulose is the most abundant renewable carbon source found in nature and is of great interest to various industries, including the food industry. Generally, microorganisms can be used to convert cellulosic materials into higher protein content, which can subsequently be used for animal feed. However, without the aid of cellulase, the cellulose structure is resistant to degradation. In this study, genome shuffling was used to improve cellulase production from Aureobasidium pullulans Y-2311-1. One strain developed via genome shuffling (A. pullulans GS23) displayed the largest increase in total cellulase activity on day one, which was a 6-fold increase compared to the wild-type strain. The A. pullulans GS23 strain also had an increase in exoglucanase and β-glucosidase activity compared to the wild-type strain (6.95-fold and 1.52-fold increase, respectively). The crude protein amount of A. pullulans GS23 had a 1.04-fold increase compared to the wild-type strain after 5 days of fermentation. As A. pullulans GS23 has increased cellulase production compared to the wild-type, this strain has the capability to develop improved, protein-rich animal feed by decreasing the amount of commercial enzymes used in the fermentation process. This is the first known attempt to increase cellulase with genome shuffling in A. pullulans.
Introduction
Cellulose is the most abundant renewable carbon source found in nature. The abundance of cellulose has generated considerable interest from researchers interested in converting cellulosic materials into protein-rich cell biomass for animal feed to increase the initial protein concentration in lignocellulosic biomass. 1,2 Cellulose is a β-1,4-linked glucose polymer with a physical and morphological structure that creates complex structures within higher plants, resulting in plant material that is difficult to degrade. 1,3 The crystalline structure of cellulose requires supplementary enzymes for most microorganisms to effectively degrade cellulose into fermentable sugars. Three enzymes that can degrade crystalline cellulose into glucose are collectively called cellulases. They work together to hydrolyze lignocellulosic biomass and are named endoglucanase (endo-1, 4-β-D-glucanase, EC 3.2.1.4), exoglucanase (1,4-β-D-glucancellobiohydrolase, EC 3.2.1.91), and β-glucosidase (β-D-glucoside glucanohydrolase, cellobiase, EC 3.2.1.21). 4,5 Endoglucanases randomly cleave internal bonds in a cellulose chain, while exoglucanases cleave the ends of the chains to release cellobiose. β-glucosidase is then activated when cellobiose is available and releases glucose monomer units. 6 These enzymes can be purchased commercially and added to the fermentation vessel, however, the major limiting factor of commercial cellulases is cost. Some strains of fungi and bacteria are known to metabolize complex cellulosic substrates by producing cellulase that can then convert fermented sugars into single cell proteins (SCPs). SCPs are produced from biomass by microbial sources and have been used as a protein supplement in animal feeds. A wide range of microorganisms (algae, bacteria, fungi) is known to produce SCPs using carbon- and nitrogen-rich agricultural and industrial wastes. 7 The advantage of using fungal strains industrially to degrade cellulosic material is that fungal strains generally contain the complete set of cellulosic enzymes, unlike bacteria. 7,8 A fungal strain that can produce effective amounts of cellulase on its own could replace the need of adding commercial cellulases to the fermentation process. Therefore, it is important to improve the production of cellulase from microorganisms that naturally produce the enzymes to make the production of increased protein levels in lignocellulosic biomass economically feasible without the added cost of commercial enzymes. 9
Our research group has previously used various strains of Aureobasidium pullulans to produce increased protein levels from several feedstocks. 10 -12 The initial industry applications of A. pullulans have been the production of pullulan, which can be used as a thickening or extending agent, adhesive or encapsulating agent, or oxygen-impermeable film and fiber. 13 When A. pullulans is grown on a solid surface (i.e., agar-based medium), the initial visible colonies can be yellow, cream, light pink, or light brown, depending on the strain. As colonies age, they typically darken to a blackish color due to the production of melanin. Therefore, A. pullulans is commonly known as the black yeast. 14,15 A. pullulans is a pleomorphic fungus that can live in diverse habitats 16 and has the ability to respond to environmental signals by altering its morphology, physiological state, or behavior, which is termed phenotypic plasticity. 17 The three morphological forms A. pullulans can differentiate into are large chlamydospores, smaller elliptical yeast-like cells, and elongated branched septate filaments that can grow at low water activities. 15,18 A. pullulans has a relatively low content of nucleic acid, unlike bacteria, which is an attractive feature. 19 Intake of SCPs with high levels of nucleic acid, like any other essential dietary ingredient, can cause physiological effects. 7 The versatile properties of A. pullulans make it an ideal candidate to be cultivated on various cellulosic materials to create fermentable sugars. Cultures of A. pullulans that have shown promising cellulolytic activities (endoglucanase and exoglucanase) originate from tropical regions. 2,20 -22 Therefore, the strain chosen for this experiment was the tropical isolate A. pullulans Y-2311-1. 23 One trait that makes this strain industrially favorable is that it is known to secrete high levels of xylanolytic enzymes into culture media. 21 Xylan is the major constituent of hemicellulose, which is the second most abundant renewable biomass after cellulose. 24,25 Wild-type A. pullulans Y-2311-1 does produce cellulase, but at a low concentration. 26 Thus, the current challenge is to increase the cellulase production of the microorganism, which would allow for more efficient cellulose utilization.
