
Abstract
This study aims to partially compensate the lack of reliable data regarding full mass balance analysis during industrial baker's yeast fermentation and the chemical composition of the resulting vinasse. The utilization degrees of carbon and nitrogen sources (sugars, amino acids, ammonia) for biomass growth and protein synthesis are discussed. Furthermore, the less complex composition and the suitability of baker's yeast vinasse for the industrial recovery of large amounts of natural betaine and invertase are highlighted. Implementing the corresponding technology would have a strong and positive impact on process profitability. Conducted analyses included the determination of sugars and protein concentrations as well as the complete amino acid profile. Also, betaine concentrations and invertase activity were determined in the molasses mixture and the resulting vinasse. The results show that vinasse contained 18.5 g/L of betaine and an invertase activity of 21.3 U/mL. Amino acid content measured in vinasse shows that yeast consumed most of the amino acids.
Introduction
The industrial production of baker's yeast is an aerobic process based on propagating cells from pure culture to large bioreactors by increasing the volume in each propagation stage in sugar based medium. To overcome the well-known Crabtree effect, which results in lower biomass yield and ethanol production, 1 the dosage of sugar, usually molasses, has to follow a specified feed rate profile during the fed-batch fermentation in order to maintain its concentration below the critical value (50–100 mg/L). 2 This way, the specific growth rate is controlled by the feed rate profile of sugar. 3 Ammonia is the usual nitrogen source for this fermentation 4 and also has to be added according to a specified rate profile. In the latter phase of cultivation, the dosage of substrates is intentionally lowered or even stopped for one or both substrates. In this stage, substrate limitation occurs, and the cells switch from the exponential to the stationary phase leading to the accumulation of reserves in the form of glycogen and trehalose. This is a crucial phase during industrial baker's yeast fermentation that allows cells to survive the upcoming stress (osmotic and mechanical stress during conditioning and packaging) and to build up their storage stability. 5 Traditionally, feed rate profiles for molasses and ammonia are built on the basis of company historic data and are specific to a determined strain and process conditions (pH-value changes, temperature profile). 1 Despite of the long history of industrial baker's yeast fermentation, a full mass balance analysis has not been reported yet for this process. Such an analysis could be very beneficial for scientists and yeast producers aiming to improve their process or valorize the resulting vinasse.
In this study, vinasse is referred to as the cell-free broth obtained after cell separation at the end of the fed-batch fermentation. Cell separation is usually achieved by centrifugation and continuous filtration via rotary vacuum filters. Approximately 2.3 tons of liquid and unconcentrated vinasse is recovered per ton of baker's yeast. After concentration and partial precipitation, vinasse is often used in Europe as an additive or feeding supplement for ruminants and non-ruminants, 6 due to its influence on animal performance leading to an increase in daily weight gain. 7 Since the beginning of the 20th century, vinasse has also been used as field fertilizer, 6 due to significant content of macronutrients (nitrogen, potassium, calcium sulfate, magnesium) and micronutrients (iron, manganese, zinc, copper). 8 All over the world, its application in agriculture reported positive feedback to soil properties. 9
However, vinasse could play more important role as raw material for the production and recovery of key biomolecules such as betaine or technical enzymes. 10 The composition of vinasse originating from ethanol production has been widely studied in order to assess its potential for animal feeding and to eliminate the environmental pollution caused by this product. 11,12 Yalçin et al. have investigated nutritive values of modified dried vinasse from baker's yeast production intended to be used as animal feed. 13 Surprisingly, very little attention has been dedicated to the investigation of vinasse resulting from industrial baker's yeast fermentation and information regarding its composition could not be found in the literature.
The present study aims to partially compensate for the lack of reliable data regarding full mass balance analysis during industrial baker's yeast fermentation and the composition of the resulting vinasse. For this purpose, the mass balance of raw materials and products during the fed-batch fermentation of Saccharomyces cerevisiae was investigated by applying a fermentation protocol based on the scale-down of an industrial process. The detailed compositions of molasses and vinasse were analyzed, reported and compared to measure the degree of utilization of molasses nutrients during the fermentation. Furthermore, potential recovery of invertase, betaine, amino acids and food grade proteins as key biomolecules was discussed.
Materials and Methods
Strain
Large-scale yeast inoculum S. cerevisiae was provided by Uniferm GmbH & Co. KG (Monheim am Rhein, Germany).
