
Abstract
The search for increased efficiency and lower costs for second-generation (2G) bioethanol production is generated by concerns regarding food safety and crop expansion, because the first generation of this fuel competes with production of food for consumption. There is also interest in decreasing associated greenhouse gas emissions and dependence on fossil fuels. However, processing 2G ethanol is expensive, inhibiting scale up of production. To produce this second-generation fuel, lignocellulosic materials need to be pretreated and hydrolyzed such that the fermentable sugars can be released from their polysaccharides. This biomass requires efficient enzymes to cleave cellulose and hemicellulose fractions (and sometimes pectin as well) into oligosaccharides (e.g. cellobiose) and monosaccharides (pentoses and hexoses) that depend on yeasts to convert them into ethanol via alcoholic fermentation. Fortunately, herbivorous insects are a potential source of enzymes and microorganisms to perform these functions, because they consume the carbohydrates from the lignocellulosic biomass. This review addresses various strategies for the bioprospection of enzymes (either endogenous or from insect microbiomes) and/or insect isolated yeasts; the efficiency of xylose and cellobiose metabolism of those microorganisms; successful experiments with such biocatalysts; and in particular, the economic impact of bioprospected enzymes and microorganisms exploited for 2G ethanol production.
Introduction
Faced with concerns regarding depletion of fossil fuels and the worsening of environmental problems attributable to excessive use of petroleum and its derivatives, worldwide attention has turned to the production of biofuels. 1 Bioethanol has garnered particular attention in the fuel market as a substitute for gasoline. The introduction of flex-fuel-powered vehicles into the market, as well as requirements on the part of some governments with respect to addition of ethanol to levels of up to 25%, has encouraged the increased production of bioethanol worldwide. 2 World production of bioethanol increased from approximately 22 to 122 billion L between 2000 and 2018. 3 The US and Brazil alone contributed 85% of global bioethanol production in 2018. 4
In Brazil, production is based primarily on the fermentation of sucrose from sugarcane. In the US, it is based on the fermentation of starch, derived mainly from corn. 5 Nevertheless, in addition to sugar- and starch-based sources, other materials may be used to produce bioethanol, including lignocellulosic biomass, which is rich in polysaccharides that may be hydrolyzed into fermentable sugars and consequently converted to ethanol—so-called second-generation (2G) ethanol. 6
This raw material is very diverse, and several types of agroindustrial waste, including bagasse, straw, stalks, leaves, grains, husks and even fruit residues, have the potential to be used as raw materials in 2G ethanol production. 7–8 These residues, produced in large amounts in most developing countries, where agriculture is the main economic activity, have little or no value in productive processes. 8–9 By contrast, the use of residues in the ethanol production, in addition to solving the problem of food versus energy (which is critical for first-generation production of this fuel), also contributes to the optimization of energy production and to the reduction of costs. 8,10
On the other hand, one of the current challenges for 2G ethanol production is making it economically viable. To reach this end, process productivity plays a crucial role. The concept entails achieving high concentrations of final product, with the highest yields and the least possible processing times. This requires glycoside hydrolases (GH) with high activity and low K m (high affinity), so as to generate product in short periods of time, even with low concentrations of substrate. Simultaneously, yeasts able to rapidly consume the hydrolysis-released monosaccharides are required to allow utilization of all (or almost all) the carbon source through fermentation (instead of metabolizing sugars through the Krebs cycle and the respiratory chain).
Another challenge is mitigation of the environmental impacts generated by the production processes, particularly those related to freshwater consumption. Despite the fact that 2G ethanol generates smaller water footprints than first-generation fuel (including freshwater for crop irrigation), 2G also demands high volumes of water to fill tanks and must preparation. 11 These needs have stimulated the search for alternatives to increase water safety during this process, including the constitution of very high-gravity musts (more concentrated, i.e., with more solutes per liter of water) or musts prepared with seawater instead of freshwater. 12,13 In order to fulfill this role, however, it is desirable to identify halotolerant enzymes and microorganisms, or at least biocatalysts capable of tolerating high osmotic pressures.