Improving industrially relevant phenotypes using recombinant DNA techniques in microorganisms has proven to be difficult. One way to overcome this deficiency is through the use of genome shuffling. This process combines breeding procedures with rapid screening to quickly improve microbial phenotypes by simultaneously changing genes positions throughout the entire genome without the need to know the genome sequence. 27 -29 Genome shuffling has successfully been used on a variety of microorganisms to improve different applications, including: acid tolerance in Lactobacillus, 28 degradation of pentachlorophenol by Sphingobium chlorophenolicum, 30 hydroxycitric acid production in Streptomyces, 31 thermotolerance, ethanol tolerance, and ethanol production in Saccharomyces cerevisiae, 32 pullulan production in A. pullulans, 33 and cellulase production in Trichoderma viride. 34 The specific aim of this study was to assess if genome shuffling can be used in A. pullulans Y-2311-1 to generate the strains with improved cellulase production capacity. Hence, a shuffled strain of A. pullulans Y-2311-1 was developed via genome shuffling by screening a library for an increased cellulase production phenotype. Developing an improved strain would alleviate the cost of using commercial cellulases in fermentations and potentially lead to an industrial cellulosic process that is economically feasible.
Materials and Methods
Microorganisms and Culture Materials
A. pullulans NRRL Y-2311-1 (known hereafter as A. pullulans) was provided by the USDA National Center for Agricultural Utilization Research (Peoria, IL). It was cultured in 5% glucose yeast extract growth medium (GYE, w/v, 5% glucose, 0.5% yeast extract) and maintained on potato extract agar (PDA, w/v, 2.4% potato extract broth, 1.5% agar). To develop mutants with increased cellulase production, A. pullulans was cultivated in carboxymethylcellulose broth (CMC broth, w/v, 0.125% CaCl2, 0.0625% (NH4)2SO4, 0.0625% yeast extract powder, 0.0625% asparagine, 1.25% carboxymethylcellulose, 0.125% KH2PO4) and mutants were selected on carboxymethylcellulose agar (CMC agar, w/v, 0.5% carboxymethylcellulose, 0.1% NaNO3, 0.1% K2HPO4, 0.1% KCl, 0.05% MgSO4, 0.05% yeast extract, 0.1% glucose, 1.7% agar).
The solid-state fermentation medium was prepared by a method adapted from Xu et al. 34 Soybean hulls were used as the cellulosic substrate, acting as both a carbon source and inducer of cellulase production. 35 The medium was prepared in 250-mL Erlenmeyer flasks containing 6.25 g of soybean hulls, 0.03 g (NH4)2SO4 (nitrogen source), 6.25 mL of distilled water (dH2O), and pH adjusted to 5 with 3.6 N H2SO4. The solid-state fermentation medium was autoclaved for 20 min at 121°C.
The submerged-state fermentation medium was prepared using 2 mm ground corn bran. Corn bran was selected for the carbon source in accordance with Saha, Freer and Bothast, 36 who found it to be effective at expressing β-glucosidase from another strain of A. pullulans. The medium was prepared in 250-mL Erlenmeyer flasks containing 5 g of corn bran, 95 mL of dH2O, and pH adjusted to 5 with 3.6 N H2SO4. The submerged-state fermentation medium was autoclaved for 20 min at 121°C. Glucose and CMC were purchased from Sigma-Aldrich (St. Louis, MO). Yeast extract and chemicals (analytical grades) were procured from ThermoFisher Scientific. DifcoTM potato dextrose agar was purchased from BD Biosciences.
Mutagenesis
Mutant populations of A. pullulans with potentially enhanced cellulolytic capabilities were developed from wild-type A. pullulans. These developed strains were used as starting strains for the genome shuffling protocol. The mutants were considered improved when the cellulase content was significantly increased compared to the wild-type strain as determined by a two-tail t-test (α = 0.05). The treatments used to develop these mutants were short wavelength ultraviolet (UV-C) radiation and methyl methanesulfonate (MMS) and are further described below. The mutagenic agents' lethality was measured, and the D-values were calculated from the survival curves. The D-value is the time required to kill 90% of exposed microorganisms to ensure damage has occurred, particularly to the DNA. This method has commonly been used in mutagenesis studies. 37 -39
The ultraviolet (UV-C) treatment methodology was adapted from Kang et al. 33 For this treatment, A. pullulans was grown in 100 mL CMC broth in a 250-mL Erlenmeyer flask for 3 d. Then, 5 mL of the culture was transferred to an aseptic Petri dish and exposed to UV-C irradiation with the lid removed for 40 min (D-value). The UV source was a Spectroline Fluorescence Analysis Cabinet CX-Series (Westbury, NY). It contained a rear and front UV lamp pointed toward the middle of the cabinet. The Petri dish was ∼16 cm away from the UV lamps and visible light exposure was kept to a minimum when processing the sample after UV exposure. The reason for limiting visible light exposure was to decrease the chances of activating photoreactivating enzymes, which repair DNA damage and negate mutagenesis efforts. 40
The MMS treatment methodology was adapted from Lindegren et al. 41 and Wu et al. 42 A. pullulans was grown in 100 mL CMC broth in a 250-mL Erlenmeyer flask for 2 d. Then, 5 mL of the culture was transferred to a sterile, 15-mL conical and centrifuged at 4,000 rpm for 10 min at room temperature. The cells were then washed once with 5 mL of sterile dH2O and centrifuged at 4,000 rpm for 10 min at room temperature again. The washed cells were resuspended in 2 mL of 25 mM Tris-HCl buffer, pH 6. In a separate aseptic tube, 0.1 mL of cell suspension was added to 0.8 mL of 0.1 M potassium phosphate buffer, pH 7. Then 0.1 mL of 20% glucose and 0.01 mL of MMS was added to the cell suspension and potassium phosphate buffer, pH 7 mixture. The tube was mixed and placed in a static incubator at 30°C for 35 min (D-value). After the mixture was taken out of the incubator, 5 mL of 6% sodium thiosulfate was added to inactivate the MMS. After 10 min of exposure to the sodium thiosulfate, the cells were centrifuged at 4,000 rpm for 10 min at room temperature. The 6% sodium thiosulfate treatment was repeated to completely inactivate the MMS before the resulting cell pellet was resuspended in 1.5 mL of 25 mM Tris-HCl buffer, pH 6.