Medium and Cultivation
A 1-L bioreactor (Sartorius, Biostat® Qplus, Göttingen, Germany) was filled with 0.5 L water and 2.7 g monoammonium phosphate (MAP) (Sigma-Aldrich, St. Louis, MS) and autoclaved at 121°C for 20 min (Autoclave Tuttnauer 5075 ELV, Biomedis Laborservice GmbH, Gießen, Germany). 466.05 g of a molasses mixture (10% sugar cane and 90% beet molasses) and 183.95 g of tap water (together with 25% sulfuric acid (Merck KgaA, Darmstadt, Germany) required to adjust pH-value to 5.4) was supplemented with a trace element solution and 300 μL of antifoam and autoclaved at 121°C for 20 min. Directly after the sterilization, the molasses mixture was placed on a shaker (IKA Shaker® KS 250, Staufen, Germany) at 150 rpm, aerated with air (2 slpm) for 1 h and then supplemented with vitamin stock. Medium's weight lost due to water evaporation during the sterilization and aeration was adjusted with sterilized water.
The fermentation carried out in this study was performed according to the method developed at Uniferm GmbH & Co. KG. In this regard, dosage profiles, pH profile, temperature profile and aeration profile are similar to those in large-scale industrial fermentations and enable maximum yield of yeast with high fermentative capacity and long storage stability. It is important to note that the mentioned fermentation process was optimized over several years to accurately reproduce the behavior of the industrial-scale bioreactor used every week by the company to prepare large-scale batches. The plausibility of the results has been discussed and positively evaluated by the personal of Uniferm GmbH & Co. KG. The bioreactor, filled with 500 g of water and 2.7 g of MAP, was inoculated with 18.47 g of yeast. The medium was then gradually added to the bioreactor and the fed-batch fermentation was started. After 20 h and 40 min of fermentation, the bioreactor was discharged and the yeast cake was separated from the vinasse by means of vacuum filtration. Molasses mixture (after the treatment) and vinasse, as by-product, have been further analyzed.
Determination of Dry Matter
1 mL of sample was pipetted, weighed (Mettler Toledo AT200; Columbus, OH) and heated for 24 h at the temperature of 105°C (drying oven, Shel Lab, Cornelius, OR).
14
Dried sample was once again weighted and the dry matter content (DM) was calculated according to the following formula:
where mdS = mass of the crucible with the dried sample (g); m = mass of the crucible (g); and VS = volume of the sample (L).
Determination of Ash Content
A muffle furnace (Nabertherm S27 Controller L3, Lilienthal, Germany) was used for the determination of ash content. 1 mL of the sample was pipetted, weighed and heated for 2 h at 550°C. Ash content was determined according to the following formula:
where mbS = mass of the crucible with the burnt sample (g); m = mass of the crucible (g); and VS = volume of the sample (L).
Determination of Betaine Concentration
Betaine concentration was determined by the method proposed by Chastellian with minor modifications.
15
The samples were acidified with 37% hydrochloride acid (Carl Roth, Karlsruhe, Germany) to pH 3. A column filled with 55 mL of a cation-exchange resin (Dowex® X8 50W 50-100, Alfa Aesar, Haverhill, MA) was washed with 75 mL of 2.5 N hydrochloride acid (Merck KgaA, Darmstadt, Germany) and subsequently neutralized with 75 mL of water prior to measurements. 50 mL of the sample (molasses mixture and vinasse) was injected in the column and eluted with 50 mL of distillated water. The column was then injected with 75 mL of 2.5 M HCl and washed with 75 mL of distillated water. Finally, column was acidified with 50 mL of 2.5 M HCl and 100 mL of distillated water. Fractions were collected and analyzed via nuclear magnetic resonance (NMR) spectroscopy (NMR-spectrometer AscendTM 400, Bruker, Billerica, MA). For this purpose, 700 μL of the corresponding fraction was filled in an NMR-tube and 300 μL of internal standard (tert-butylamine hydrochloride, 33.33 mg/mL (Sigma-Aldrich, St. Louis, MS) as well as 100 μL of deuterium oxide (Deutero, Kastellaun, Germany) were added. The betaine concentration was calculated according to the following formula:
where cB = concentration of betaine (g/L); cRef = concentration of the internal standard (g/L); Nref = number of protons in the internal standard; Nref = number of protons in betaine; Mx = molar mass of betaine (g/mol); Mref = molar mass of the internal standard (g/mol); Ax = integral value of the signal representing betaine; and Aref = integral value representing the internal standard.
Determination of Sugar Concentrations
The concentrations of sucrose, glucose and fructose were determined with commercial kits (Enzytec™ D-Glucose/D-Fructose/Sucrose, R-Biopharm AG, Darmstadt, Germany).
Determination of Protein Concentration
The protein concentrations were determined with commercial kit (BCA protein assay kit, Novagen®, Darmstadt, Germany). For the SDS-page electrophoresis (Mini-Protean® Tetra System, Bio-Rad, Munich, Germany), the samples were precipitated with 60% trichloroacetic (Merck KgaA) acid and then treated with 10x Tris/Glycin 16 and Laemmli buffer according to Laemmli, with minor differences. 17 The 5x Laemmli buffer contained 1.5 M Tris/HCl (Carl Roth, Karlsruhe, Germany), 0.01% Bromphenol Blue (Carl Roth) and none of 2-mercaptoethanol. The gel (7.5% Mini-PROTEAN® TGX™ Gel, Bio-Rad) with proteins was recorded with a ChemiDoc™ XRS+ System (Bio-Rad).
