
Abstract
This work studied the optimization of carotenoids production by Sporidiobolus salmonicolor (CBS 2636) in a fed-batch bioreactor using coproducts (crude glycerol, corn steep liquor, rice parboiling water and yeast hydrolyzate). The maximum total carotenoids concentration, cell productivity and total were 4753 μg/L, 0.076 g/L·h and 48 μg/g, respectively, with a flow rate of 112 mL every 12 h, using a medium composed of 80 g/L crude glycerol, 80 g/L corn steep liquor and 20 g/L rice parboiling water at 25°C, pHinitial 4.0, agitation rate of 180 rpm, aeration rate of 1.5 vvm and 96 h of bioproduction. The specific maximum growth speed (μmax) was 0.052/h, obtaining the major carotenoid all-trans-γ-carotene (49%).
Introduction
Large demand on resources and growing consumer preference for natural products have increased interest in research concerning the production of carotenoids via biotechnology, which can be produced by bacteria, yeasts, algae and filamentous fungi. Among several types of microorganisms, red yeasts of the Sporodiobolus genus are advantaged by their unicellular nature, relatively high growth rate, and ability to accumulate both lipids and carotenoids, specifically β-carotene. 1 In addition, they are able to biosynthesize carotenoids from substrates containing mono and disaccharides and low-cost renewable raw materials (molasses, whey, rice parboiling water, corn steep liquor, crude glycerol, etc.) with optimum growth temperature between 25–35°C. 2 –7
Carotenoids are natural pigments, present mostly in red, orange and yellow food coloring. Their market was valued at $1.5 billion in 2014, with expectations to grow to $1.8 billion by 2019—an annual growth rate of 3.9%. 8 The interest in these pigments is due to their many applications; besides uses as a dye in food and animal feed, these compounds have antioxidant properties, which makes it possible to use them in preventive treatments for cancer, heart disease and macular degeneration. 9 –14
Bioprocesses are generally applied in large-scale production of natural pigments and dyes due to the higher cell density and higher growth rate of microorganisms. 15 Fed-batch processes are reportedly effective and versatile in most fermentations, including carotenoids production. In such processes, especially those with high cell densities, productivity is improved due to high viable cell counts. Fed-batch mode enhances control of substrate concentration while minimizing substrate inhibition. 7,16,17 Generally, any improvement achieved in carotenoid productivity by varying physico-chemical and nutritional parameters can significantly increase the yield of the product of interest. Therefore, it is of great interest to study the optimization of carotenoids production in a bioreactor, aiming to maximize yield and reduce costs of substrates by the best use of medium components. 18
In addition, the possibility of using low-cost agricultural by-products and/or residues as a raw source of carbon, nitrogen and minerals, among other nutrients, may increase the process economic viability. In the literature, there are reports of the use of agro-industrial substrates such as cheese whey, 3,19,20 grape must, 21 sugarcane processing derivates, 22 –24 corn processing derivates, 23 –25 and glycerol from biodiesel production. 6,7,26 –28 To this end, carotenoids production from by-products is a potentially viable alternative, and is in line with current strategies on the valorization of unexploited natural resources. 29
In addition, fed-batch fermentations are commonly used due to the substantial effect that varying substrate concentrations has on cell biomass and metabolite production in many fermentations. The feeding amount and feeding time also play very important roles in maximizing production of the desired products. 30 In the literature, fed-batch production of carotenoids by yeast is reported. 2 This work examines the fed-batch strategy using corn steep liquor, crude glycerol, rice parboiling water and yeast hydrolyzate as substrates for enhanced carotenoid production by Sporidiobolus salmonicolor (CBS 2636).
Materials and Methods
Conditions of Cultivation and Bioproduction of Carotenoids
S. salmonicolor (CBS 2636) (Centraalbureau voor Schimmelcultures, Utrecht, The Nederlands) was used in the bioproduction of carotenoids in a bioreactor. The lyophilized strain was hydrated in YM medium (Yeast Malt Extract): 3 g/L yeast extract, 3 g/L malt extract, 5 g/L bacto peptone and 10 g/L glucose, at 25°C for 72 h. The culture was transferred to slant tubes, containing Yeast Malt Extract Agar (YMA) medium with the same composition of YM medium as well as 20 g/L agar, and incubated at 25°C for 48 h. After growth, the slants were kept at 4°C and were subcultured for 2 months each.
The inoculum was prepared in Erlenmeyer flasks with 100 mL of YM medium. After sterilization, the flask was inoculated with a suspension of cells from the stock slants and incubated at 25°C, 180 rpm for 48 h (Optical density, OD600 = 0.7). 23 Carotenoid bioproduction were carried out for 96 h in a room without illumination in shake flasks with 100 mL of working volume and Biostat bioreactor (Braun Biotech International) with 1 L of working volume, with addition of inoculum 10% (v/v). Temperature was controlled using a water bath and pH was monitored during the bioproduction.
Agro-Industrial Substrates
The agro-industrial substrates used were corn steep liquor (CSL) donated by Corn Products (Mogi Guaçu, Brazil), parboiled rice water (PRW) acquired from Industrial Nelson Wendt (Pelotas, Brazil), crude glycerol (by-product of the conversion of oils into biodiesel) acquired from Industrial Olfar (Erechim, Brazil) and yeast hydrolyzate (Prodex Lac® Sd), donated by Industrial Bio Springer (Valinhos, Brazil).
The corn steep liquor was chemically pretreated using phosphoric acid according to the methodology described by Valduga et al. 23 The corn steep liquor was used at concentrations of 100 g/L. The pH value of the corn steep liquor was first adjusted to 3.0 using 1 mol/L phosphoric acid (Nuclear). The substrates were left for 24 h at 24°C and centrifuged (Eppendorf 5403) at 5,000 rpm for 15 min. Finally the pH value was adjusted to 5.5, using a 2 mol/L NaOH solution (Vetec).