Therefore, the bioprospecting of enzymes and microorganisms in herbivorous insect guts has been proposed. Because these invertebrates absorb sugars as nutrients from lignocellulosic biomass, they rely on biocatalysts in their digestive tracts, either from the resident microflora or from their own cells (endogenous enzymes). The present review will address various strategies and successes in bioprospection, highlighting the main characteristics of the various insect enzymes and microorganisms described in the literature.
Main Challenges for 2G Ethanol Production
Agroindustrial wastes contain compounds with structures that hinder access of microorganisms and enzymes to the polysaccharide fraction available in plant cell walls, 7 resulting in cellulose, hemicellulose and pectin not being available for hydrolysis, in order to be converted into fermentable sugars. 14 The choice of pretreatment method, or the combination of different methods, depends primarily on the characteristics of the biomass. 15 –18 When the biomass is enriched with lignin, for example, it becomes more resistant to the pretreatment process, and although such characteristics require more aggressive pretreatment processes, it is important to point out that the degradation process of lignin-rich compounds can form furan compounds that may inhibit alcoholic fermentation. 19
Various physical, chemical and physicochemical pretreatment technologies have been employed for the production of second-generation ethanol from lignocellulosic biomass. Physical pretreatments increase the reactivity of cellulose in hydrolysis using microwave, extrusion, heat, ultrasound or a combination of these methods; however, these methods require higher energy consumption than chemical pretreatments. 20 –25 The chemical pretreatments are primarily acid or alkaline. The acid method, in addition to decomposing the lignin structure to increase subsequent cellulase accessibility, may cause hydrolysis of the hemicelluloses. Several acids have been used, including hydrochloric, phosphoric, acetic, formic, nitric, peracetic and sulfuric acid. 26 –30 The acid pretreatment may be carried out with diluted or concentrated acid, the latter being less indicated because of its potential inhibitors and corrosion problems, 31 observed primarily with the use of sulfuric acid at temperatures above 120°C. 32 Alkaline pretreatment breaks down bonds between lignin and hemicellulose, thereby allowing the removal of the former polymer as well as acetyl groups and uronic acids. Simultaneously, this method generates swelling into cellulose that reduces its crystallinity and degree of polymerization. 33 Such approaches are carried out using NaOH, although reagents as KOH or Ca(OH)2 may also be effective. 31 Among the physicochemical pretreatments, steam explosion is one of the most studied methods. 34 In this process, lignin and hemicellulose are converted into soluble oligomers using the high-pressure saturated steam autohydrolysis reaction, 35 affected mainly by temperature, chip size and moisture availability.
Finally, it is also worth noting that pretreatments may liberate compounds such as resins, fatty acids, phenols and tannins that are present in the agro-industrial wastes in small amounts, but are capable of inhibiting enzymes and microorganisms in the subsequent steps of hydrolysis and fermentation in 2G-ethanol production. 36,37 This is particularly true when treating the biomass at high temperatures and in acidic conditions, favoring the degradation of the sugars in furfural, hydroxymethylfurfural and later in formic acid and levulinic acid, contributing to the hydrolysis of the acetyl group in hemicelluloses to acetic acid. 38,39
After pretreatment, the hydrolytic stage or a combination of hydrolysis and fermentation in one single step (the simultaneous saccharification and fermentation process, SSF) takes place. Among the hydrolytic processes described in the literature, enzymatic hydrolysis is a less aggressive and greener method than alkaline or acid hydrolysis, because the enzymes act in milder conditions and generate less impactful waste. Furthermore, through enzymatic hydrolysis, no inhibitory compound is released into the must, thereby restricting the production of those compounds (as by-products of the process) to the pretreatment stage.
The high cost of enzymes makes 2G ethanol production relatively unattractive economically. 40,41 Therefore, enzymes from alternative sources are compelling strategies for increasing efficiency and reducing operational costs associated with this ethanol production step. 42
During enzymatic hydrolysis, various GHs act on cellulose, hemicellulose and pectin, releasing pentoses and hexoses that may be then promptly fermented by yeast cells. Nevertheless, despite the fact that current industrial yeast strains easily ferment glucose, the fermentation of pentoses generate much lower productivity than that of hexose, directly influencing the alcoholic fermentation yield. 43,44 This is due to the ability to naturally ferment xylose on the part of a few microorganisms, including yeasts of the genera Spathaspora 45 and Candida. 46 Therefore, the search for microorganisms able to efficiently metabolize pentoses has proved to be a challenge for further investigations to increase bioethanol productivity.