Determination of Amino Acid Content
All the chemicals used in this analysis were obtained from Merck KgaA. The samples were treated with methanol in ratio 1:4 and frozen overnight to precipitate the proteins. Proteins were removed via centrifugation at 14,000 rpm for 15 min (Heraeus™ Pico™ 21 Centrifuge, Thermo Fischer Scientific, Waltham, MA) and filtered (Sartorius™ Minisart™ PES Syringe Filter 0.2 μm, Göttingen, Germany). The samples were diluted with borate buffer (0.4 M) to reach pH 10. For the determination of cystin/cysteine concentration 100 μL of the sample was mixed with 250 μL of borate buffer (pH 8), 50 μL of 20 mM dithioerythritol, 50 μL of 400 mM methyl iodide and 100 μL of 4 M sodium hydroxide. After 10 min the reaction was stopped with 100 μL of 4 M hydrochloric acid and 350 μL of borate buffer (pH 10) was added.
Two mobile phases were prepared: A (40 mM monosodium phosphate pH 7.8 and 5 mM sodium azide) and B (acetonitrile, methanol and water in ration 45:45:10). Two derivatization reagents were prepared; OPA-reagent: 10 mg o-phthaldialdehyde, 6.5 mg mercaptopropionic acid in 500 μL methanol and 500 μl borat puffer (0.4 M, pH 10) and FMOC-reagent (9 mg/mL 9-fluorenylmethyl chloroformate in acetonitrile). Injection diluent was prepared with 100 mL of eluent A and 0.4 mL of concentrated phosphoric acid. The injected samples consisted of the following: 15 μL sample, 20 μL OPA-reagent, 20 μL FMOC-reagent and 50 μL of injection diluent. The high-performance liquid chromatography (HPLC) apparatus consisted of a pump (Agilent 1200, Santa Clara, CA), a gasifier (3 Canal gasifier, Sykam, Fürstenfeldbruck, Germany), an injector (Triathlon, Spark Holland BV, Emmen, Netherlands), a column heating device (Techlab T-1, Blacksburg, VA), an analytical column (Agilent Zorbax Eclipse Plus C18, 3.5 μm,4.6 mm x 150 mm), a detector (Shimadzu RF-10AXL Fluorescence Detector, Kyoto, Japan) and computer program (Clarity 5.0; DataApex, Prague, Czech Republic).
Determination of Invertase Activity
The invertase activity was determined by the method of Timerman with minor modifications.
18
One unit U of enzyme activity is defined as the amount of enzyme required to either hydrolyse 1 μmole of sucrose per min or produce 1 μmole of invert sugar per min. This method relies on the spectrophotometric determination of monosaccharides in the presence of an alkaline 3,5 dinitrosalicylate solution (DNS) (0.20 M NaOH (Merck KgaA), 23 mM 3, 5-dinitrosalicylate (Merck KgaA) and 0.53 M sodium potassium tartrate (VWR, Radnor, PA). 1 mL of sucrose solution (100 mM) (Merck KgaA) was mixed with 0.1 mL of the sample in a glass tube. After 5 min, 2 mL of DNS was added, the tubes were placed in boiling water for 10 min, and afterwards diluted with 3 mL of deionized water. The absorbance of the samples was measured at 540 nm (Spectrophotometer DR5000, Hach Lange, Düsseldorf, Germany). Finally, the invertase activity was determined according to Beer-Lambert law:
where I. A. = invertase activity (U/mL); ΔABS = absorbance value above the reagent blank; VTotal = total volume (6.1 mL); VSample = sample volume (0.1 mL); a = absorptivity constant (2.0 mL/(μmole cm)); b = cuvette light path (1 cm); and t = time or reaction (5 min).
Since the sample was brown colored and could absorb light at 540 nm, a blank had to be prepared to properly set the auto zero of the photometer. This blank consisting of 1 mL of deionized water, 0.1 mL of sample and 2 mL of DNS mixed with 3 mL of deionized after incubation.
Determination of Raw-Protein Content in Yeast
Quantitative determination of nitrogen was performed according to the method of Kjeldahl. 19 The raw protein content in yeast is calculated by multiplying with factor 6.25.
Determination of Residual Ammonium Ions in Vinasse
The concentration of residual ammonium ions in vinasse was determined with commercial kit (Ammonium cuvette test 2.0–47.0 mg/L NH4-N LCK, Hach, Düsseldorf, Germany).