Bioproduction in Shake Flasks
Initially, the effects of culture medium composition were evaluated in shake flasks. First, Plackett Burman (Screening Design) factorial design was used, where the independent variables investigated were crude glycerol (40–60 g/L), corn steep liquor (40–60 g/L), rice parboiling water (20–60 g/L) and yeast hydrolysate (0–5 g/L). Temperature (25°C), pHinitial (4.0) and agitation rate (180 rpm) were set at fixed levels 7 in an orbital shaker for 96 h. After statistical analysis, a second complete design 22 was used, where the independent variables (factors) studied were crude glycerol (65.9–94.1 g/L) and corn steep liquor (65.9–94.1 g/L), keeping constant pH (4.0), temperature (25°C), agitation rate (180 rpm) and rice parboiling water (20 g/L). The response or dependent variable studied was total carotenoids (μg/L), biomass (g/L) and yield of carotenoids (μg/g).
Carotenoids Production in Fed-Batch Bioreactor
In the fed-batch production, optimized medium in shake flasks was employed. An initial volume of 500 mL of the bioproduction medium was autoclaved in the Biostat bioreactor (Braun Biotech International) with 1 L of working volume. The feed rate was varied from 75 to 150 mL of medium each 12 h, maintaining fixed settings for aeration rate-air volume per volume of culture medium per minute (1.5 vvm), stirring rate (180 rpm), temperature (25°C) and pHinitial 4.0 for 96 h. 7 The fed-batch system was conducted with addition of the culture medium every 12 h considering the results in the growth curve (data not shown), where the biomass remained practically constant, this being the end of the exponential phase of cell growth.
Recovery of Total Carotenoids
The recovery of total carotenoids was carried out according to the method described by Valduga et al. 23 with modifications. After centrifugation, the supernatant fraction was removed and 10 mL of NaCl solution at 20% (w/v) and 10 mL of petroleum ether (Dinâmica) were then added. After stirring and phase separation was addition of sodium sulfate (Na2SO4, Merck) and a supernatant phase removed. The extracted fractions were evaporated on a rotary evaporator (Tecnal TE-210) at 35°C, and the pigments dissolved in methanol (Merck) for quantification.
Kinetics of the Carotenoid Bioproduction
The kinetics for total cell mass, carotenoid production, pH evolution, substrate consumption (glycerol; total nitrogen, TN; and total organic carbon, TOC in the medium) was followed by periodic sampling of the medium (12 in 12 h for a total period of 120 h). The experiments were performed in triplicate (n = 3).
The conversion factors (Yp/s, Yx/s and Yp/x), productivity in cells and carotenoids, instantaneous and specific rates (rx, rp, rsg, rsN, μx, μp and μs) were determined in the different runs performed in the bioreactor at fed-batch mode.
The conversion factor for the substrate in the product, Yp/s (μg carotenoids/g substrate), substrate in the biomass, Yx/s (g cells/g substrate), and the specific carotenoid production, Yp/x (μg carotenoids/g cells), were calculated by Equations 1, 2 and 3, respectively.
31
where rx is cell growth rate (g/L·h); rsg, rsN and rsC are glycerol, nitrogen and TOC consumption rate (g/L. h), respectively; and rp is carotenoid production rate (μg/L·h).
Instantaneous productivity in cells and carotenoids in a fed-batch system at constant volume are defined as the rx and rp rate, respectively. The rates of microbial growth (rx), product formation (rp) and substrate consumption (rs) can be determined by the mass balance for each component at a given time, as presented in Equations 4, 5 and 6.
31
The specific rate for growth (μx), product formation (μp) and substrate consumption (μs) can be obtained by dividing the instantaneous rate by the cell concentration, as expressed by Equations 7, 8 and 9.
31
Analytical Methods
Determination of total carotenoids
The absorbance of the sample after extraction was measured in a spectrophotometer (Agilent 8553) to 448 nm, using the equation described by Davies. 32 The coefficient of absorbance referent to β-carotene in methanol was used: E1%1cm = 2,550.
The concentration of carotenoids was expressed in terms of total carotenoids (μg/L) and specific production of carotenoids (μg/g). The specific production of carotenoids represents the total concentration of carotenoids (μg) in relation to the cell mass (dry weight) obtained in 1 L of a fermented medium.
Determination of cell mass
After extraction of the carotenoids, the cells were washed with distilled water and dried at 105°C (Fanem SE-320) until a constant mass was obtained. 7
Determination of pH
pH was determined using a digital pH meter (Digimed DMPH-2). 7
Determinations of total organic carbon and total nitrogen
The total organic carbon (TOC) and total nitrogen (TN) in the medium were determined by the catalytic combustion method, using a TOC/TN analyzer (Shimadzu model TOC-VCSH).
Determination of glycerol
The glycerol concentration was determined following the standard method UNE-EN 14105. 33
HPLC-DAD-MS/MS analysis
An aliquot of stock solution was prepared according to Rodrigues et al. 34 The carotenoids were analyzed in a high-performance liquid chromatograph (Shimadzu HPLC) equipped with quaternary pumps (model LC-20 CE), an on-line degasser and a Rheodyne injection valve with 20 μL loop. The equipment included a DAD detector (Shimadzu, model SPD-M20A) connected in series to a mass spectrometer with an atmospheric pressure chemical ionization (APCI) source and an ion-trap analyzer (Bruker Daltonics, model Esquire 4000). The carotenoid separation was carried out on a C30 YMC column (5 μm, 4.6 × 250 mm, Waters) with a flow of 0.9 mL/min and column temperature set at 29°C, the linear gradient of methanol/methyl tert-butyl ether (MeOH/MTBE) went from 95:5 to 70:30 in 30 min, to 50:50 in 20 min, was maintained at 50:50 for 10 min and finally modified to 95:5 in 2 min. The mass spectrometer parameters were set as follows: positive mode, current corona, 4000 nA; source temperature, 450°C; dry gas, N2 with a temperature of 350°C and a flow of 5 L/min; nebulizer, 60 psi. The MS/MS experiments were run in automatic mode, with fragmentation energy of 1.4 V. The mass spectra were acquired with scan range of m/z from 100 to 1,500.
The carotenoids were identified on the basis of elution order on the C30 YMC column, the UV–visible spectra features, such as spectral fine structure (% III/II) and intensity of cis peak (% AB/AII), and characteristics of the mass spectra (protonated molecule ([M+H]+) and its MS/MS fragments). The percentage of each carotenoid was calculated considering the total area of all identified carotenoids.