Genome Shuffling
The genome shuffling methodology was adapted from Hida et al. 31 and Kang et al. 33 The complete scheme of genome shuffling is presented in Fig. 1, including mutagenesis. An A. pullulans mutant strain selected from the UV treatment and MMS treatment were grown separately in 100 mL CMC broth in a 250-mL Erlenmeyer flask for 6 d. Then, 4 mL of the UV and MMS strains were harvested separately and centrifuged at 4,000 rpm for 10 min at room temperature. The pellets were washed twice with 1.5 mL of hypertonic solution I (0.5 M KCl, 25 mM Tris-HCl buffer pH 6) by centrifuging at 4,000 rpm for 10 min at room temperature. The washed pellets were then treated with 4 mL of lysozyme (10 mg/mL in hypertonic solution I) and put into a 35°C static incubator to allow the protoplast to form (protoplasts occur when the fungal cell wall is partially removed). Protoplasts were created to allow efficient genetic transfer between the UV and MMS strains. To check for protoplast formation, wet mounts were prepared with 0.1 mL sample and stained with methylene blue before being viewed under a Leica ATC 2000 compound light microscope (Leica, Buffalo, NY). After the protoplast had formed for both UV and MMS strains, half of each UV and MMS protoplast strains' cultures (2 mL) were mixed together in a 15-mL aseptic tube. The 4 mL of mixed protoplasts were then manually fractionated into two 2 mL aliquots. One aliquot was transferred into an aseptic Petri dish and exposed to UV radiation for 40 minutes (D-value) and the other aliquot was heat-treated in a static 60°C incubator for 44 min (D-value) in an aseptic tube. Protoplast inactivation by two different methods before fusion was to prevent the formation of parental-type clones by hindering their ability to divide. However, to reactivate the protoplasts prior to fusion, inactivated protoplasts formed from a different treatment were needed. 43 Thus, the next step involved mixing both aliquots together and centrifuging at 4,000 rpm for 10 min at room temperature. The pellet was resuspended in 1 mL of hypertonic solution II (0.5 M sucrose, 25 mM Trish-HCl buffer pH 6) and then treated with 9 mL of 35% PEG 6000 and 0.1% CaCl2. The mixture was incubated for 40 min in a 35°C rotatory shaker at 100 rpm. The fused protoplasts were then centrifuged at 4,000 rpm for 10 min at room temperature and washed twice with 1.5 mL of hypertonic solution II using the same centrifugation parameters as described above. The resulting cell pellet was resuspended in 1.5 mL of hypertonic solution II. This protoplast fusion technique leads to a stable recombinant with the combination of genetic traits of both parents. 44