Results and Discussion
Yeast Cultivation and Mass Balance Analysis
Figure 1 depicts the general mass balance of the fermentation process. During the feeding phase, 349.8 g of molasses mixture and 48.7 g of 10% ammonium solution were pumped into the bioreactor. 9.9 g of 25% sulfuric acid were required to correct the pH value during the fermentation. The cultivation yielded 241.7 g of yeast, with a dry matter content of 27.23% (approximately 7.4 g dry cell weight per kg of broth) and a raw protein content of 49.03%. The achieved high cell density and protein content are very similar to the values obtained by operating an industrial process. 20 After vacuum filtration, 645.8 g of vinasse were obtained.
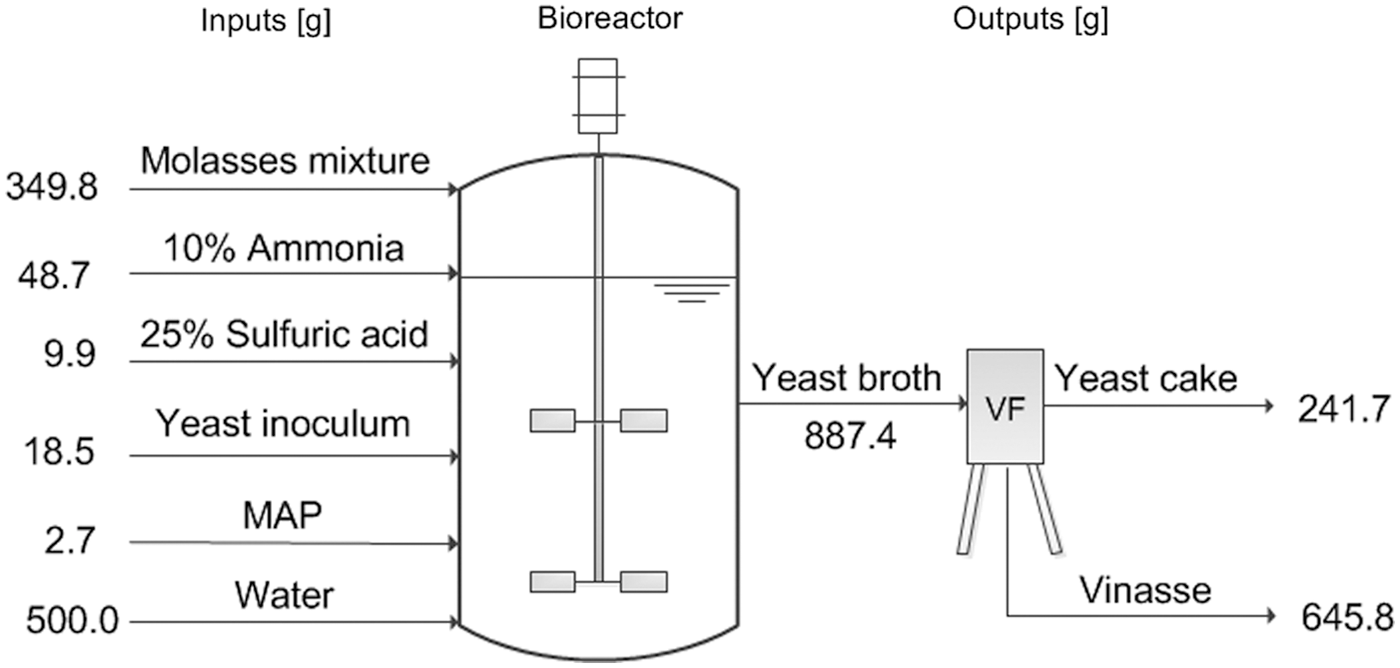
General mass balance analysis of the fed-batch fermentation performed in this study (MAP-monoammonium phosphate; VF-vacuum filtration).
Utilization of Sugars and Invertase Production
Absolute masses and contents of sucrose, glucose and fructose measured in the molasses mixture and the vinasse are summarized in Table 1. As expected, sucrose was found to be the prevailing sugar in the molasses mixture, followed by fructose and glucose. The sugar composition measured for this medium is in a good agreement with the literature. 21 22
Absolute Masses of Sugar (g) and Sugar Contents (g/L) Measured in Molasses Mixture and Vinasse
The yield coefficient of yeast biomass based on total sugar consumption (YX/sugar) was 0.584 g/g, which fits well with data reported in the literature. 20,23 Sucrose is a disaccharide that has to be hydrolyzed in order to be metabolized by S. cerevisiae. 24 Although a considerable portion of external invertase is located within the yeasts cell walls, portion is being released in the fermentation broth, where it accumulates. 25 External invertase (21.3 U/mL) was released in the cultivation medium by S. cerevisiae during the fermentation. In this regard, a yield of activity based on yeast biomass Yinv/X = 210.9 U/g was determined. Invertase is a key biomolecule which currently belongs to the most important technical enzymes and should be regarded as an attractive by-product of this process. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis confirmed the presence of external invertase, with a molecular weight of 270 kDa (Fig. 2). 26 Furthermore, only peptides and proteins with low molecular weights were found in the molasses mixture while the protein fraction of vinasse seems to consist of larger proteins which can be easily separated from other molecules by size exclusion (e.g., ultrafiltration).