Statistical Analysis
The results obtained were performed in triplicate and treated by analysis of variance (ANOVA) followed by Tukey's test using Statistica® software (Statsoft version 5.0, Tulsa, OK).
Results and discussions
Carotenoids Production in Shaker Flasks
Table 1 presents the matrix (coded and real values) for Plackett-Burman factorial design (Screening Design) and response in terms of total carotenoids. The results indicated that the total carotenoids yield increased proportionately with crude glycerol and corn steep liquor contents, within the range studied. On the other hand, yeast hydrolyzate displayed a significant negative effect (p < 0.05) for carotenoids bioproduction, biomass decrease and suggesting an inhibition of pigments, in this way, this variable was excluded from the next stages of bioproduction (Table 2). Rice parboiled water did not present any significant effect, however, it showed a significant positive effect (p < 0.05) when associated to glycerol, that can be better viewed on the Pareto graph (Fig. 1) along with the variables estimated effects, in this way the variable PRW was set at 20 g/L (Table 2).
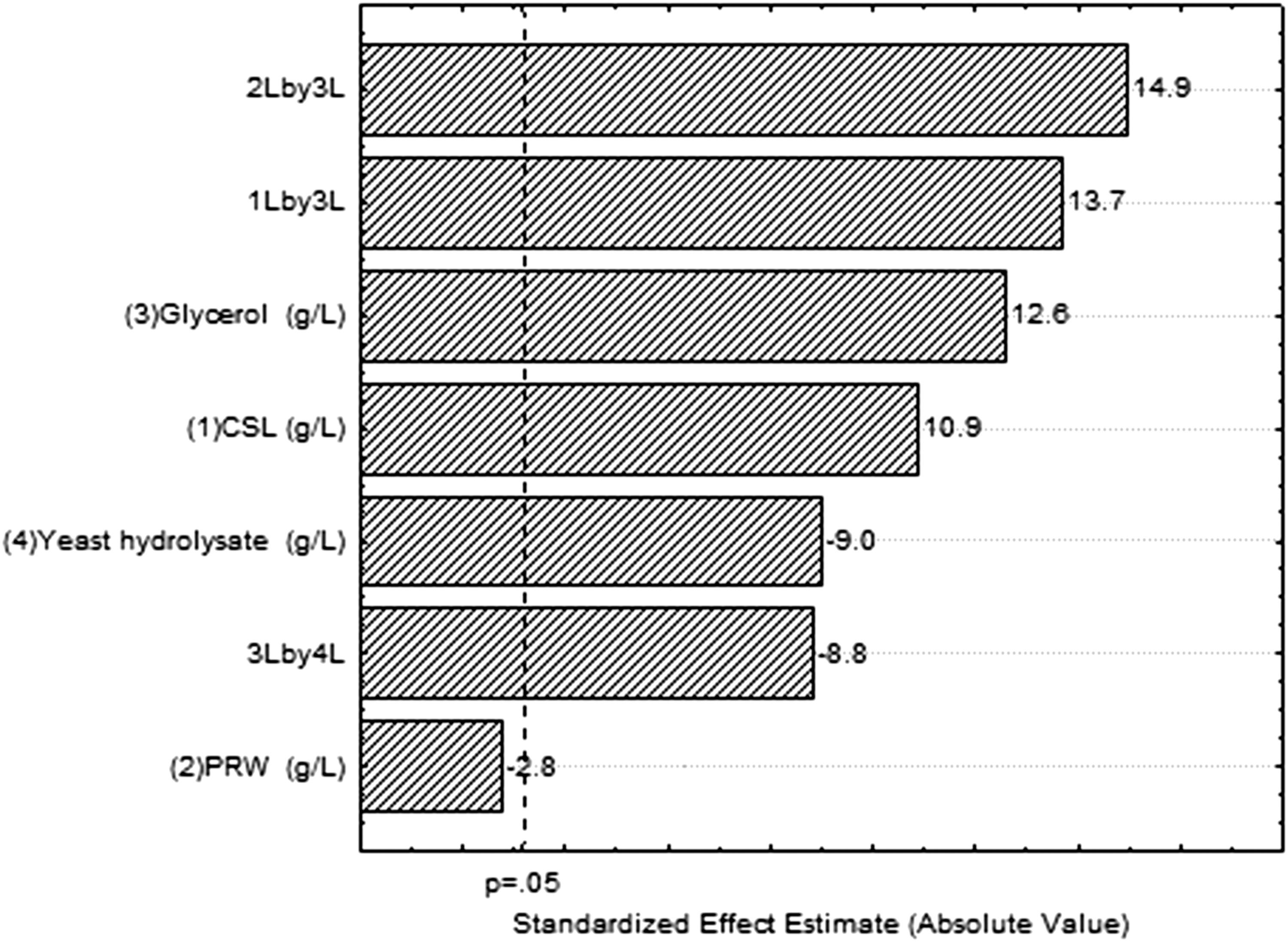
Pareto graph for the production of total carotenoids in shake flasks with coproducts as a function of the independent variables.
Matrix Plackett Burman (Real and Coded Values) and Response in Terms of Total Carotenoids and Biomass of Bioproduction with Coproducts in Shaker Flasks
X1 = CSL (g/L), X2 = PRW (g/L), X3 = Glycerol (g/L), X4 = Yeast hydrolysate (g/L); Fixed independent variable: 180 rpm, 25°C, 96 h e pHinitial 4.0.
Matrix Complete Experimental Design (Real and Coded Values) and Responses in Terms of Experimental and Predicted Total Carotenoids, Deviation and Relative Deviation and Yield of Carotenoids of Bioproduction with Coproducts in Shaker Flasks
X1 = CSL (g/L); X3 = Glycerol (g/L). Fixed variables: 96 h, 180 rpm, 25°C, pHinitial 4.0, 20 g/L of PRW.
Based on the results, a 22 complete factorial design was performed in order to optimize total carotenoids bioproduction. Table 2 presents the matrix (coded and real values) for the 22 complete factorial design and the response in terms of total carotenoids concentration. The highest carotenoids concentration was 2276 μg/L (540.65 μg/g cell), employing 80 g/L CSL, 80 g/L glycerol, and 20 g/L PRW (Run 9).