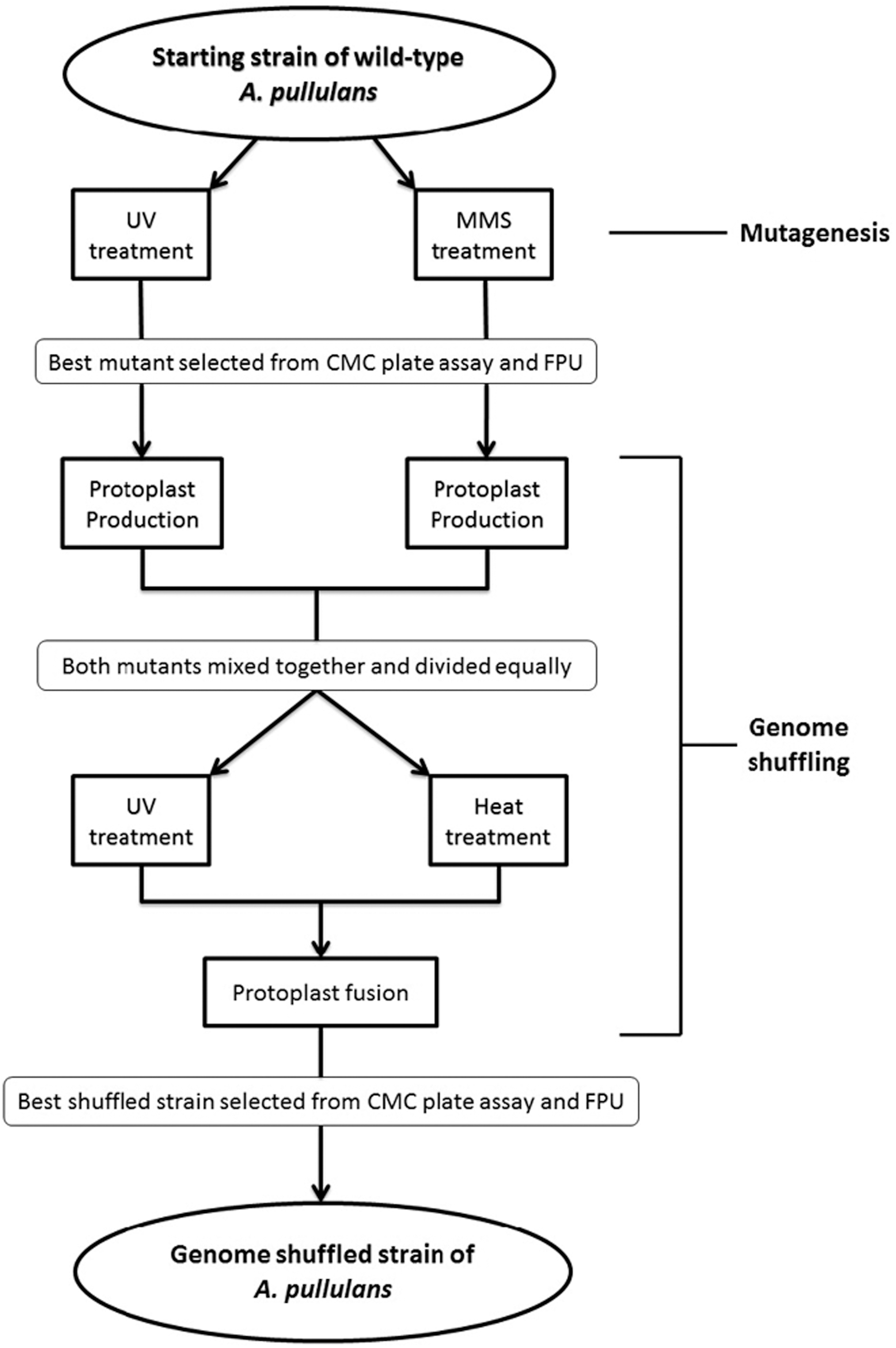
The complete procedure of genome shuffling for improving cellulase production of A. pullulans.
CMC Plate Assay Screening
Cultures subjected to UV treatment, MMS treatment, and genome shuffling were screened to determine the strains' capability of cellulolytic degradation via cellulase activity by performing CMC plate assays with Gram's iodine as adapted from Kasana et al. 45 When each treatment was completed, the sample was serologically diluted in 1 M phosphate buffer and spread evenly on CMC agar plates. The colonies were incubated on the CMC agar plates for 4 days at room temperature. Each colony was then transferred to two new CMC agar plates (duplicates) by selecting a colony and stabbing it straight down into the agar with a sterile toothpick for each plate. Next, the colonies were spaced evenly on the plates (12 colonies per plate) and were incubated for 4 days at room temperature before performing a CMC plate assay with Gram's iodine. One of the duplicated CMC agar plates was flooded with 50% v/v of Gram's iodine for 5 min and then rinsed with dH2O. The colonies with the largest clear zones (measured from edge of colony to edge of zone) were selected and taken from the duplicated plate with no Gram's iodine for enzyme assay analysis. As Gram's iodine is toxic to microorganisms, the duplicate plates were necessary to maintain surviving mutants.
Enzyme Assays
Crude enzymes of the wild-type, mutant, and genome shuffled strains were prepared to detect cellulase as described by Latifian et al., 46 with modifications. Three mL of a four-day culture of each strain previously selected by the CMC plate assay, and wild-type strain, were evenly pipetted onto the surface of the soybean hull medium in 250-mL Erlenmeyer flasks. Soybean hulls were chosen to induce cellulase due to its rich cellulosic composition. Half of the soybean's composition is cellulosic, including cellulose and hemicellulose. 47 The flasks were incubated in a 30°C static incubator before adding 5-fold (62.5 mL) of sterile dH2O to the 12.5 g soybean hulls in the soybean hull medium. The mixture was then homogenized for 10 min with a VirTis VirTishear Model 225318 homogenizer with 115 voltages, 5 amps, and 50/60 Hertz (Gardiner, NY) set at 45 arbitrary units. The mixture was centrifuged twice at 5,000 x g for 10 min and the resulting supernatant was used as the source of crude enzymes.
