
Abstract
The aim of this work was to maximize the production and the partial characterization of carboxymethylcellulase (CMCase) and filter paper enzyme activity (FPase) using a new isolate by solid state fermentation with agro-industrial substrates and potential application of hydrolytic cellulosic substrates. Four isolates were initially evaluated, and the isolate I-130, identified as Penicillium sp., was selected for FPase and CMCase production. The maximized conditions for the enzyme bioproduction were inoculation of 5 to 7 log spores/g, in soybean hull, at 70% moisture for 7 d at 30°C. The best solvent for extracting the enzymes was citrate buffer (0.05 M, pH 4.5) with a time of 30 min, temperature of 50°C, and agitation of 195.5 and 125 rpm for CMCase and FPase, respectively. The optimum pH and temperature for FPase and CMCase activity were 4.9 and 50°C. Both enzymes were more stable at pH 4.9. FPase and CMCase showed thermal stability only up to 40°C, with rapid deactivation at higher temperatures; the FPase being more stable at this temperature range. The deactivation energy was 89.3 and 78.0 KJ/mol/ for FPase and CMCase, respectively. The highest hydrolytic activity occurred at 350 rpm for soybean hulls (22%) and corn stover and cobs (4.4%).
Introduction
Cellulases production using alternative substrates has been well-studied, because it is a promising way to reduce enzyme production costs, one of the major bottlenecks for its application. The composition of the culture medium significantly affects product concentration, yield, and volumetric productivities, which is of central importance for economically viable production. The use of agro-industrial residues is an interesting strategy for reducing costs associated with the culture medium formulation of cellulase production. 1
In this context, agro-industrial wastes appear as ideal substrates for microbial fermentation, because of their rich organic components content, low cost, and wide availability. 2,3 Agricultural residues are byproducts of certain crops, such as rice, wheat, oil seeds, and grains, and are among the most abundant agricultural byproducts in the world. 4 Furthermore, the use of agricultural byproducts presents a better energy balance and a lower environmental impact than those of pure substrates. 5
Solid state fermentation (SSF) is a suitable strategy for producing low cost enzymatic products. 2 The SSF of lignocellulosic biomass presents important advantages over the conventional submerged fermentation, such as reduced energy requirements, high productivity, and less inhibitory effects for enzymatic production. 6 Also, SSF allows the use of organic wastes for biotransformation into bioproducts, enabling the production of a concentrated product because of the low amounts of water used in the process. 3,7
Cellulolytic enzymes play an important role in the bioconversion of cellulosic biomass; nevertheless, the production cost of these enzymes has been an obstacle in industrial production and bioprocesses. Hence, there is a focus on identifying microorganisms that secrete these enzymes with higher efficiency and cheaper carbon sources. 8
Recently, many microorganisms such as bacteria, actinomycetes, and fungi, have been recognized for their ability to hydrolyze lignocellulosic materials. The most commonly used microbes to produce hydrolytic enzymes are Pseudomonas, Clostridium, Bacillus, Aspergillus, Trichoderma, and Penicillium. 9,10
The objective of this study was to select a new strain for the bioproduction of FPase and CMCase by solid state fermentation using agro-industrial substrates, and then maximize the production and the extraction process, partially characterize the obtained crude enzymatic extracts, and evaluate the potential application on cellulosic substrates' hydrolysis.
Materials and Methods
Microorganism Selection
Four microorganisms (I-70, I-26, I-130, and I-34), previously isolated by Griebeler et al. 11 were used, because they had potential for production of carboxymethylcellulase (CMCase) and filter paper enzyme activity (FPase) in qualitative screening.
Enzyme Bioproduction
The soybean hulls (Olfar Company, Frinape, Brazil) substrate was used at all stages, without prior treatment and characterized as crude fiber, crude protein, and fixed mineral residue by near infrared reflectance spectroscopy (NIRS).
The solid-state fermentations to the isolates' selection (Table 1) 7 -25 were carried out with 10 g of dry soybean hulls and subsequently, with humidity adjusted to 70% and injection of moist air into the germination chamber (MOD 347 CDG), inoculum of 1x10 7 spores/g at 30°C for 14 d of fermentation. The extraction of the enzymes was done using a ratio of 1:15 (w/v) fermented substrate and citrate buffer (0.1M pH 4.8), with agitation at 100 rpm and 50°C for 30 min. 12 The spore suspension was prepared by incubating the cultures on Potato Agar Dextrose (PDA) plates at 30°C for about 5–6 days, until sufficient sporulation was observed. The spores were harvested using 0.1% tween 80 solution (v/v).
Selection of Microorganisms for the Production of FPase and CMCase Enzymes
Means followed by the same lowercase letters in the columns for each enzyme and upper case in the lines for each fungus, do not differ significantly from each other by the Tukey test with 95% confidence.
After the selection of the microorganism, the influence of the temperature (20, 25, 30 and 35°C) on the bioproduction of CMCase and FPase was evaluated. The bioproduction was carried out with soybean hulls (humidity of 70%), spore concentration 10 7 , and 7 d of fermentation. The extraction conditions were 50°C, 100 rpm, for 30 min using 0.1M citrate buffer at pH 4.8, as described by Dhillon et al. 12
To evaluate the effect of the spore concentration and the moisture content on the production of FPase and CMCase for 7 and 14 d of fermentation, a matrix of the central composite rotatable design (CCRD) 2 2 (Table 2) was performed, varying the spore concentration (4.6, 5.0, 6.0, 7.0, and 7.4 log spores/g dry substrate) and moisture (34, 40, 55, 70, and 76%), keeping the fixed conditions of 10 g of soybean hull and 30°C for bioproduction. The extraction conditions were 50°C and 100 rpm for 30 min using 0.1M citrate buffer at pH 4.8, as described by Dhillon et al. 12
Matrix of the Central Composite Rotatable Design (CCRD) 2 2 (Real and Coded Values) for FPase and CMCase Activity at Different Concentrations of Spores and Moisture Content for 7 and 14 D of Culture
X1: Spore concentration (Log); X2: Moisture (%); Fixed independent variables: 10 g of soybean hull, 30°C.