Equations 10 and 11 presents the second-order model, which described the total carotenoids concentration and yield of carotenoids as a function of the independent variables analyzed (factors), in the range studied. The models were validated by the analysis of variance obtaining a 0.97 correlation coefficient and calculated F value 6.5 times and 17.5 times greater than the tabulated one for total carotenoids and yield of carotenoids, respectively. Hence, allowing for the construction of contour curve (Fig. 2a and b) that presented the maximum total carotenoid production using a medium composed of 80 g/L crude glycerol, 80 g/L corn steep liquor and 20 g/L rice parboiling water (TOC/TN ratio of 35) at 25°C, pHinitial 4.0, agitation rate of 180 rpm, aeration rate of 1.5 vvm and 96 h of bioproduction. It can be seen that the relative deviation (Table 2) were low in the desired region where the total carotenoids are maximized, demonstrating that the model was adjusted the experimental data.
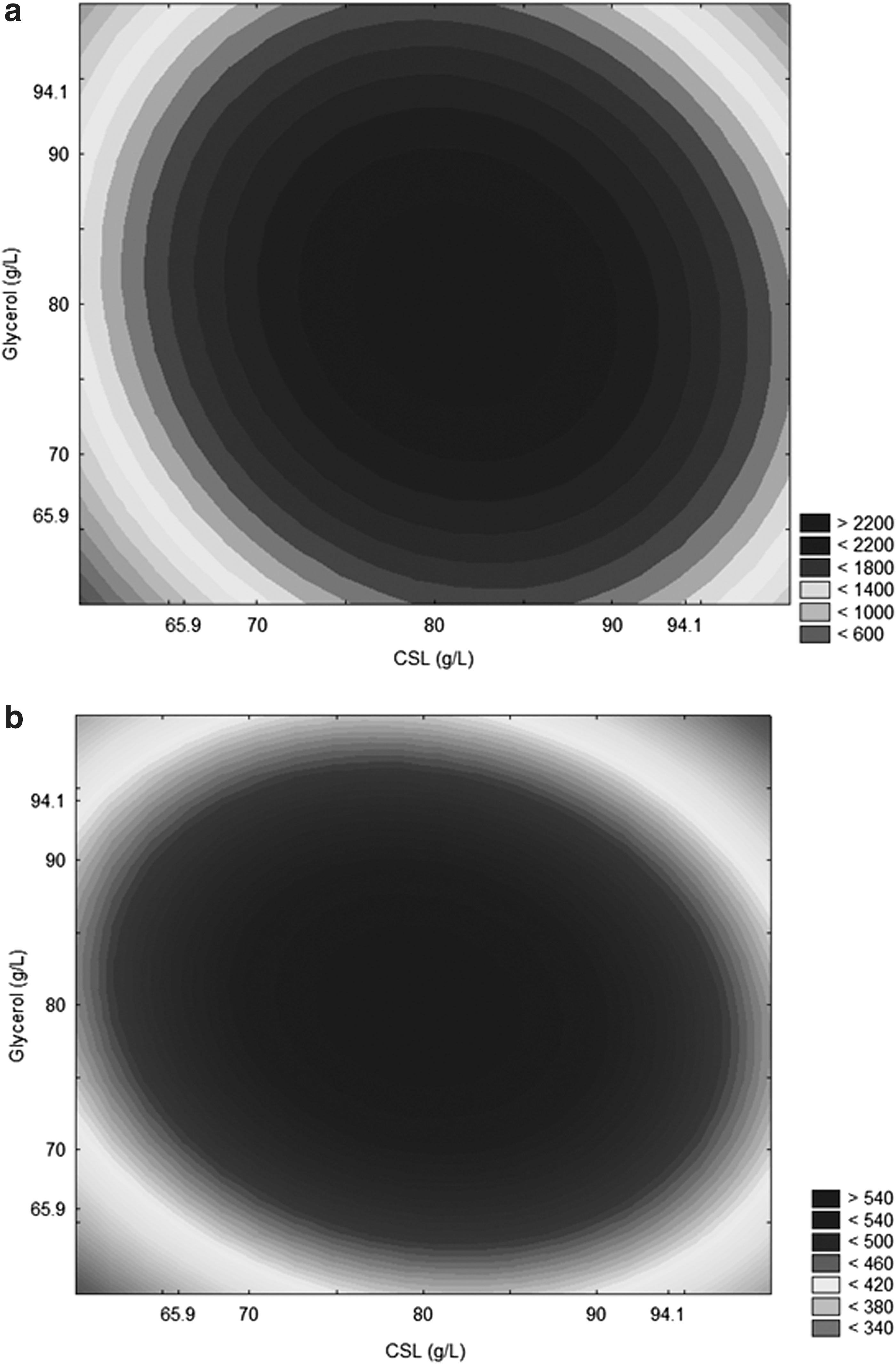
Contour curve for
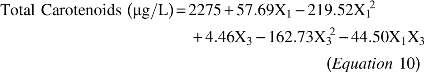

where: X1 = Corn steep liquor (g/L); X3 = Crude glycerol (g/L)
An increment of 50% in carotenoids bioproduction occurred using 22 complete factorial design (Table 2) in relation Plackett-Burman factorial design (Table 1). Thus, demonstrating that S. salmonicolor biosynthesizes carotenoids from crude glycerol and corn steep liquor, these sources promising carbon for yeast. Other studies report that the glycerol and/or corn steep liquor can be used as a source of carbon and increase its concentration to improve the production of carotenoids. Valduga et al. 6 optimized carotenoids bioproduction from Sporidiobolus pararoseus using the complete second-order design, obtaining a 843 μg/L total carotenoids maximum concentration (40 g/L glycerol, 40 g/L corn steep liquor, and 20 g/L rice parboiled water, at 180 rpm, 25°C and pHinitial 4.0). Machado and Burkert 27 optimized carotenoids bioproduction from S. pararoseus at 640 μg/L with a 22 rotatable central composite design, using 9 to 10 g/L of crude glycerol and 26 to 30 g/L of corn steep liquor at 25°C, 180 rpm, for 168 h. Manowattana et al. 35 compared the bio pigments production obtained from the by-product supplemented commercial medium with that from a synthetic medium, in which carotenoids production obtained from S. pararoseus (TISTR5213) yeast was 16.55 mg/L, 8.65 g/L biomass, with 35 g/L glycerol supplemented in BMP medium. On the commercial medium (YM), a 4.87 mg/L production with 6.68 g/L biomass was obtained at 24°C, 200 rpm, for 120 h, and pHinitial 6.0.