Filter paper units (FPU, representing total cellulase activity) was determined using the methods described by Ghose
48
with modifications provided by Dr. Lu-Kwang Ju (The University of Akron) via personal correspondence. For each strain that exhibited potential cellulase enhancement in the CMC assay, 0.1 mL crude enzyme was added to 1.4 mL of 0.05 M sodium citrate buffer, pH 4.8. Dinitrosalicylic acid (DNS) was prepared by dissolving 10 g of DNS in 600 mL of dH2O. Then, 16 g of NaOH was gradually added and allowed to dissolve before slowly adding 300 g of Rochelle salt (potassium sodium tartrate) over 20–30 min. After the solution was completely dissolved, it was diluted with dH2O to 1 L. The solution was covered with aluminum foil at all times to avoid light exposure due to its sensitivity to light. After 3 mL DNS solution was added to the samples and standards as described by Ghose,
48
both samples and standards were boiled for 10 min before being submerged in an ice bath for 1 min. The standards used were different glucose concentrations made with 0.05 M sodium citrate buffer (0, 0.2, 0.4, 1, and 2 mg/mL). The amount of reducing sugar released (mg) was determined with a pre-established calibration of the glucose concentrations and the total cellulase activity was calculated using the following equation:
Carboxymethyl cellulase (CMCase) (endoglucanase activity) and cellobiase (β-glucosidase activity) were also determined using the methodology described by Ghose. 48 The Amplex® Red Glucose/Glucose Oxidase Assay Kit (Molecular Probes Inc., Eugene, OR) was used to quantitate β-glucosidase. Cellobiohydrolase (exoglucanase activity) was determined using the methodology described by Wood and Bhat, 49 using Avicel. Enzyme activities were measured in units (U = conversion of 1 micromole of glucose from substrate per minute). Protein concentrations were quantitated using the PierceTM BCA Protein Assay Kit (Waltham, MA) to calculate the enzyme activity per gram of lignocellulosic biomass.
Crude Protein Analysis
The wild-type and best shuffled strain of A. pullulans were analyzed for their crude protein levels. Two mL of 4-day culture of both strains were used to inoculate the corn bran medium. Time points 0 h and 120 h (5 days) were sampled by collecting the entire contents into a 300-mL centrifuge bottle and centrifuging at 5,000 rpm for 10 min. The supernatant was decanted and the solids were dried in an 80°C static incubator for 3 days. The dried samples were then homogenized in a Krups F20342 grinder (Millville, NJ) before their crude protein levels were analyzed using a LECO model FP528 (St. Joseph, MI). Samples weights of ∼0.25 g were placed into foil packages and were then combusted in the LECO to measure the total nitrogen gas content in the sample. Protein percentage was then calculated from the nitrogen content of the sample using a conversion factor of 6.25. Samples were simultaneously assessed for dry weight by drying ∼0.25 g of sample at 80°C for 24 h.
Statistical Analysis
Cellulase activity and crude protein trials were completed in triplicates. Statistical significance and effect size evaluation were used to provide complementary information of practical significance. Using practical significance aids in the evaluation of whether the results will be applicable in industrial settings. The significance test indicates the likelihood that the observed effect is due to chance, while the effect size indicates the magnitude of the observed effects between variables.
50
To determine significant differences between the wild-type and mutant/shuffled strains, a two-tailed t-test was performed using t-Test: Two-Sample Assuming Unequal Variances (α = 0.05) in Microsoft® Excel Analysis ToolPak. Effect size value was determined using Hedges' g, rather than Cohen's d, which eliminates bias by putting weight on the pooled standard deviation based on its small sample size (n = 3).
51
Hedges, however, still uses Cohen effect sizes classification which are small (0.2), medium (0.5), and large (
To determine significant differences among the developed strains, one-way analysis of variance (ANOVA) was performed using the R statistical power analysis program and packages reshape2 and agricolae. 53 -55 Duncan's new multiple range tests (MRT) were used for post hoc tests of significant difference between means. 56
Results and Discussion
Mutagenesis Screening
The UV treatment in this study involved screening 198 A. pullulans mutants and the MMS treatment involved screening 84 mutants before producing one mutant strain with the desired phenotype of increased cellulase activity. The UV and MMS mutants were then tested for total cellulase activity (FPU) after being grown on the soybean hull medium in batch trials (t = 5 d, Fig 2). The MMS mutant yielded statistically greater total cellulase activity than the wild-type strain with effect size value g = 4.06, which showed substantial practical significance. The UV mutant produced statistically similar total cellulase activity compared to the wild-type strain, but did have a significant effect (g = 1.95).
Genome Shuffling Screening
After subjecting the two mutant strains to genome shuffling, 120 shuffled strains were screened, and from this screen nine shuffled strains were selected based on the large clear zones in the CMC plate assay (zones at least 2 mm larger than the average clear zones from the wild-type strain of ∼6 mm, n = 21). After growing the nine shuffled strains on the soybean hull medium and evaluating total cellulase activity, only two strains showed an increase in activity compared to the wild-type strain. The shuffled strains, labeled GS (genome shuffled), were named for the number designated to them during screening, resulting in GS23 and GS29.