Enzyme Extraction
The solvents, sodium citrate buffer (0.05 M pH 4.5), sodium acetate buffer (0.2 M pH 5.5), tween 80 (0.1%), NaCl solution (1.0%), and distilled water were evaluated for FPase and CMCase extractions, performed with 50 mL solvent and 3.33 g fermented substrate, agitated at 100 rpm, at 50°C for 30 min.
The influence of temperature (35.9, 40, 50, 60, and 64.1°C) and agitation (54.5, 75, 125, 175, and 195.5 rpm) was evaluated through a matrix of the CCRD 2 2 in 30 min of extraction (Table 3).
Matrix of the Central Composite Rotatable Design (CCRD) 2 2 (Real and Coded Values) for FPase and CMCase Activity in Relation to Temperature and Shaking in Extraction
X1 = Temperature (°C); X2 = Shaking (rpm). Fixed independent variables: 30 min of extraction.
Partial Characterization of Enzymes
The pH (3.9 to 6.3) and temperature (36 to 64°C) of determination of FPase and CMCase activity were evaluated by matrix of the CCRD 2 2 (Table 4). The solutions that were employed for the adjustment of the pH buffer were 0.05 M Tris-HCl and 0.01 M NaOH.
Matrix of the Central Composite Rotatable Design (CCRD) 2 2 (Real and Coded Values) for FPase and CMCase Activity in Relation to Optimal pH and Temperature
X1: pH and X2: temperature.
The kinetics of the enzyme activity stability at different pH (4.0, 4.9, 6.0, and 6.5) were performed at 25°C for up to 160 min.
Thermal Stability
To determine the thermal stability of the enzyme extract, these were subjected to different temperatures (-20, 4, 25, 40, 50, 60, and 70°C) at pH 4.9. The stability at different pH values (4.0, 4.9, 6.0, and 6.5) was evaluated at 25°C. The samples were collected periodically to evaluate the enzymatic activities. The thermal and pH stability was expressed in terms of the residual enzymatic activity (%).
The half-life time (t1/2), thermal denaturation rate constant (Kd), D-value, and Z-value were determined at temperatures of 25, 40, 50, 60, and 70°C (Table 5).
Experimental Values of kd, t1/ 2, D Value, Ed and Z Value for CMCase and FPase in Relation to Temperature
The data obtained experimentally for the thermal stability (25, 40, 50, 60, and 70°C) of the crude enzyme extract were plotted as ln (A/A0) as a function of time (min), where A and A0 are the residual and initial activities (U/g), respectively. Following this procedure, the thermal deactivation constant (Kd) was obtained at each temperature. A plot of ln (Kd), as a function of 1/T (K), was used to calculate the deactivation energy (Ed) for the deactivation reaction. 13
Cellulosic Substrates' Hydrolysis Evaluation
Enzymatic saccharification for each non-previously-treated substrate was carried out as described by Liu et al. 14 The assays were done in erlenmeyer flask with the addition of 2 g cellulosic residue (soybean hulls, SH; corn stover and cobs, CSC), and autoclaved for 15 min at 121°C. Subsequently, 100 mL of a reaction mixture containing 95 mL sodium citrate buffer of 0.05 M, pH 5.0, and 5 mL crude enzymatic extract was added.
The kinetic behavior of the lignocellulosic substrates' hydrolysis process (for SH and CSC), using crude enzymatic extract, was monitored by removing the samples from the reaction mixture at various time intervals. The hydrolysis reactions were performed using an orbital shaker at 37°C at three different agitations (200, 300, and 350 rpm) for 48 h. The total reducing sugars (TRS) released in the hydrolysis process was estimated using DNS (dinitrosalicylic acid) methodology. 15
The saccharification percentage was calculated according to Equation 1.
16
where, RS = released sugar; 0.9 = correction factor; P = polysaccharides in lignocellulosic substrate (0.0505 for SHs 17 and 0.584 for CSCs 18 ).
CMCase AND FPase ACTIVITY
The carboxymethylcellulase activity was measured according to Ghose, using carboxy methyl cellulose (CMC) as the substrate and 3,5-dinitrosalisylic acid (DNS) as the coupling reagent. 19 The reaction mixture contained 1 mL of 0.05 M citrate buffer of pH 4.8, 0.5 mL of 2% CMC, and 0.5 mL of the extract crude enzyme. After incubation at 50°C for 30 min, 1 mL of DNS reagent was added. The mixture was boiled for 5 min and cooled immediately in ice. The absorbance was measured at 540 nm against the reagent blank. The released glucose was determined by comparing the absorbance with the standard glucose solution. One unit of CMCase activity was defined as the amount of enzyme that liberates 1 μmol of reducing sugars, equivalent to glucose per minute, under the assay conditions described.
The filter paper enzyme activity (FPase) was measured according to Ghose, using Whatman N°1 filter paper as the substrate. 19 To correct the reducing sugar present in the enzyme preparation, a blank without filter paper was also run. In this assay, 0.5 g Whatman filter papers (1.0 × 6.0 cm) is the reaction product, and its absorbance is measured by UV–vis spectroscopy. Then, 0.5 mL of extract crude enzyme and 1 mL citrate buffer (0.2 mM citrate buffer, pH 4.9) were incubated for 1 h at 50°C. After this time, 0.5 mL of the mixture was poured into another tube, and 0.5 mL DNS reagent was added to the mixture. The tubes were kept in a boiling water bath for 5 min and cooled immediately in ice. Finally, the absorbance of the solution was read at 540 nm. One filter paper nit (FPU) was defined as the amount of the enzyme that released 1 μmol of glucose per minute from the original substrate at the experimental conditions.