Improving 2G Ethanol Production with Endogenous Insect Enzymes
Herbivorous insects play a major role in biogeochemical cycles because of their high capacity for degradation of lignocellulosic biomass. 47 Throughout evolution, many herbivorous insects have developed the ability to produce endogenous enzymes capable of hydrolyzing lignocellulosic biomass. 48,49 The evolution of endogenous insect enzymes may be a consequence of horizontal gene transfer 50 or even of co-evolutionary processes between these invertebrates and their symbiotic microorganisms. 48,51 Ancestral gene duplication processes may have enabled some insects, including those in the family Phasmatodea, to specialize as folivores and to diverge from their omnivorous ancestors. 52 Shelomi et al. 53 observed high homology among endo-β-1,4-glucanases from distantly-related insect species, not only phasmids but also termites, cockroaches and crickets, suggesting a general distribution of cellulase genes in some endogenous endoglucanase-producing insects.
It should be noted that the degradation of a polysaccharide such as cellulose in the insect gut, as it occurs in the hydrolytic stage of the 2G ethanol production, depends on the synergistic action of various glycoside hydrolases: endoglucanases, exoglucanases and β-glucosidases. Cairo et al. 54 showed that, in the lower termite Coptotermes gestroi, synergy between β-1,4-glucanase (with endo- and exo- activity) and β-1,4-glucosidase (capable of hydrolyzing cellooligosaccharides) increased by 2.5- to 10-fold the complete hydrolytic activity of cellulose compared to the individual application of each of these enzymes. This result suggests that a single insect can provide various enzymes to compose an enzymatic complex that may be used in sugar-alcohol factories.
Tokuda et al. 55 observed that a species of flagellate-harboring termite (Reticulitermes speratus) expressed endogenous cellulase genes specifically in the salivary glands, whereas the termite species that do not have symbiotic organisms (Nasutitermes takasagoensis) showed cellulase gene expression only in the midgut. Even with a similar concentration of endo-β-1,4-glucanase activity and luminal volume in the midgut between xylophagous higher termites (N. takasagoensis, which digests cellulose without the aid of symbiotic protists) and lower termites (Coptotermes formosanus, which requires hindgut protists), the higher termites have shown to be able to ingest larger particles of lignocellulosic material. 56 High cellulolytic activity, similar to endo-β-1,4-glucanase, was observed in the foregut and midgut regions of Dissosteira carolina, even when compared to other insect groups. 49 Endogenous endoglucanases (glycoside hydrolase family 45) were also isolated from the wood-feeding beetles Oncideres albomarginata chamela 57 and Mesosa myops 58 as well as from the salivary gland of Reticulitermes speratus and gut of N. takasagoensis. 59 Endogenous endo-β-1,4-glucanase expression levels may differ over the life cycle of the insects, as observed in the termite Reticulitermes speratus. 60
Among all the insect endogenous glycoside hydrolases, β-glucosidases are probably the most prominent. Recently, several studies have characterized these enzymes, especially in termites. Feng et al. 61 found two endogenous β-glucosidases with different substrate specificities in the termite Coptotermes formosanus. Both presented maximum activity between 30°C and 40°C and optimum pH at 5.0, corresponding to a well-tolerated temperature and the optimum pH for yeasts—a desirable feature for SSF processes. More recently, Li et al. 62 characterized a cockroach β-glucosidase with activity against cellobiose, cellotriose and cellohexaose. Although it retains only 50 to 60% of its maximum activity in temperatures ranging from 30°C to 40°C, this enzyme has an optimum pH of 6.0, retaining more than 90% of its maximum activity at pH 5.0. Among insect endogenous β-glucosidases, two enzymes described in two studies by Uchima et al. 63,64 deserve to be highlighted. In the second article, the authors presented a β-glucosidase from the higher termite N. takasagoensis that displayed maximum activity at pH 5.5, and although it had showed a relatively high optimal temperature (65°C)—not a desired feature for SSF—it had retained almost 100% of its activity against 100 mM glucose. Even more impressive were the findings of those authors one year prior: a β-glucosidase from the lower termite Neotermes koshunensis with an optimum pH of 5.0 was stimulated 1.3-fold when glucose was present at 200 mM. 64 Considering that β-glucosidases are expected to be inhibited by glucose, 65 both β-glucosidases are excellent alternatives for the hydrolytic step in 2G ethanol production. Examples of prominent insect β-glucosidases are summarized in Table 1.