Carotenoids Production in Fed-Batch Bioreactor
Figures 3 –5 shows the cell growth kinetics (biomass), carotenoids production, pH variation and glycerol, TOC, and TN consumption from assays carried out in fed-batch bioreactor in an optimized medium with agro-industrial waste (80 g/L CSL, 80 g/L crude glycerol, 20 g/L PRW, and TOC/TN ratio of 35) at 75, 112.5 and 150 mL feeding rates every 12 h cultivation, respectively.
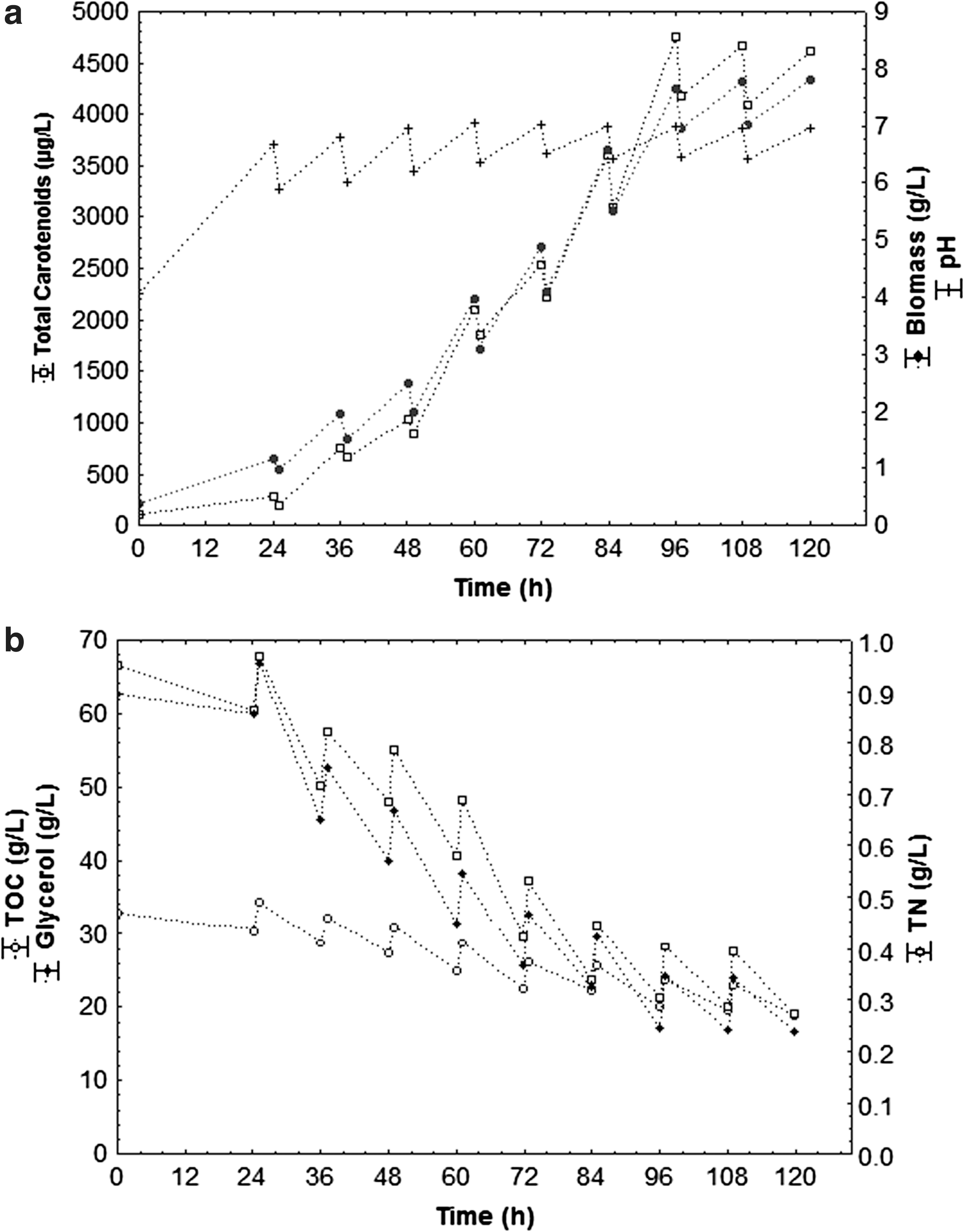
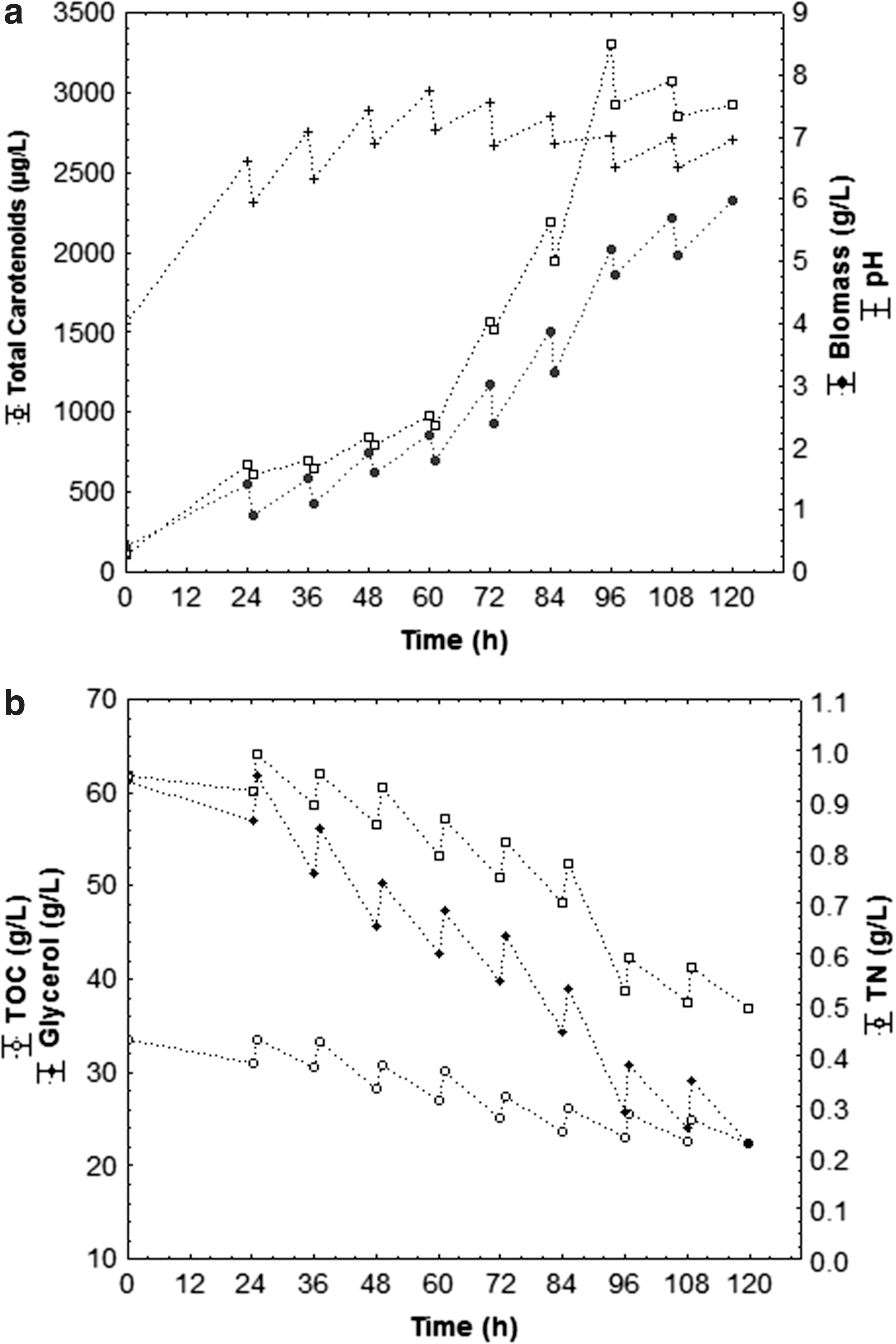
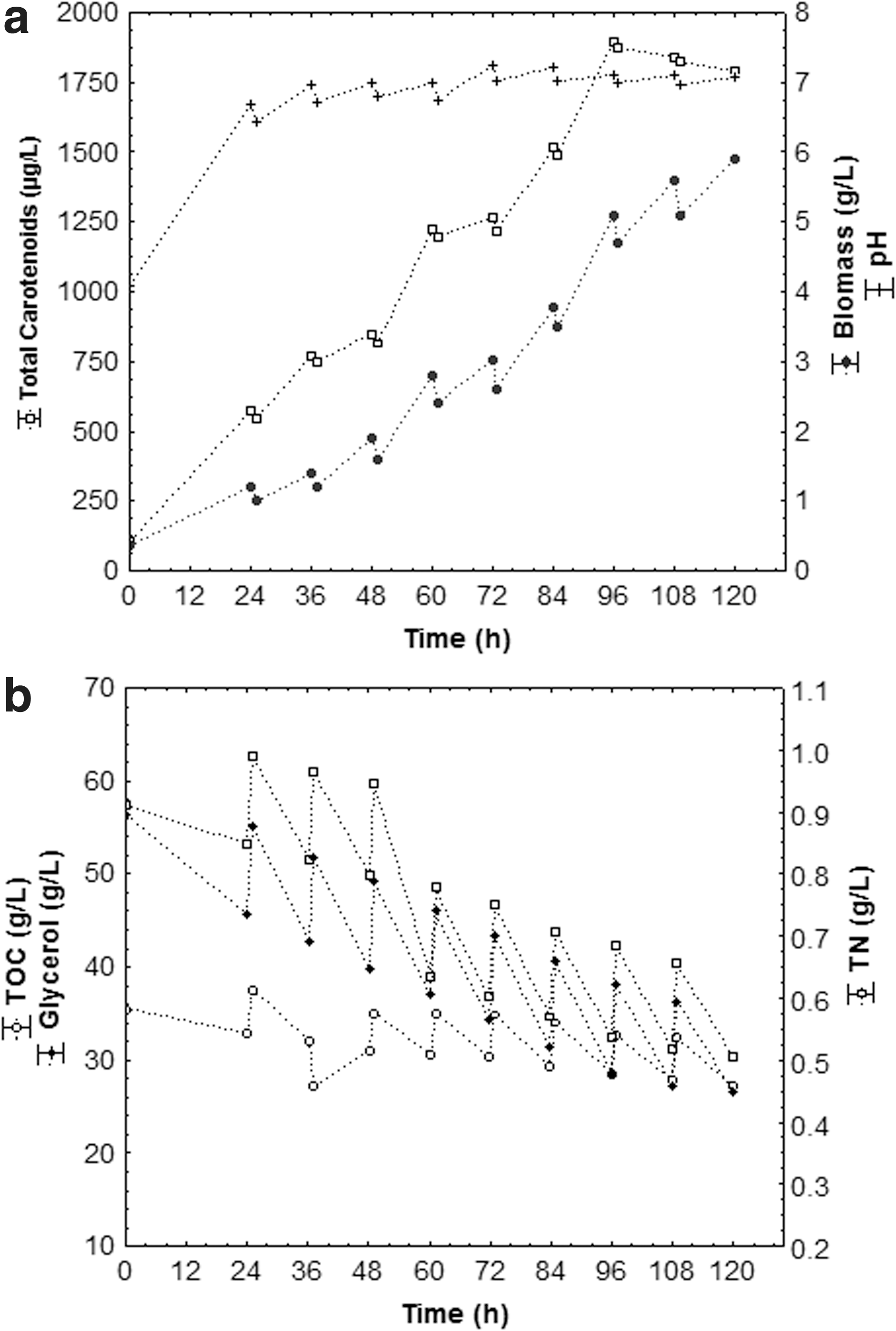
Kinetics of total carotenoids, biomass and pH (a); substrates consumption of glycerol, TOC and TN with feed rate of 150 mL every 12 h (n = 3) and the following operational conditions: 1.5 vvm, 180 rpm, 25°C, pHinitial 4.0.