Statistical Analysis
The results were treated using the software Statistica, version 5.0 (Statsoft Inc, Tulsa, OK), considering a level of 95% of confidence (p < .05).
Results and Discussion
Microorganism Selection
The agro-industrial substrate used in this study was soybean hull (SH) for its abundance and low cost. It presented moisture content of 10.06%, crude fiber content of 41.14%, crude protein content of 9.85%, and fixed mineral residue of 4.94%. The high fiber and crude protein content justifies its use as a substrate for bioproduction.
Four isolates (I-70, I-26, I-130, and I-34) were preselected based on the halo diameter, when cultured on the carboxymethyl cellulose agar, for selection of the microorganism with the potential for production of CMCase and FPase on SH substrate. 11 For FPase enzyme, it was verified that the highest enzymatic activity (1.03 U/g) was at 7 d of fermentation with I-130—not statistically differing from other fungi, except for I-26 isolate (0.74 U/g), which was statistically lower. For the CMCase, the isolates I-130 (8.77 U/g), I-70 (7.19 U/g), and I-26 (8.16 U/g) showed the highest enzymatic activities in 10 d, not differing statistically from each other, but differing for the I-34 isolate (2.48 U/g) (Table 1).
The isolate I-130 (Penicillium sp.) was selected for maximizing bioproduction and characterization of the crude enzymatic extracts because it presented the highest activities of FPase and CMCase (Table 1).
Enzymes Bioproduction
The optimum temperature for the bioproduction of CMCase and FPase was 30°C, with activities of 16.21 and 1.42 U/g, respectively. Both low (20°C) and high (35°C) temperatures caused a significant reduction in the production of CMCase (10.3 and 4.3 U/g) and FPase (1.28 and 0.63 U/g), respectively. According to Kim et al., 20 the optimum temperature range to produce enzymes from Penicillium sp. oscillates between 25 and 35°C, with the optimum temperature of 30°C to produce FPase and CMCase, which was confirmed in this work.
The influence of initial spore concentration and the substrate moisture (SH) on FPase and CMCase production after 7 and 14 d of solid state fermentation by Penicillium sp. (I-130 isolate) was evaluated by a central composite rotatable design (CCRD) 2 2 (Table 2).
The maximum activities for FPase (1.19 U/g) and CMCase (16.37 U/g) were obtained with 5 and 7 log spores/g inoculation respectively, both with 70% moisture for 7 d of culture. These results are promising when compared to Silva et al., 8 who studied the production of FPase and CMCase, with hydrogen peroxide-alkaline pre-treated carnauba straw residue, obtaining 0.9 and 13 U/g, respectively.
Equations 2 and 3 present the second order coded models that describe the activities of FPase with 7 and 14 d of cultivation, within the studied ranges, respectively. The models were validated using variance analysis. The obtained correlation coefficients were 0.97 and 0.98, and the calculated F values were 5.25 and 6.97 times greater than the tabulated F, respectively. The non-significant parameters were added to the lack of adequacy for variance analysis (ANOVA) test. The model presented in Equation 4 describes the activity of CMCase with 14 d of cultivation, obtaining correlation coefficient of 0.97 and an Fcalculated 4.62 times greater than the value of tabulated F.
where, FPase and CMCase activity is in U/g; X1 = Spore concentration (Log); X2 = Moisture (%).