Remarkable Features of Microbial and Endogenous Enzymes Prospected in Insects
not named by the authors; b p-nitrophenyl-β-D-xylopyranoside; c p-nitrophenyl-β-D-glucopyranoside.
Proteomic studies have also evaluated the presence of endogenous β-glucosidases genes in insects, 66,67 highlighting their importance for biotechnological purposes through heterologous expression. 68 Currently in GenBank, 69 a total of 181 β-glucosidase gene sequences from insects are deposited, of which 113 are endogenous. Figure 1 shows the number of known sequences of β-glucosidase in the animal kingdom, highlighting the number of insect sequences.

Total sequences of β-glucosidase gene deposited on GenBank retrieved from the animal kingdom and endogenous β-glucosidase genes from insects (data obtained from
Within the class Insecta, the order Isoptera is very likely to be the most prominent for the decomposition of lignocellulosic biomass. Quoting Brune, 70 termite guts are “the world's smallest bioreactors.” Indeed, they are thought to assimilate a significant proportion of the cellulose (74–99%) and hemicellulose (65–87%) components of lignocelluloses they ingest. 47,71 Taking this into account, Ben Guerrero et al. 72 prospected and evaluated cellulases and hemicellulases from two Argentinian native termite gut digestomes: Nasutitermes aquilinus and Cortaritermes fulviceps. In that study, the authors found considerable digestion success was due to the cooperation between the endogenous enzymes and those expressed by their microbiota, because the combination in total gut extracts allowed hydrolysis of sugarcane bagasse and Napier grass thanks to their high endoglucanase and xylanase activities. In fact, this profitable relationship had been seen previously by other authors. 73 –75 As reviewed by Taggar, 71 there is compartmentalization of both endogenous and microbial cellulases, with the former ones found especially in the foregut, midgut and salivary glands and the latter ones localized mainly in the hindgut. Taken together, these features underscore the high potential of herbivorous insects to contribute to the improvement of bioethanol production.
Mining Enzymes on Insect Guts Microbiomes
In addition to endogenous insect enzymes, several species of herbivorous insects depend on the symbiotic relationships with bacteria, fungi and/or protozoa in their gut that are also potential sources of important enzymes for the degradation of plant biomass. In this symbiosis, the principle role of microorganisms is to subject the lignocellulosic structure to a variety of enzymes capable of delignifying the material and hydrolyzing its polysaccharide content, 76 primarily represented by cellulose and hemicellulose. Subsequently, the insect cells absorb the monosaccharides generated during the hydrolytic process carried out inside their gut.
Therefore, insect microbiota may be seen as a major source of biotechnological enzymes, especially for 2G ethanol production that uses residual biomass as raw material. As previously mentioned, in the context of 2G ethanol, it is necessary for the lignin to be separated from the polysaccharides for the latter to be hydrolyzed by glycoside hydrolases (GH), so that their monosaccharides can be fermented into ethanol. 7 Indeed, considering the similarity between the first two stages (pretreatment and hydrolysis) of the 2G ethanol production process and the digestion process of lignocellulosic biomass in the gut of herbivorous insects, several researchers have successfully sought microbial GH in these class of arthropods.