Mutagenesis and Genome Shuffling Analysis
Figure 2 shows the total cellulase production (FPU) of UV/MMS mutant strains, shuffled strains, and the wild-type strain during batch trials (t = 5 d). The MMS mutant yielded significantly more total cellulase activity (FPU) than all other strains evaluated and had an effect size value of g = 4.06, which was a substantial practical significance compared to the wild-type strain. GS23 yielded statistically similar total cellulase activity (FPU) as the UV mutant strain and GS29 but had a significant effect (g = 2.53) compared to the wild-type strain. The UV mutant and GS29 had an effect size value of 1.95 and 0.91 compared to the wild-type strain, respectively. The three strains' cellulase production, based on effect size values, resulted in GS23 having the largest difference in FPU, followed by the UV mutant and GS29 when compared to the wild-type strain. The different values of the effect sizes observed in the three strains suggest they are not similar to each other and GS23 had high practical significance while the UV mutant and GS29 had modest practical significance compared to the wild-type strain.

Total cellulase activity (FPU) produced in batch trials (t = 5 d). (g = gram of LC biomass dry weight, WT = wild-type; UV = ultraviolet mutant; MMS = methyl methanesulfonate mutant; GS23 and GS29 = genome shuffled strains). Each data point represents the average of triplicate replications and the error bars represent the standard deviation. Different lower case letters indicate a significant difference between strains determined by Duncan's new multiple range test.
Genome Shuffling Daily Analysis
The most promising shuffled strains developed in this study (GS23 and GS29) and wild-type strain's total cellulase activity (FPU) were evaluated daily during batch trials (t = 7 d) while the UV and MMS mutant strains were only evaluated on day five of batch trials. Figure 3 shows that the total cellulase activity for GS23 was larger than the wild-type strain from day one through day five. GS23 yielded 2.4, 2.5, 3.76, 3.94, and 3.88 FPU for days one through five, respectively, and the wild-type strain yielded 0.4, 1.12, 2.09, 2.04, and 2.2 FPU on days one through five, respectively. Table 1 (data obtained from same trials as Fig. 3) shows that FPU activity for GS23 was statistically higher than the wild-type strain and had large effect size values through day five of the fermentation. The increased significance and large effect size values of GS23 compared to the wild-type strain strongly suggest substantial practical significance during the first five days of fermentation. The largest effect size value observed for GS23 was g = 5.88 on day one. This was a 6-fold increase (2.4 FPU) compared to the wild-type strain (0.4 FPU) on day one. GS29, on the other hand, produced significantly higher total cellulase activity than the wild-type strain on days one and two, but contained large effect size values on days one through five (g = 3.03 – 0.91), except day three which was small (g = 0.29). GS29 had high practical significance on days one and two, but only a modest practical significance during days four and five (p > 0.05, g = 1.02 and 0.91, respectively).
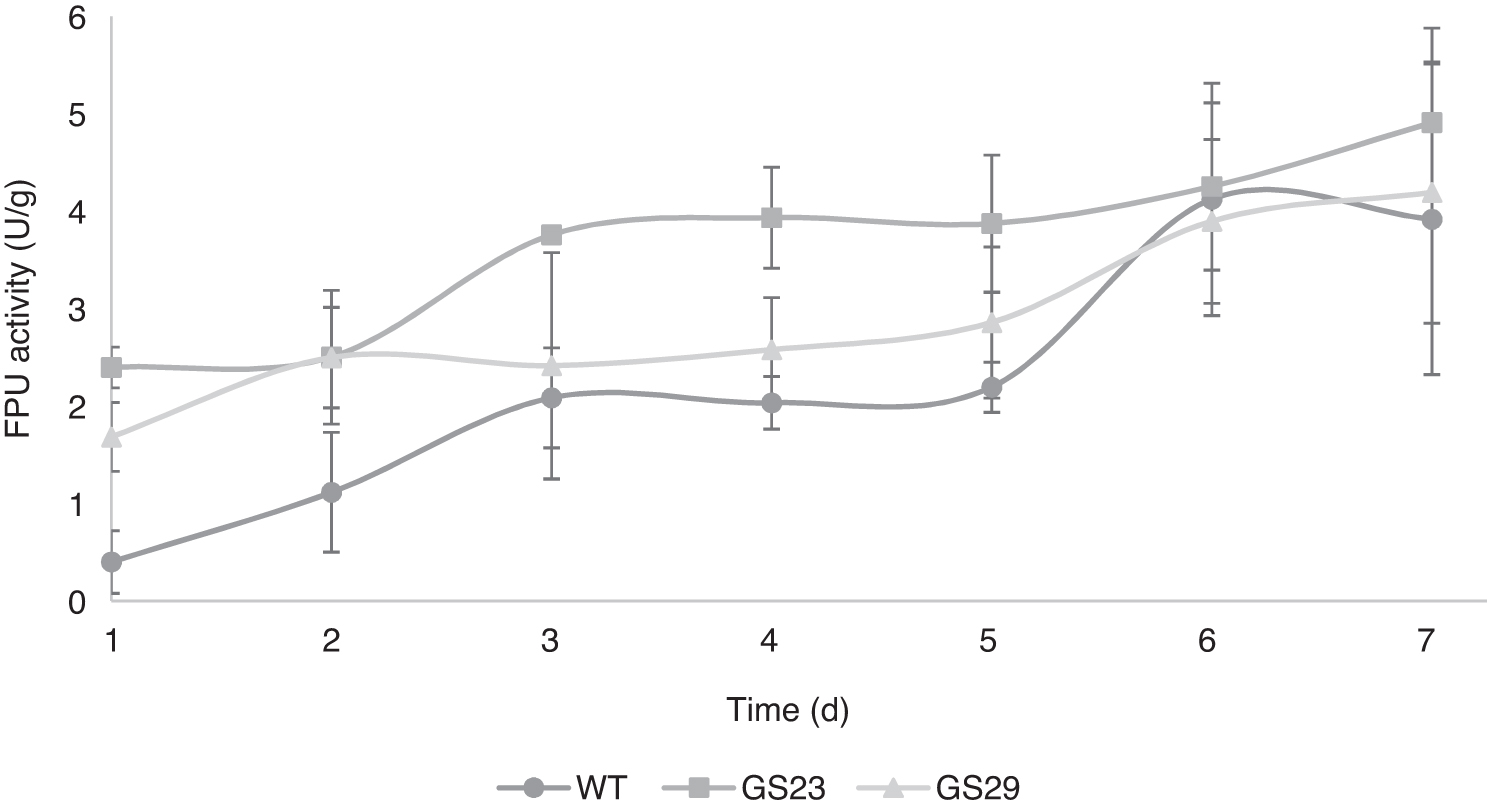
Total cellulase activity (FPU) evaluated daily during batch trials (t = 7 d). (g = gram of LC biomass dry weight, WT = wild-type; GS23 and GS29 = genome shuffled strains). Each data point represents the average of triplicate replications and the error bars represent the standard deviation.
Total Daily Cellulase Activity (FPU) of Two Fungal Strains Developed Via Genomic Shuffling (GS23 and GS29) Compared to the Wild-Type Strain
FPU evaluated by significant difference and effect size values. The p-values were found using a two-tailed t-test and the effect sizes were found by calculating Hodges' g. Asterisks (*) denote which days the shuffled strains experienced high practical significance (p < 0.05 and g ≥ 0.8) compared to the wild-type strain.
Enzyme Assay Analysis