From the validation of the models, it was possible to construct the contour curves (Fig. 1), where it can be observed that the maximum FPase production, at 7 and 14 d of cultivation, occurred with a concentration of 6 log spores and moisture of 70%.
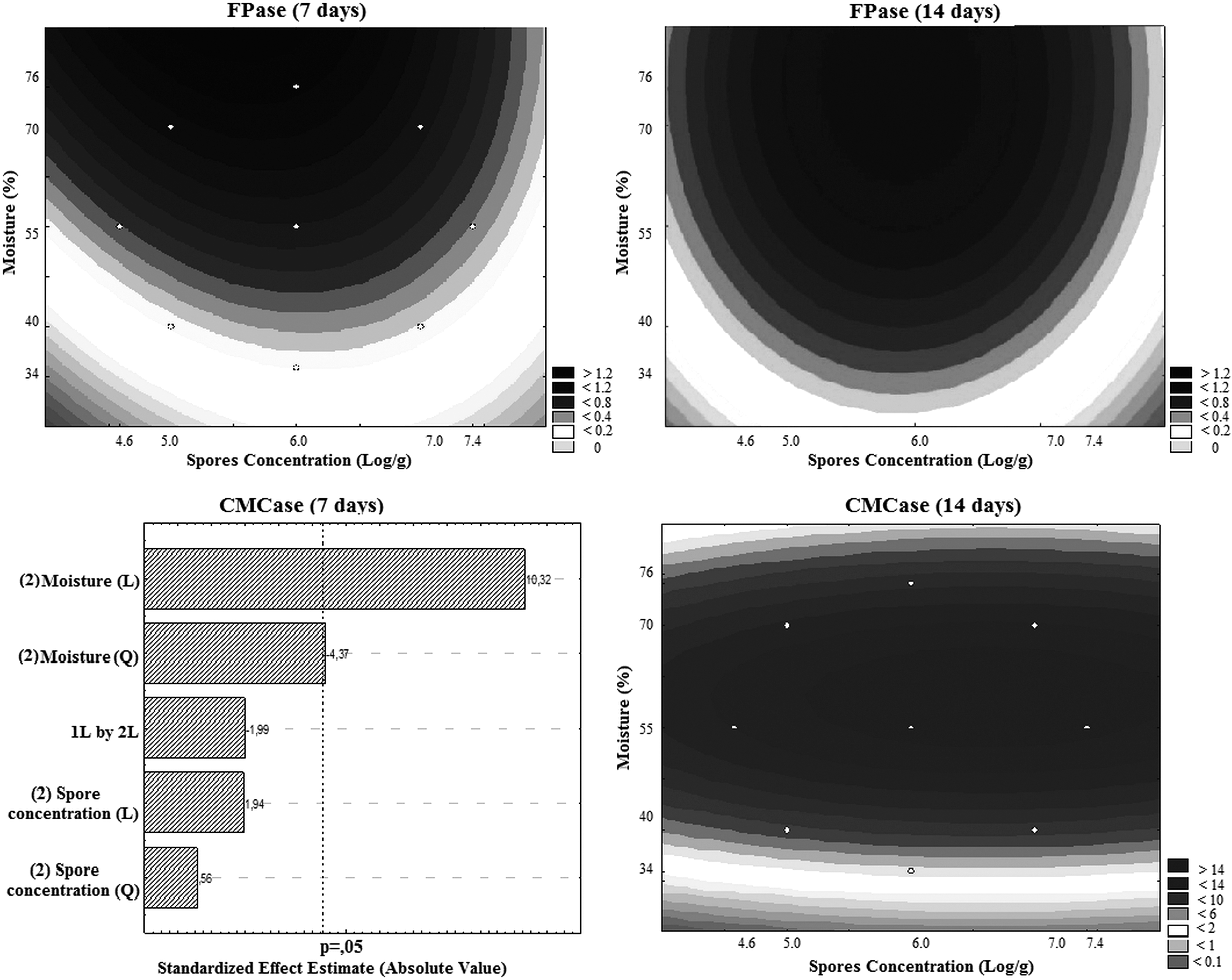
Contour curves for bioproduction of FPase (7 and 14 d) and CMCase (14 d) and Pareto graphic of CMCase (7 d), in relation to spore concentration and moisture in bioproduction.
The results of the effects of moisture and spore concentration on CMCase activity were treated statistically, and the variable moisture (linear and quadratic) showed a significant positive effect (p < .05), demonstrating that an increase in the moisture will probably cause an increase in CMCase activity for 7 d (Fig. 1). The maximum CMCase production in 7 and 14 d was between 55 and 70%, independent of the spore concentration and within the range evaluated (Table 2, Fig. 1 ).
In general, for both enzymes, it was observed that the variable with the greatest effect was moisture (Fig. 1), i.e., the higher the humidity, the higher the production of the enzymes for any concentration of spores, regardless of the bioproduction time. Therefore, the best strategy to produce FPase and CMCase can be defined as humidity of 70%, with any concentration of spores (within the studied range), for 7 d. Reductions of 37–43% and 44–46%, for FPase and CMCase, were observed for runs 3 and 4 (Table 2), respectively, in 14 d.
FPase AND CMCase Extraction
The solvents sodium citrate buffer (0.05 M, pH 4.5), sodium acetate buffer (0.2 M, pH 5.5), tween 80 (0.1%), NaCl Solution (1.0%), and distilled water presented extraction results of 2.7, 2.1, 1.3, 1.2, and 1.6 U/g for FPase and 14.0, 12.0, 10.9 , 13.7, and 10.6 U/g for CMCase. It was observed that the sodium citrate buffer is the best solvent for extracting FPase and CMCase, corroborating Dhillon et al. 21 The recovery of enzymes obtained by solid-state fermentation is influenced by the interacting forces between the enzymes and their substrates. The highest enzymatic recoveries were achieved with solvents that promoted the reduction of these interactive forces. 22 Because of the hydrophobic/hydrophilic nature of the fungal mycelium, the Vander Waals forces and the ionic and hydrogen bonds also determine the efficiency of the enzyme recovery. 23
The influence of temperature and agitation was evaluated through a central composite rotatable design (CCRD) 2 2 (Table 3). The maximum activity of FPase (3.28 U/g) and CMCase (26.63 U/g) was obtained at 50°C and 125 and 195.5 rpm, respectively.
Equation 5 presents the coded second order model that describes the activity of FPase as a function of temperature and agitation, within the studied ranges. The model was validated using variance analysis. The obtained correlation coefficient was 0.97 and the calculated F value was 83.69 times greater than the tabulated F. The non-significant parameters were added to the lack of adequacy for variance analysis (ANOVA).
where, FPase activity is in U/g; X1 = Temperature (°C); X2 = Agitation (rpm).
From the validation of the model it was possible to construct the contour curve (Fig. 2), where it can be observed that the maximum extraction of FPase is in the range of 40–60°C and 75–175 rpm. The results of the effects of temperature and agitation on CMCase activity were treated statistically (p > .05) and showed no significant effect (Fig. 2).