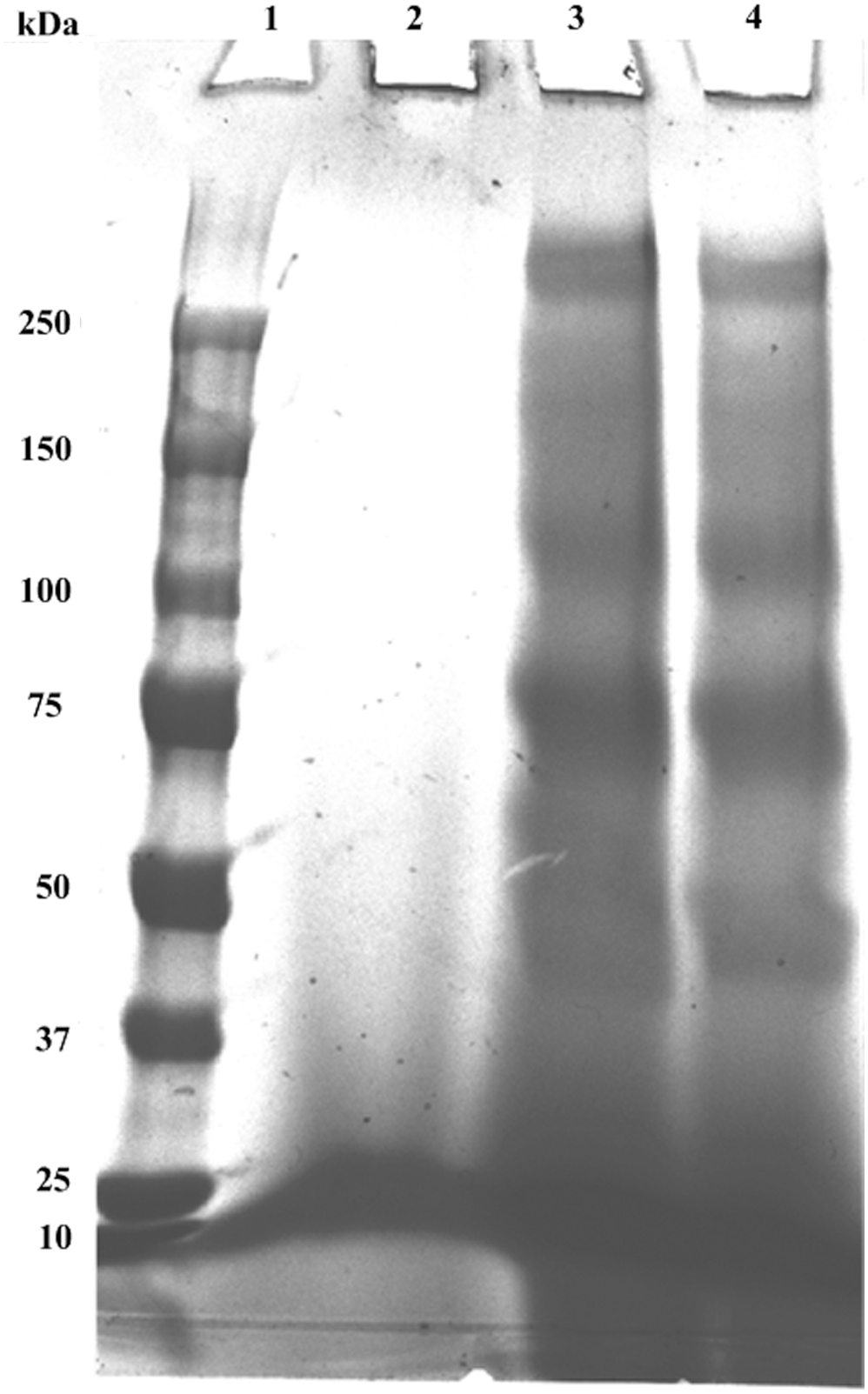
SDS-PAGE analysis. Lane 1: molecular weight markers (250–10 kDa); lane 2: molasses mixture; lane 3: vinasse with 2-mercaptoethanol; lane 4: vinasse (20 μL).