GS23 was determined to be the most promising shuffled strain developed in this study, based on the substantial practical significance of FPU activity compared to the wild-type strain. Once this was established, assays were used to determine endoglucanase (CMCase) activity, exoglucanase (cellobiohydrolase) activity, and β-glucosidase (cellobiase) activity compared to the wild-type strain. Table 2 shows GS23 yielded significantly more exoglucanase than the wild-type strain with an effect size of g = 3.74, showing high practical significance. Endoglucanase activity was statistically similar for GS23 and the wild-type strain, and had a small effect size (g = 0.1), suggesting low practical significance. β-glucosidase activity was also statistically similar for GS23 and the wild-type strain, but did have a medium effect size (g = 0.69), suggesting a moderate practical significance.
Cellulase Activities of Wild-Type (WT) and Shuffled Strain (GS23) of A. pullulans Produced in Batch Trials (t = 5 d)
Expressed in units per gram of LC biomass dry weight. The data represents triplicate replications given as the mean ± standard deviation. Asterisks (*) denote which activity experienced high practical significance (p < .05 and g ≥ 0.8) compared to the wild-type strain.
Crude Protein Analysis
The crude protein content was also determined from the wild-type strain and GS23 strain to determine if the genome shuffled strain created more total protein versus the wild-type. Protein levels increased from 131.08 mg/g (mg of crude protein per gram of lignocellulosic biomass dry weight) in corn bran to 141.25 and 146.93 mg/g after submerged state microbial conversion of the wild-type strain and GS23 strain, respectively. This represents a relative improvement of ∼7.76 % for the wild-type strain and ∼12 % for the GS23 strain. Table 3 shows GS23 produced significantly more crude protein along with its rate of change then the wild-type strain with an effect size of g = 10.13, showing high practical significance. The crude protein content and crude protein rate of change of GS23 verses the wild-type were a 1.04-fold increase and 1.49-fold increase, respectively.
Crude Protein Comparison of Wild-Type (WT) and Shuffled Strain (GS23) of A. pullulans Produced in Batch Trials (t = 5 d)
Expressed in mg of crude protein per gram of LC biomass dry weight; 2 Expressed in mg of crude protein per gram of LC biomass dry weight per day of fermentation. The data represents triplicate replications given as the mean ± standard deviation. Asterisks (*) denote which experienced high practical significance (p < 0.05 and g ≥ 0.8) compared to the wild-type strain.
Discussion
The objective of this study was to develop a strain of A. pullulans with increased cellulase production via genome shuffling. The relatively short time period needed to develop the UV/MMS mutants and shuffled strains makes genome shuffling an ideal technique for industrial applications due to its ability to accelerate the creation of a final optimized product, while adhering to strict time constrains usually experienced in industry. 27 Due to the structural complexity of cellulase composition, biochemical characteristics, and enzymatic effects, cellulase enzyme activities can create inconsistency. 57 These factors lead to the increased probability of random error being involved in experimentation due to the complexity of cellulase, which creates difficulty in accurately quantifying the results. Triplicate trials of each strain were analyzed on the same day and larger trial numbers were not conducted because the chance of detecting a significant difference was not guaranteed due to the inconsistency in enzymatic activities. Also, by increasing the sample size to a certain undefined point, significant differences can be found even when the difference is very small because it is increasing the chance of finding a difference. 58 Thus, effect size was determined in this study to give weight based on how large of a difference occurred between the group means.