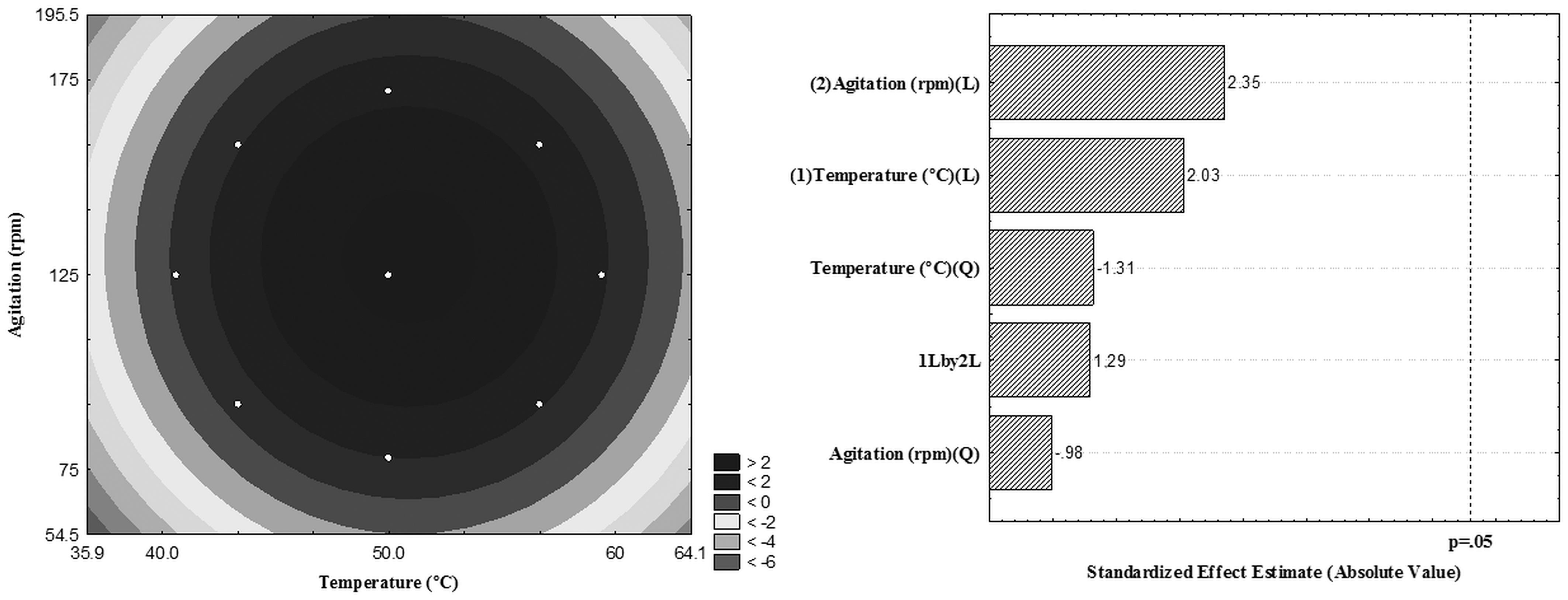
Contour curve and Pareto graphic with the estimated effect (absolute value) of the variables tested in the complete factorial design 2 2 , for the extraction of FPase and CMCase, respectively.
These results are comparable to those obtained by Astolfi, 24 who, with Trichoderma reesei NRRL 3652 cultivation in SH, obtained CMCase activity of 5.45 U/g and FPase activity of 6.71 U/g after 3 d. Florencio, using mixed substrate of sugarcane bagasse and wheat bran (50%:50%), with spore concentration of 10 7 spores/g, at 30°C in 8 days of cultivation of Trichoderma sp., obtained activity for CMCase of 5.95 U/g. 25
However, higher production results have already been reported by other authors with other substrates and microorganisms. Dhillon et al., using Aspergillus niger, obtained a CMCase activity of 48.2 U/g and FPase of 383.7 U/g after 120 h, using wheat bran substrate. 12 Pirota et al. evaluated the production of FPase and CMCase in wheat bran with several strains isolated from the fungi, and the highest production of FPase (2.25 U/g) was with the P75P1 lineage. 26 For CMCase, the highest yield was 294.33 U/g and 351.79 U/g with A. fumigatus P40M2 and A. niger, respectively.
Partial Characterization of FPase AND CMCase
Optimum pH and temperature
The optimum pH and temperature evaluation was performed using the CCRD 2 2 . Table 4 presents the experimental design matrix with the studied variable ranges and the responses in terms of FPase and CMCase activity. The highest activity was 2.92 and 22.00 U/mL (runs 9, 10, and 11) at 50°C and pH 4.9.
These results were statistically treated, and Equation 6 presents the empirical mathematical models obtained through the variance analysis, referring to the enzymatic activities of FPase, as a function of pH and temperature within the studied range.