Utilization of Ammonia and Protein Production
As already mentioned 48.7 g of 10% ammonia were pumped into the bioreactor during the fed-batch fermentation and only 0.5 g of unassimilated nitrogen measured in form of ammonium ions were found in the vinasse. This finding confirms the substantial utilization of ammonia by S. cerevisiae and reveals that absolute nitrogen limitation did not occur during the late step of the process. Ammonia is then converted into glutamate and glutamine in order to be used as nitrogen source. 4 The yield coefficient of yeast biomass based on nitrogen consumption (YX/N) was 16.04 g/g. Baker's yeast utilizes the nitrogen source in anabolic processes for the synthesis of proteins, enzymes and nucleic acids. 27 Approx. 29.6 g of new synthesized protein were found in yeast biomass. According to Table 3, the mass of protein in molasses mixture and vinasse amounted 27.1 g and 19.2 g respectively. This in turn suggests that molasses protein and peptides were catabolized during the fermentation. The presence of proteins with higher molecular weight as well as the measured invertase activity indicates the release of new synthesized proteins in the fermentation medium due to controlled secretion or cell lysis.
Utilization of Amino Acids
It is known that amino acids are metabolized by S. cerevisiae. 20 The distribution of amino acids measured in the molasses mixture and the vinasse is summarized in Table 2. According to the literature, 28 amino acid with the highest content present in molasses is aspartic acid. Other amino acids having a majority share are asparagine and glutamic acid. A much lower absolute mass of amino acids was measured in vinasse (0.59 g in vinasse versus 2.56 g in molasses), which confirms the utilization of amino acids during the last hours of the fermentation when substrate limitation occurred. With exception of aspartic acid and glutamic acid, all amino acids present in the molasses mixture were almost fully metabolized. The considerable amount of glutamic acid might result from the combined utilization and production during ammonia conversion. Glutamine and histidine were not detected in the molasses mixture. However, they were measured in small amounts in the vinasse, meaning they were synthesized in the course of the fermentation. Glutamine could have been produced during the growth on ammonia, as mentioned above. In this pathway glutamate and α-ketoglutarate are being produced as well as histidine.
The Distribution of Amino Acids in Molasses Mixture and Vinasse Presented as Absolute Mass (g) and Concentration (g/L)
Mass Balance of Betaine
Betaine is a well-known amino acid derivative originating from beet molasses. The betaine content in molasses mixture was 53.91 g/L and represented 8% of dry matter weight, which is in agreement with literature data. 29 Approximately 70% of the betaine present in the molasses mixture was found in the vinasse. According to literature, betaine is not being metabolized to any significant extent by S. cerevisiae 30 and we believe that a part of the betaine remained trapped in pores of the yeast cake after vacuum filtration. Nevertheless, an appreciable amount of betaine remained in the vinasse (11.04 g at 18.5 g/L), which also has a considerable economic potential. Natural betaine is an important biomolecule in animal feeding that is expected to gain relevance over its synthetic form. Supplementation with natural betaine improves carcass lean deposition particularly under production stress, reduced the impact of coccidia challenge on intestinal lesion scores, positively affected nutrient digestibility and feed efficiency in broilers. 31
Overview of Molasses and Vinasse's Composition
The full compositions of molasses mixture and vinasse are shown in Table 3. Figures 3–4 depict the results of the chemical analyses of the molasses mixture and the vinasse and clearly show that the vinasse is much less complex medium than the molasses mixture. During the fermentation, baker's yeast “purified” betaine from the molasses mixture by assimilating almost all available sources of carbon. The composition of the vinasse gained from ethanol production has higher content on organic matter and contains sugar. Furthermore, it is characterized by a higher amino acid content and a considerable ethanol concentration. 11,12 The amount of invertase and betaine has not been reported yet for this kind of vinasse. However, it is important to state that the determined vinasse composition found in this work results from a specific molasses mixture (10% sugar cane molasses and 90% beet molasses).