The biodiversity of the gut microbiota is directly linked to the specialization of the diet of each species of insects and to the various stages of its life cycle, 77 –81 directly reflected in the diversity of enzymes. This ends up stimulating bioprospecting of enzymes in the most varied of situations. From a termite gut protist cDNA library, Todaka et al. 82 cloned 11 cellulases with similar or up to 2.6-fold greater performance than Trichoderma reesei endoglucanase I (Genbank accession in M15665), a reference enzyme in the cellulose hydrolysis. Such considerable success was also achieved by Bashir et al., 83 however with enzymes obtained from termite endosymbiont cellulolytic bacteria. Moreover, in this case, the biocatalysts not only presented high activity against carboxymethyl cellulose, but also against the mannan and xylan polysaccharides (the main component of the hemicellulose fraction in grasses). In the beetle Anoplophora glabripennis, Geib et al. 84 suggested that the presence of a fungus related to the Fusarium solani/Nectria haematococca species complex may be associated with the lignin degradation process that occurs in the digestive tract of this coleoptera. In a study carried out with the cultured microflora of the larvae of the beetle Euoniticellus intermedius, Mabhegedhe 85 generated a cosmid library from which seven clones with cellulase activity were obtained. Shi et al. 86 collected gut juice from 2,000 third to fifth instar grasshoppers, 100 fourth to fifth instar silkworm larvae and 30 woodborer larvae and analyzed their cellulase and xylanase activities. In general, the enzymatic activities obtained from the grasshopper and woodborer samples were significantly higher than those of the silkworm under various conditions. These authors also verified that both pH and temperature had significant impacts on the enzyme activities in the gut contents. In addition, it is worth noting the xylanase activity of approximately 5,500 μM mg−1 min−1 in woodborer larvae gut juice at 30°C and pH 7.0 as well as the cellulase activity of approximately 480 μM mg−1 min−1 in grasshopper gut juice at 50°C and pH 7.0.
Despite the fact that these examples demonstrate the substantial potential of these enzymes in industrial ethanol production, the bioprospection of enzymes of biotechnological interest from the insect microbiomes has been enhanced in the last decade by the application of metagenomic tools. Because it allows access to genetic material even from non-cultured microorganisms, metagenomics allows the screening of up to a 100-fold higher number of microbial species. 87 –92