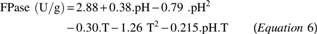
The calculated F, greater than the tabulated value (R 2 > 95%), allowed the statistical validation of the empirical mathematical model (p < .05) and the construction of the contour curve presented in Fig. 3, which confirms that for the FPase enzyme, the optimum temperature was close to 50°C and the optimum pH was 4.9. The results of pH and temperature effects on CMCase activity were treated statistically and showed no significant effect (p > .05), as shown in the Pareto graphic (Fig. 3). In general, the cellulases, produced by filamentous fungi, have optimum pH values in the acidic range (3.6–5.0) and optimal temperatures above 40°C. 24,27 -29
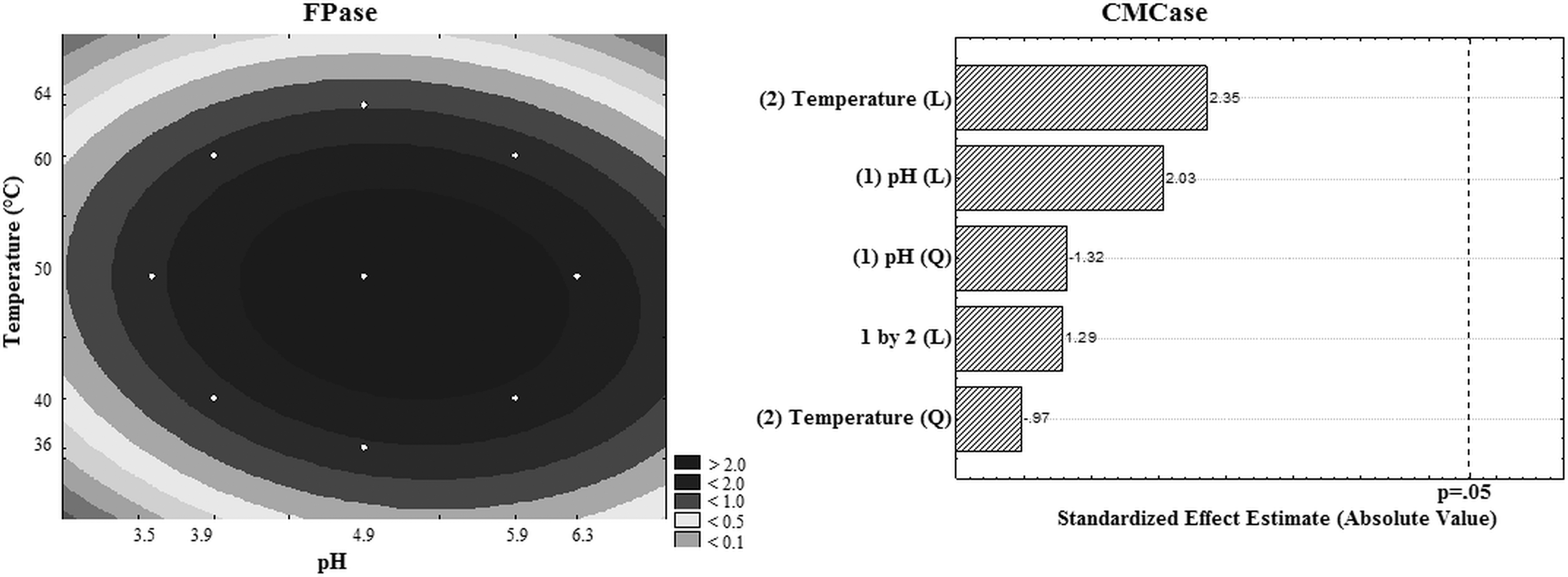
Contour curves and Pareto graphic in relation to optimal pH and temperature for FPase and CMCase activity, respectively.
Enzymatic pH stability
The stability of crude CMCase and FPase enzyme extract (4.0, 4.9, 6.0, and 6.5) was performed to a different pH level by monitoring the behavior as a time function. Figure 4 shows the results of the kinetics of enzymatic stability at different pH, where both enzymes were more stable at pH 4.9, with a reduction of 50% of the activity after 132 and 144 h of storage at 25°C for CMCase and FPase, respectively. Similar results have been reported in Bacillus subtilis strain LFS3, 30 Thermomonospora, 31 Bacillus sp. M-9, 32 B. licheniformis, 33 and Bacillus sonorensis HSC7. 34 The results of the enzymatic pH stability characterize the acidophilic nature of the enzyme, demonstrating the potential of the new isolate, Penicillium sp., for the production and use of the cellulolytic enzymes in the industrial processes, such as in the juice industry.
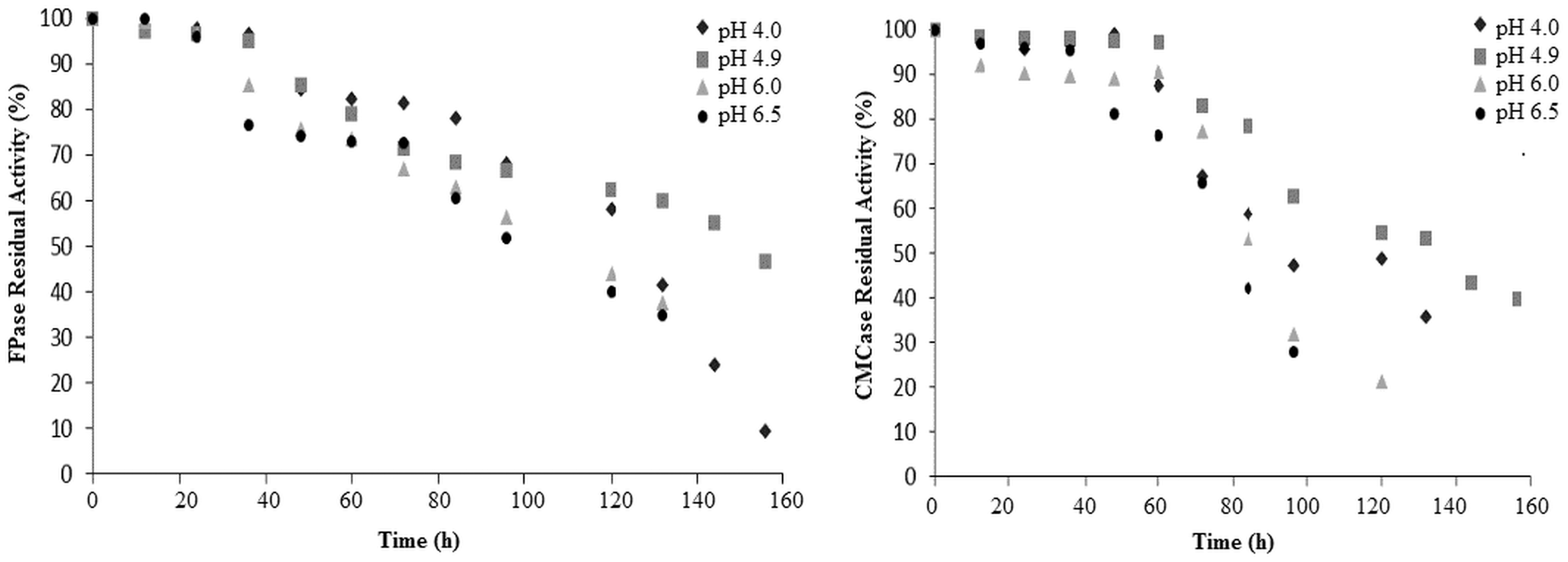
Kinetics of enzyme activity and stability for FPase and CMCase at different pH values.