Composition of Molasses Mixture and Vinasse Presented as Absolute Mass (g) and Concentration (g/L)
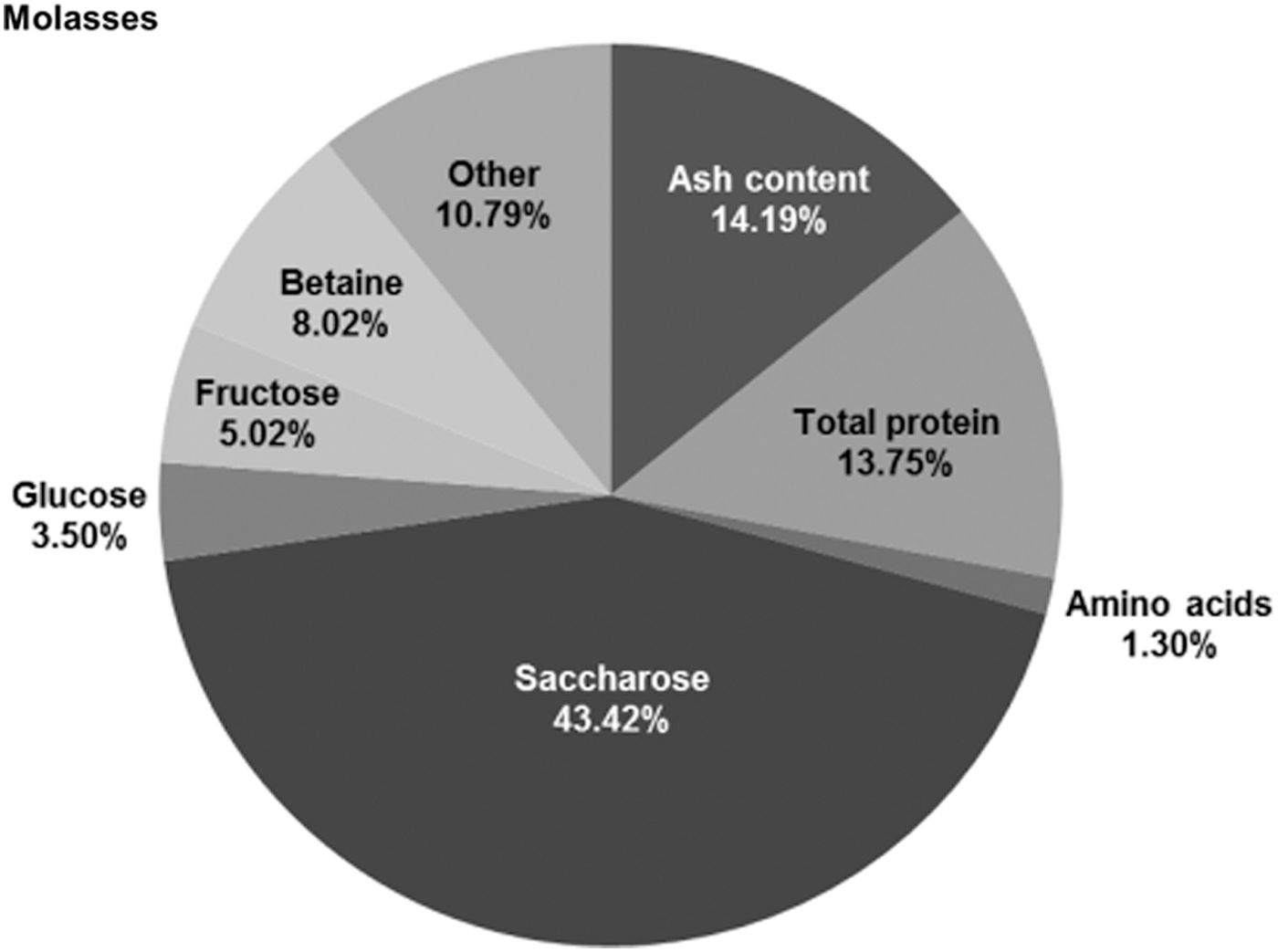
Composition of molasses mixture based on dry matter shown in percentage.
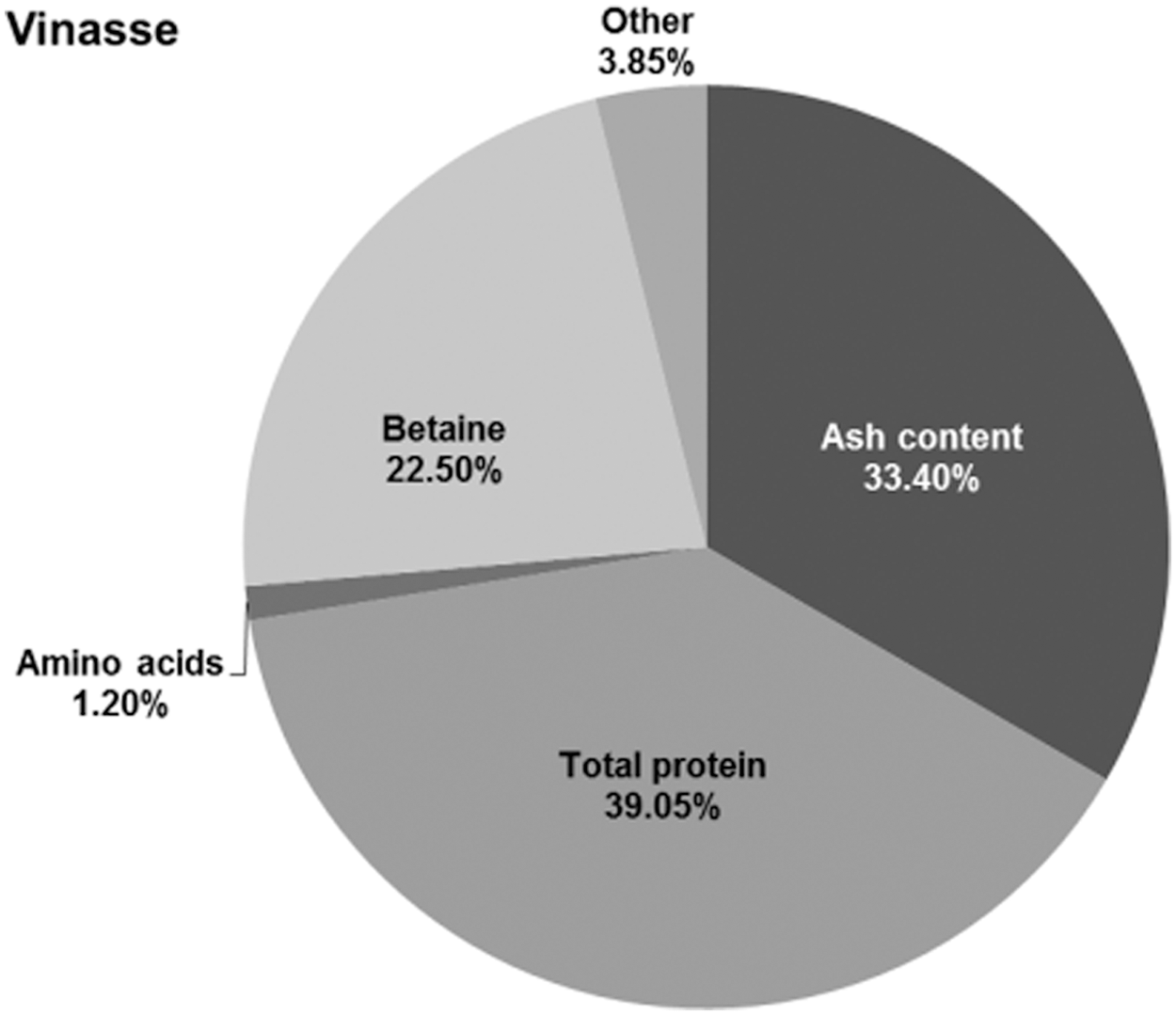
Composition of vinasse based on dry matter shown in percentage.
Conclusion
Considering that the fed-batch fermentation of baker's yeast performed in this study is a well-established process developed over the years by scientists and experts in Uniferm GmbH & Co. KG, the results obtained allow the upscaling to a large-scale process in the industrial environment. Annual production of 40,000 tons of compressed baker's yeast would generate 92,000 tons of vinasse that would need to be processed a bit differently than in the usual baker's yeast plant. External invertase, as a large glycoprotein with a molecular weight of approximately 270 kDa, can be separated and concentrated by ultrafiltration (e.g., 100 kDa molecular weight cut-off (MWCO)). The remaining protein fraction can be removed from small molecules (betaine, aspartic acid, glutamic acid and ashes) by an additional ultrafiltration step (e.g., 1 kDa MWCO). At neutral pH value, betaine exhibits zero net charge and does not bind on any ion exchanger. Under these conditions, aspartic acid and glutamic acid are negatively charged so that betaine purification (de-ashing and removal of aspartic acid and glutamic acid) can be achieved by ion exchange. According to the findings, it would lead to the bioproduction of approximately 1.8 x 1012 invertase units. The current market price for invertase involved in the production of invert sugar is approximately $2.5 for 106 invertase units (activity 20 U/mg). 