To carry out metagenomic analysis in an insect gut, the insect surface needs to be sterilized before beginning the dissection process. Subsequently, the gut is longitudinally cut and the lumen content is gently squeezed into a buffer. 93 From this content, it is possible to extract the DNA directly from the resident microorganisms without the need of prior culture to construct metagenomic libraries. 94 With the library assembled, enzyme screening can be performed using two general strategies: sequence- or functional-based approaches, 92 as seen in Figure 2.
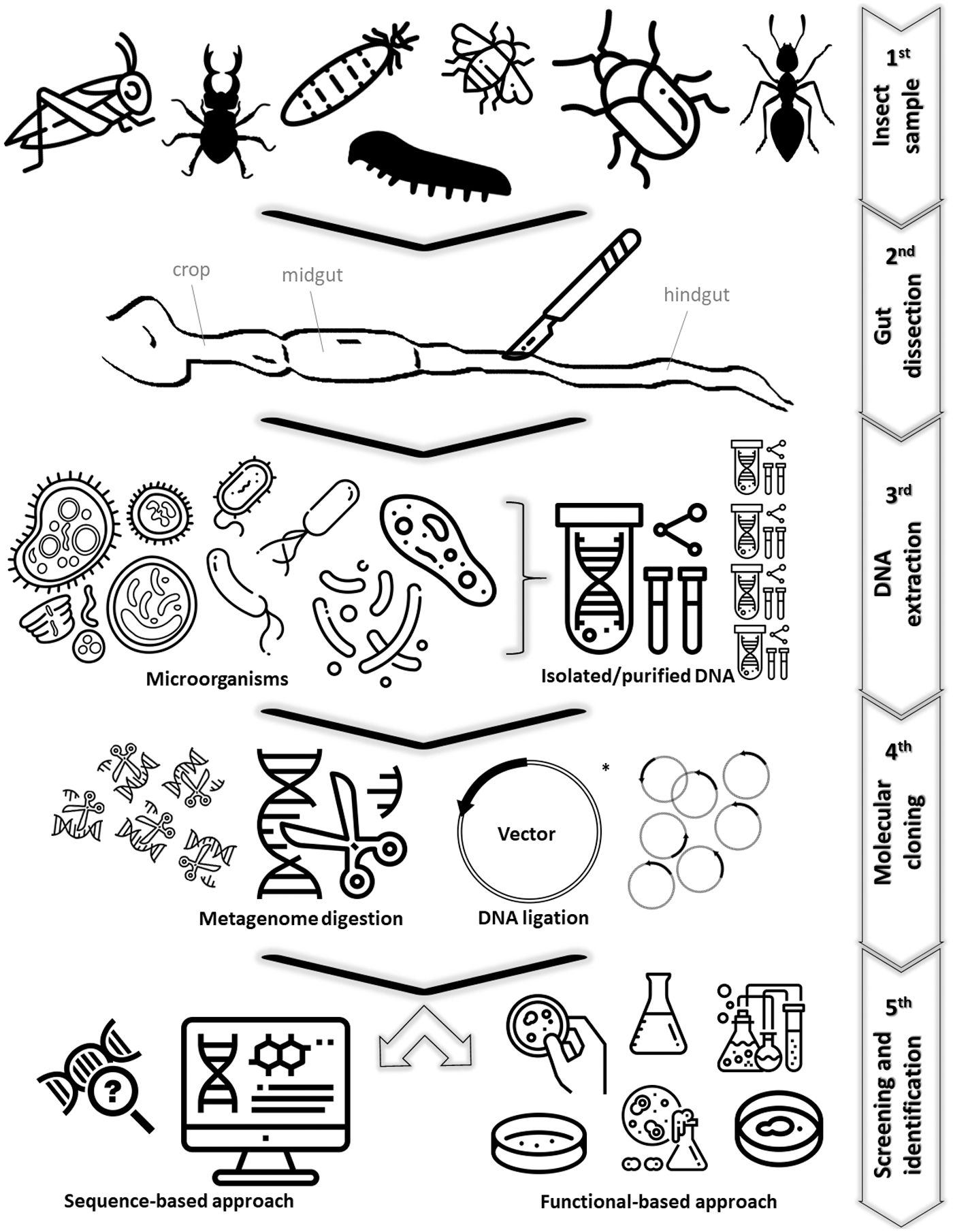
Bioprospecting enzymes using insect gut microflora metagenomes. Icons were retrieved from Flaticon, except plasmids* (by Cornelia Scheitz) that were retrieved from the Noun Project, and insect gut that was drawn based on Reid et al. 95
Using amplicon and shotgun metagenome sequencing, Scully et al. 96 analyzed the midgut community derived from third instar larvae of the invasive beetle Anoplophora glabripennis (Asian longhorned beetle) collected while they were actively feeding in the heartwood of sugar maple trees (Acer saccharum). The authors detected 4,394 reads of enzymes belonging to 36 families of glycoside hydrolases, with GH1 (∼13%), GH3 (∼16%) and GH32 (∼9%) being most dominant families. It is interesting to note that these three families of GH contain representatives capable of promoting the hydrolysis of cellulose and xylan as well as of their derivative oligosaccharides. In fact, the high number of carbohydrate active enzyme in the microbiome of herbivorous insects has already been observed by other authors. Shi et al. 78 dissected 2,000 third to fifth instar grasshoppers and 50 fourth to fifth instar cutworms to extract the genomic DNA of their gut symbionts. The metagenomic DNA was then analyzed and compared to the termite gut metagenome (JGI IMG Database GOLD ID: GM00013 and Sample ID: GS0000048). 97 In this analysis, the authors found a total of 31, 40, and 52 different GH families in the grasshoppers, cutworms and termite metagenomes, respectively. The termite gut featured the most abundant putative cellulases and hemicellulases, with 125 GH5 cellulases and 101 GH10 xylanases. Liu et al. 94 combined large-scale functional screening of fosmid libraries, shotgun sequencing, and biochemical assays to interrogate the gut microbiota of about 100 whole guts of wood-feeding termite Globitermes brachycerastes, and detected polysaccharide-degrading activities in 464 clones from the assembled metagenomic cosmid library. Of this total, they identified dozens of clones with endoglucanase (267), exoglucanase (24), β-glucosidase (72) and endoxylanase (101) activities. Therefore, the metagenome of only one insect provided enzymes able to hydrolyze the three major polysaccharides in the residual plant biomass: cellulose, hemicellulose (xylan) and pectin.