Enzymatic thermal stability
The crude FPase and CMCase extracts (Fig. 5) were stored at -20, 4, 25, 40, 50, 60, and 70°C, and the enzymatic activity was measured periodically. The thermal stability evaluation of the FPase and CMCase enzymes consisted of incubating the crude enzyme extract (at pH 4.9) for up to 160 h at temperatures ranging from 25 to 70°C or -20 to 4°C, evaluated every 20 d. FPase presented higher thermal stability in relation to CMCase, both in high and low temperatures.
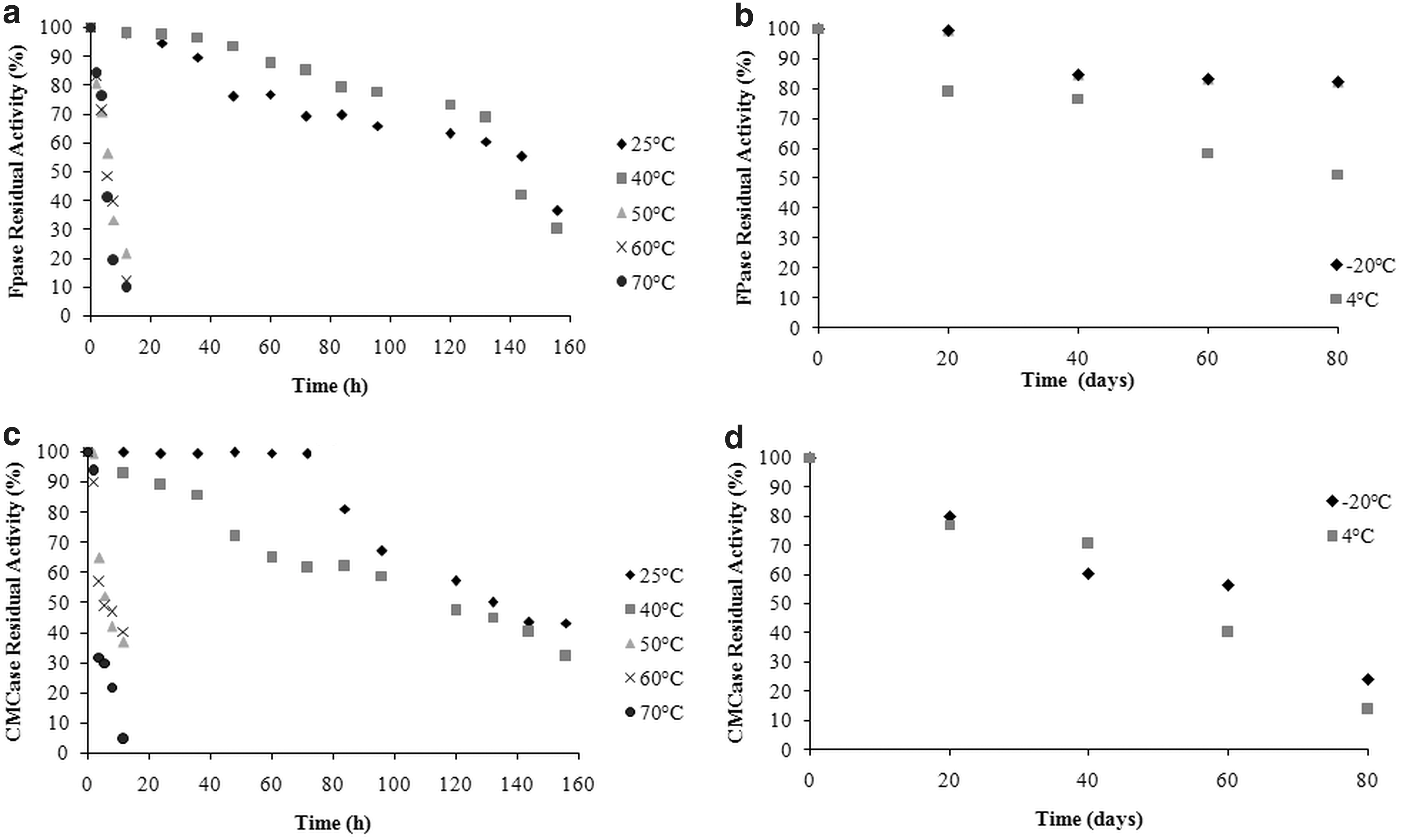
Stability of
The FPase enzyme presented higher stability when stored at temperatures of 25 and 40°C, with about a 50% reduction in activity after 140 h of storage. High enzymatic activity losses were observed at temperatures of 50, 60, and 70°C (Fig. 5a), with a reduction of about 50% of the enzymatic activity in less than 10 h. After 90 d of storage at -20°C, the FPase viability was around 74%. At 4°C, a lower residual activity was observed for FPase—around 58% after 60 d (Fig. 5b).
The CMCase showed higher stability at 25 and 40°C, with a reduction of about 50% in activity after 132 and 96 h of storage, respectively. Drastic losses in enzyme activity were observed when the crude extract was stored at temperatures of 50, 60, and 70°C, with a 50% reduction after 4–6 h (Fig. 5c).
At low temperatures, residual activity of 39 and 56% was observed at 4 and -20°C, respectively, after 60 days of storage (Fig. 5d). Similar or even lower results regarding the stability of CMCase and FPase were reported by Astolfi, 24 Falkoski et al., 29 Ladeira et al., 35 and Santos et al. 36
Kinetic parameters
The half-life (t1/2), kinetic thermal denaturation constant (Kd), D-value, deactivation energy (Ed), and Z-value for the 25, 40, 50, 60, and 70 °C temperatures of CMCase and FPase are shown in Table 5.