32 Combining this price with the large vinasse volumes resulting from this process reveals a high economic potential for this by-product. In this industrial production, the recovered amount of food grade proteins amounts 2,730.4 ton/y. These proteins, which are obtained through valorization of baker's yeast vinasse, are high added-value products and as such are expected to have a higher average price of 7–8 kg/€ compared to traditional proteins. 33 Although baker's yeast consumes most of the amino acids available in molasses, still the appreciable amount could be recovered from vinasse (Table 4). The largest share of the amino acid market is consisted of feed amino acids used in the nutrition of swine, poultry and ruminants. 34,35 The recovered amount of theses amino acids from vinasse is 5.5 ton/y. Furthermore, 64.1 ton/y of amino acids used in human nutrition as food enhancers could also be recovered. Other amino acids (recovered amount of 14.2 ton/y) are employed as raw materials for synthesis of chiral active ingredients, which find application in pharmaceutical, cosmetics and agriculture industries. 34 The yearly volume of vinasse released by a baker's yeast production plant would allow producing up to 1,700 tons of natural betaine per year. With a current market price of approximately 1,000 $/ton, 36 recovering natural betaine from industrial baker's yeast fermentation could have a very high impact on the profitability of this process.
Yearly Production of Amino Acids for Feed and Food Sector
Footnotes
Acknowledgments
We would like to thank the laboratory staff from Uniferm, Monheim am Rhein for conducting the fed-batch fermentation as well as the analyses regarding molasses, baker's yeast and vinasse. We would also like to thank Martina Weiss, Gottfried Wilhelm Leibniz University, Hanover, for the measurement of amino acid profiles.
Funding Information
No funding was received for this article.