The maximum carotenoids concentration (Fig. 3) was 4753 μg/L (639 μg/g), at 96 h bioproduction, with a 112.5 mL feeding rate every 12 h. However, the lowest total carotenoids concentration was 1894 μg/L (326 μg/g cell) at 96 h of bioproduction (Fig. 5), which received the highest fed volume (150 mL) every 12 h. In the 75 mL feeding rate every 12 h (Fig. 4) a decrease of carotenoids (3250 μg/L, 591 ug/g cell) occurred when compared to 112.5 mL, due to the low rate of feeding of substrates, which stimulated carotenogenesis under stress conditions. It could also be noted that total carotenoids production decreased after 96 h bioproduction, with ongoing substrate consumption after that period having no advantage in keeping bioproduction process for more than 96 h.
An increment of 108% in carotenoids bioproduction occurred using fed-batch bioreactor (Fig. 3) in relation to shaker flasks (Table 2). Yet, when comparing the fed-batch production with the one obtained by Colet et al., 7 which employed a medium containing commercial glycerol (80 g/L), malt extract (5 g/L), peptone (15 g/L), at a 112.5 mL feeding rate every 12 h (96 h bioproduction), there was an approximately 8% yield increase. Making it all the more interesting since they are low-cost and nutrient-rich raw materials and substrates, moreover, it is a viable alternative to industrial by-products reuse, such as crude glycerol, PRW, and CSL. In another study of the group 36 it was verified that the CSL has the highest levels of nitrogen (3,643 mg/L), sodium (2,850 mg/L), magnesium (640 mg/L), iron (10 mg/L), zinc (18 mg/L), copper (1.5 mg/L) and substantial source of organic carbon (34,010 mg/L). The glycerol has the highest concentration of total organic carbon (436 mg/L) and considerable amounts of calcium (256 mg/L), sodium (211 mg/L) and magnesium (32 mg/L), macro-nutrients that play important roles in cell metabolism and co-factors for various enzymatic reactions.
Carotenoids concentration is associated with the culture medium composition as well as to the bioproduction system. Saenge et al. 26 also noted the advantage of using a fed-batch system, whith the carotenoids production from Rhodotorula glutinis (TISTR 5159) being increased by approximately 13% when compared to the simple-batch system.
pH level is one of the most important environmental parameters influencing cell growth and product formation. As such, initial pH and its behavior were monitored during carotenoids production. Figures 3 –5 show that, independently of the fed volume, a similar pattern of pH was observed, with an increase during the first 24 h bioproduction, then small oscillations over the remaining period. Carotenogenesis and growth were induced by pH increase (pH 4.0 to 7.8), and pH 7.0 was ideal for cell growth and biopigments formation, with maximum carotenogenesis on the analyzed conditions (4753 μg/L, 639 μg/g cell) at pH 7.0 (Fig. 3). Lower level of residual glycerol at higher pH condition can be attributed to the higher metabolic activities of the cells leading to enhanced carotenoids in these cases
These changes in pH may be related to proteolysis of the microorganism, a natural phenomenon that occurs after complete substrate exhaustion as a consequence of amino acid degradation, with the formation of ammoniacal compounds that cause the pH to rise, principally when the strain cannot use another carbon source as reported in an another investigation using agroindustrial substrates (corn steep liquor, sugar cane molasses and yeast hydrolyzate). 3,6,37
Carotenoids bioproduction from S. salmonicolor (CBS 2636) yeast presented a carotenoids formation related to cell growth during the exponential phase (Fig. 3 to 5). With a 112.5 mL fed volume, every 12 h (Fig. 3), the highest cell concentration was 7.66 g/L for 96 h bioproduction.
Such high concentration of carotenoids was due to the fed-batch system, for when additional media is fed, cells are able to use more glycerol and mineral components and, as such, have more nutrients available to maintain their metabolism, this allows for a higher cell mass concentration, which was converted into a higher total carotenoids production, as well as specific ones.
Carotenogenesis is regulated in many microorganisms by nutritional factors, such as nitrogen availability. Nitrogen and carbon consumption rates play an important role in secondary metabolites synthesis, determining the types of synthesized metabolites and the production extension. 38,39 In this respect, it was noted from Fig. 3 to 5b, a gradual total nitrogen, total organic carbon (TOC), and glycerol consumption during the bioproduction process. Such pattern suggested that the cells consume glycerol as a carbon source. It could also be noted that there was higher nitrogen (71%), and total organic carbon (42%) consumption, with a 112.5 mL feeding rate, every 12 h (Fig. 3b) for 120 h bioproduction. Nevertheless, the lowest nitrogen (∼45%), and total organic carbon (∼24%) consumption were verified using 150 mL feed volume every 12 h (Fig. 5b). Colet et al.7 also noted higher TN (68%) and TOC (65%) consumption in the assay with 112.5 mL feeding for 96 h.
The gradual glycerol consumption during bioproduction was directly related to production time and the microbial growth phase, possibly affected by the absence of essential micronutrients for cell metabolism. 40 According to Figs. 3–5b, the glycerol consumption rate increased progressively during bioproduction on all assays. A greater glycerol consumption (74%) was verified with a 112.5 mL feeding rate every 12 h (Fig. 3b), and lowest (53%) at a 150 mL feeding rate, every 12 h (Fig. 5b). This was possibly related to residual glycerol excess in the medium from the assays where the fed volume was greater, causing cellular inhibition due to substrate overload.