The increase in temperature affected the thermal deactivation process of the enzymes, evaluated with similar inactivation rates (Kd) and different temperatures, between CMCase and FPase. The FPase was shown to be more stable at thermal deactivation than CMCase, with half-life times (upper to 154.03 h) and D-values (upper to 511.68 h) at 40°C. The high values of these parameters are desirable during the industrial operations because they guarantee that the enzyme is more resistant to the deactivation caused by the effect of temperature. 37,38 However, temperatures of 50°C or higher caused a strong enzymatic deactivation for both the enzymes.
The Ed values of CMCase and FPase were 78.0 and 89.3 kJ/mol, respectively. Different authors have found variable Ed values of cellulases of enzymatic extracts from different isolates. In the study by Balsan et al., the value found for Ed was 177.6 kJ/mol, produced by the enzyme cellulase from the fungus Trichoderma reesei (NS 50013) in media of lignocellulolytic cultures. 39 Calsavara et al., using Aspergillus niger on the cellobiose substrate, obtained values for thermal deactivation energy of 80.6 Kcal/mol in free enzyme. 40
Z values of 25.0 and 22.2 indicate that a change in these temperature ranges would generate a reduction of one logarithmic cycle in the enzymatic activity of CMCase and FPase, respectively.
Cellulosic Substrates Hydrolysis
The kinetic behavior for the hydrolysis process of the SH and CSC substrates using the crude enzymatic extract is shown in Fig. 6. For SH and CSC, the highest percentage of hydrolysis (22 and 4.4%, respectively) was observed in 36 h of the reaction, at 350 rpm. For the SH, there is no statistical difference in the percentage of hydrolysis after 36 h of reaction, while for CSC, this behavior is observed after 24 h. Teixeira, evaluating the hydrolysis of SH and CSC with commercial cellulase of Aspergillus niger (Sigma-Aldrich, St. Louis, MO), obtained 47.95 and 26.05% hydrolysis, respectively. 41 Hydrolysis of SH with crude extract of the new isolate reached 48% in relation to that obtained by Teixeira with commercial enzyme—demonstrating its high potential in the hydrolysis of this substrate. 41 However, the same potential was not observed in CSC hydrolysis; only 17% of the hydrolysis was obtained with the commercial enzyme.
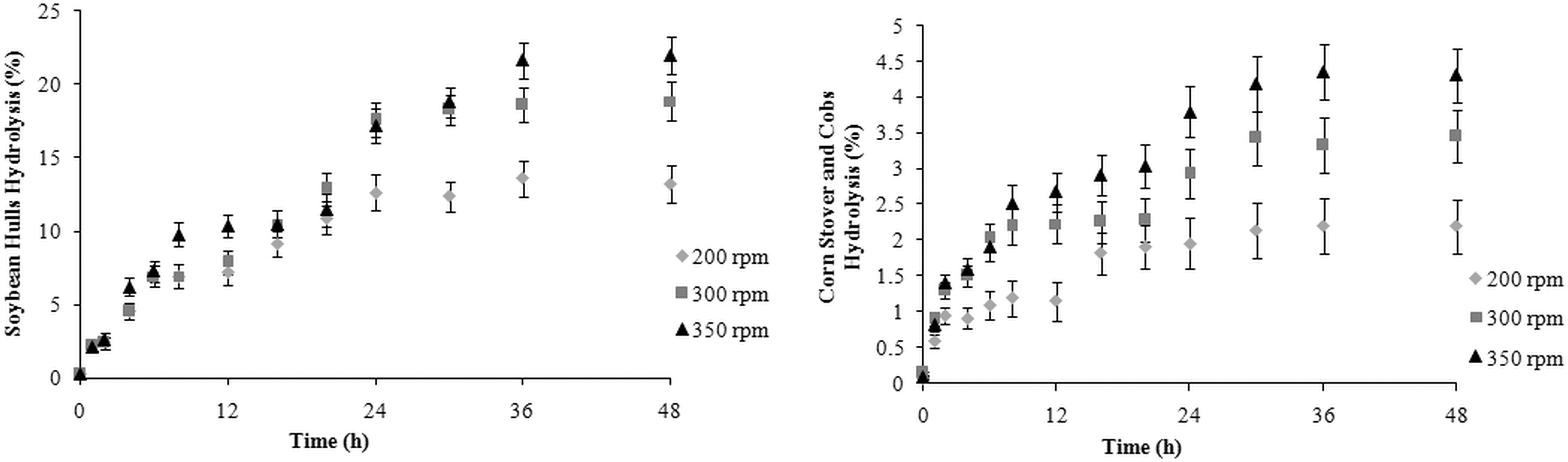
Influence of agitation on hydrolysis of soybean hulls (SH) or corn stover and cobs (CSC) with crude extract enzymatic.
According to Reis et al., different groups of enzymes acting synergistically may be more effective at breaking down complex polysaccharides. 42 However, the low hydrolysis values obtained in this work can be related to the fact that CSC presents 39.8% cellulose and 22.5% lignin, which was also reported by Oliveira. 43 Already, better hydrolysis results were obtained with the soybean hull, which can be explained by the lower lignin content (1.75%) and the maintenance of similar levels of cellulose (36.4%). 44
In this context, studies suggest the use of enzymatic cocktails for effective enzymatic degradation of the cellulosic materials, 45 and in many situations, enzymes produced from the mixture of several microorganisms.
Conclusions
The substrate moisture had the greatest influence on the bioproduction of FPase and CMCase from the new isolate. The enzyme recovery was higher with citrate buffer at 50°C and 150 rpm. The partial characterization of the enzymatic extract indicated the acidic nature and thermal stability up to 40°C. The enzymatic extract presented differentiated hydrolysis for the evaluated substrates, indicating potential application in substrates with low lignin content, such as the soybean hull.
Footnotes
Acknowledgments
The authors thank the CAPES, CNPq, and FAPERGS for financial support.
Author Disclosure Statement
No competing financial interests exist.