For the evaluation of the daily total cellulase activity of the shuffled strains and wild-type strains during batch trials (t = 7 d), it was determined that GS23 had substantial practical significance throughout day five of the batch trial compared to the wild-type. This is ideal as fermentation trials with A. pullulans are commonly within this timeframe. 12 GS23 (2.4 FPU) also had a 6-fold increase in FPU activity compared to the wild-type strain (0.4 FPU) on day one of the trials. The ability of GS23 to release more glucose early in the fermentation via cellulolytic degradation allows the strain to utilize these extra sugars to increase the protein content, which could then be used to enhance animal feed. GS23 was cultured for three generations were the glucose released was measured by the filter paper method (total cellulase activity) on day five. All generations showed similar glucose levels, suggesting GS23 is genetically stable.
The most promising shuffled strain developed in this study (GS23) was further analyzed to determine endoglucanase, exoglucanase, and β-glucosidase activity. An accurate quantification of each enzyme produced is important because it shows which specific enzymes the strain uses to breakdown cellulose. GS23 exhibited low practical significance with endoglucanase activity (p = 0.93, g = 0.1), but had a substantial practical significance with exoglucanase (p = 0.03, g = 3.74) when compared to the wild-type strain and was determined to be effective at releasing cellobiose due to its large exoglucanase activity. β-glucosidase production in GS23 yielded a moderate practical significance compared to the wild-type due to its medium effect size (g = 0.69). These findings allowed us to conclude that GS23 is a preferred cellulolytic strain due to its increased ability to release cellobiose due to exoglucanase activity (6.95-fold increase compared to the wild-type strain). Also, the release of cellobiose activates β-glucosidase activity to release more glucose into the growth medium than the wild-type strain. GS23 had a 1.52-fold activity increase in β-glucosidase activity compared to the wild-type strain.
The crude protein was assessed to determine if GS23 could produce higher total protein levels then the wild-type. The corn bran fermented with wild-type strain and GS23 strain were analyzed on day five for increased protein levels since this is the time frame of growing this organism for increased protein production 12 and was the last day it showed significant difference. Visual differences could be seen between the wild-type and GS23 strains when it was grown on submerged-state corn bran. After day five, the wild-type strain turned the media a murky, dark brown color while the GS23 strain was a light tan. After both strains were centrifuged, the collection of cells could be seen on top of the corn bran. The wild-type strain pigmentation was dark brown and grey while the GS23 strain pigmentation was a light pink. GS23 had high practical significance for the crude protein content and crude protein rate of change than the wild type strain (p = 0.003, g = 10.13). High levels of protein production with GS23 could be attributed to the strains' ability to degrade more cellulose, adding more glucose into the medium that could be fermented into a higher concentration of protein.
The A. pullulans strain Y-2311-1 has only been sequenced for some genes, but not cellulase. 59 Therefore, being able to identify the genetic changes in the shuffled strain GS23 would be of great interest. But identifying the specific gene or genes responsible for the phenotypic changes in GS23 through sequencing is very challenging and does not always lead to the decisive conclusion. A. pullulans has been characterized to have extreme variability and genetic instability because of their high heterokaryotic condition. The mitotic gene exchange process has been known to happen in heterokaryotic fungi and possibly contribute to the morphological and physiological variation found in the fungi. 60 Slepecky and Starmer 16 found that when the same strain of A. pullulans was grown in different environments, the changes in gene expression could lead to changes in phenotype. Also, due to the fact that genome shuffling is the result of random mutations, the phenotypic changes could be epigenetic or multigenic. Thus by sequencing, it would be inconclusive in determining the specific gene or genes responsible for the phenotypic changes and is not recommended for the shuffled strain created in this study.
Conclusion
This study serves as proof-of-concept that genome shuffling can be used to develop A. pullulans strains with increased lignocellulose conversion. One of the shuffled strains developed in this study (GS23) may have potential as an industrial microorganism in a cellulolytic process. Future studies include evaluating GS23's performance on various feedstocks and animal feeding trials to evaluate the animal's performance from feed produced by GS23. The total cellulase amounts produced by GS23 had substantial practical significance compared to the wild-type during batch trials. Specifically, exoglucanase and β-glucosidase activities had increased practical significance. The improved cellulolytic strain showed its potential to increase protein levels in lignocellulosic biomass with its substantial practical significance of crude protein compared to the wild-type strain. This capability could lead to the development of improved protein-rich animal feed by decreasing the amount of commercial enzymes used in the fermentation process.
Footnotes
Acknowledgments
The authors acknowledge Dr. Lu-Kwang Ju for providing his modified procedure for measuring total cellulase activity by the filter paper method.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funding support was provided by the South Dakota Soybean Research and Promotion Council.