Glycerol may provide both the major energy source for cell metabolism and the carbon element for biosynthesis of biomolecules. However, excessive glycerol was found to repress the synthesis of the carotenoids. The C/N ratio has a significant influence on cell growth and carotenoid biosynthesis in some microorganisms including yeasts. Most studies have suggested that a high C/N ratio is more favorable for the biosynthesis of carotenoids in these organisms. 26 Vustin et al. 41 reported that as the C/N ratio of a medium decreased, the increase in biomass was coupled with a decrease in the carotenoids (astaxanthin) production by Phaffia rhodozyma. Yamane et al. 42 proposed that a high initial C/N ratio may decrease the consumption of NADPH for primary metabolism such as protein synthesis, so as to leave more NADPH available for carotenoids (astaxanthin) biosynthesis.
Figure 6 shows the carotenoids productivity from fed-batch system on assays with 112.5, 75, and 150 mL volumes of medium fed every 12 h bioproduction. The maximum carotenoids productivity was 48.4 μg/L. h for 96 h, with a 112.5 mL feeding rate, every 12 h. Yet, at a 75 mL feeding rate, every 12 h for up to 60 h, a decrease in carotenoids production (14.4 μg/L. h) occurred, followed by an increase in carotenoids production reaching a maximum 33.3 μg/L. h for 96 h. That possibly happened due to low levels of nutrients available to the microorganism since this assay had the lowest feeding volume every 12 h. However, the maximum carotenoids production was 19.3 μg/L. h for 24 h at a 150 mL feeding rate every 12 h, remaining virtually constant until the end of bioproduction. The low carotenoids production occurred possibly due to inhibition caused by substrate surplus and a decrease in bioreactor's internal free area. 43 Colet et al. 7 noted a similar pattern with the different feeding rates, obtaining the highest productivity level (45 μg/L. h) with 112.5 mL, every 12 h in 96 h bioproduction.

Global total carotenoids production (Pcar) during bioproduction on the fed-batch process with feed rates of 112.5, 75 and 150 mL every 12 h, at 1.5 vvm, 180 rpm, 25°C, pHinitial 4.0 and 120 h (n = 3).
The maximum YP/X factor was 666 μg/g (Table 3) in 96 h, with a 75 mL feeding rate, every 12 h. According to Liu and Wu 44 the specific yield increase on carotenoids indicated an expansion on carotenoids biosynthesis from yeast cells. Such pattern was possibly related to changes in metabolism and cellular stress. YP/X in assay 3 decreased after 36 h bioproduction. That was possibly related to a greater raw material feeding, as well as to the low production values of total carotenoids and biomass, caused by raw material excess (higher feeding rate: 150 mL, every 12 h). According to Luna-Flores et al., 45 YP/X factor decreased during bioproduction due to substrate excess in the medium and a lower cells ratio to convert them into carotenoids.
Kinetic and Stoichiometric Maximum Parameters of Bioproduction in a Bioreactor from Tests in the Fed-Batch Process with the Agroindustrial Medium
The highest global conversion factors occurred in most assays during the first 24 h of bioproduction (Table 3) along with the highest observed pH increase (from 4.0 to 7.8). This explained why the cell conversion factor for glycerol, TOC, and nitrogen, and the product conversion factor for TOC and nitrogen were higher in the first 24 h. When the values obtained in shaker flasks with commercial medium 5 and in fed-batch bioreactor with commercial medium 7 are compared with the ones obtained in this study, it was verified that the differentiation in conversion factors emphasized the distinct cell growth pattern and substrates consumption using shaker flasks and fed-batch and/or simple-batch bioreactors.
The fed-batch process proved to be effective for the production of carotenoids. Fed-batch mode allowed the control of the addition of the substrates, minimizing inhibition by excess substrates, which allowed a higher concentration of cell mass and consequently a significant yield increase for the product of interest. Control of substrate addition is very important to achieve maximum output of the desired products for a batch process because it prevents underfeeding or overfeeding. In summary, the fed batch process is the preferred mode of operation due to the substantial increase in carotenoid production from S. salmonicolor, overcoming substrate inhibition in comparison to the batch process. 2,5,6,23 Three compounds separated by HPLC were identified or tentatively identified based on the combined information obtained from chromatographic elution on C30 column, UV–vis features (maximum absorption wavelength (λmax), spectral fine structure (% III/II), peak cis intensity (% AB/AII)) and mass spectra characteristics (Table 4). The MS/MS fragment characteristics of the polyenic chain and functional groups allowed the confirmation of the assigned protonated molecules. 46,47 The major carotenoid all-trans-γ-carotene (49%) could be observed.
Chromatographic, UV–VIS, Mass Spectroscopy Characteristics and Content of Carotenoids of Bioproduction in a Bioreactor from Tests in the Fed-Batch Process with the Agroindustrial Medium with Feed Rate 112.5 mL, Obtained by HPLC-DAD-APCI-MS/MS
Despite the availability of a variety of natural carotenoids from vegetables, microbial production remains an interesting topic due to the problems caused by seasonal and geographical variability in the production and marketing of plant origin dyes and colorants and there being an industrial interest in yellow carotenoids such as carotenes (β and γ-carotene).
Conclusion
The maximum carotenoids production in a bioreactor in a fed-batch system was 4,753 μg/L (639 μg/g), using a medium composed of 80 g/L crude glycerol, 80 g/L corn steep liquor and 20 g/L rice parboiling water (TOC/TN ratio of 35) at 96 h, with a 112.5 mL feeding rate every 12 h, 1.5 vvm, 180 rpm, 25°C and pHinitial 4.0, obtaining the major carotenoid all-trans-γ-carotene (49%). The fed-batch bioproduction resulted in a 2.1-fold increase in total carotenoid concentration as compared to the shaker flash.
Carotenoids production becomes an alternative use for crude glycerol, a biodiesel industry residue that is carbon-rich, low cost, and abundant, as well as for corn steep liquor and parboiled rice water.
The optimization of the fermentation process and the components of bioproduction medium (agro-industrial substrates) are important strategies to maximize the production and recovery of pigments.
Footnotes
Acknowledgments
The authors thank the National Council for Scientific and Technological Development (CNPq), Coordination for the Improvement of Higher Education Personnel (CAPES - Finance Code 001) and Research Support Foundation of the State of Rio Grande do Sul (FAPERGS).
Author Disclosure Statement
No competing financial interests exist.