During hydrolysis of the lignocellulosic biomass, a set of enzymes must act in synergy to enable the release of monosaccharides resulting from polysaccharide breakdown. In the case of cellulose, glucose (the final product of cellulose hydrolysis) inhibits β-glucosidases, 65 reducing the efficiency of the hydrolytic process and, consequently, decreasing the yield of ethanol per ton of biomass. In this scenario it is worth noting the high glucose-tolerant glucosidase Glu1392 found in a hindgut microbiome of the termite Coptotermes formosanus that presented an inhibition constant (K i) value of 2.25 M for glucose. This glucosidase retained more than 80% of its maximum activity at concentrations of up to 1.0 M glucose. 98 For this, those authors dissected ∼500 worker termites to isolate the DNA and construct a plasmid metagenomic library in the Escherichia coli DH5α as the cloning host.
Another enzyme that deserves to be highlighted is the bi-functional cellulase purified by Hatefi et al. 99 from cultured bacteria (Bacillus sp.) isolated from the digestive tract of third instar larvae of the beetle Osphranteria coerulescens, collected from infected apricot and almond gardens. The enzyme characterized by these authors was stable in a wide range of temperatures (from 20°C to 60°C) and presented its maximum activity at pH 5.0, making it an excellent choice for SSF processes, given the need to meet temperature and pH ranges amenable to yeasts. In addition, the cellulase characterized by Hatefi et al. 99 was stimulated by Na+, thereby demonstrating its potential for application in alternative processes of 2G ethanol production that aim to reduce the water footprint by using sea water as a total or partial substituent for the freshwater employed in bioreactors. 12,13
In the context of the search for cleaner, renewable energies, third-generation (3G) ethanol is beginning to gain attention. 3G ethanol is derived from algal carbohydrate biomass as the substrate for alcoholic fermentation. 100 This raw material, however, may present polysaccharides different from those of terrestrial plants, requiring different hydrolases for the hydrolytic step. In red algae, mannan is one example. 101 This polymer must be cleaved by mannanases to release mannose residues to be fermented by yeasts. Through the analysis of gut microflora metagenome from ∼1,000 larvae of black soldier flies (Hermetia illucens), Song et al. 102 discovered an endo-1,4-β-mannanase belonging to the GH26 family, with high activity against galactomannan. Interestingly, when tested against various organic solvents, this enzyme maintained more than 90% of its maximum activity against 10% (v/v) ethanol. Considering that SSF processes currently have a certain predilection to produce cellulosic ethanol, 103 the tolerance of hydrolytic enzymes to ethanol becomes an extremely desirable feature. In this way, insects may contribute to the production of third-generation ethanol.
Many of the putative enzymes found in the metagenomic libraries of insect gut microbiota have been tested and characterized, demonstrating their high potential for biotechnological application. Examples of enzymes (prospected from insect microbiomes or from the own insect cells—endogenous enzymes, as previously discussed) with potential application in the production of bioethanol are summarized in Table 1. It is worth noting that some of them have extremely desirable features for process that aim to achieve low water footprints, considering their high-salt tolerance (halotolerant enzymes). Moreover, most of the enzymes presented in Table 1 also present characteristics that made them suitable for SSF, because they are highly stable and active in pH and temperature ranges that meet yeast requirements for efficient alcoholic fermentation.
It is important to highlight that some of the enzymes in Table 1 showed better performance than some commercial enzymes. Uchima et al., 63 for example, compared the thermostability and glucose tolerance of G1mgNtBG1 with those of the most common commercial β-glucosidase preparation from Aspergillus niger, Novozym 188 (Novozymes A/S, Bagsvaerd, Denmark). After 1 h of incubation at 60°C, G1mgNtBG1 retained 93% of its total activity while Novozym 188 was quickly inactivated and retained only 24% of its total activity. Regarding inhibition by glucose, while G1mgNtBG1 retained ∼100% and 50% of its activity at 0.1 M and 0.5 M glucose respectively, only 26% and 5% of Novozyme 188 activity was retained at the same concentrations of this hexose. 63 In Table 1, there are other examples of β-glucosidases (Glu1392 and CgBG1) whose thermostability and glucose tolerance are higher than those of Novozyme 188, and one of these was even stimulated by such monosaccharide (G1NkBG). Therefore, at least four β-glucosidases listed in Table 1 appears to be a better choice than the very common Novozyme 188.
Normally, commercially-available β-glucosidases are used as supplements in enzyme cocktails for hydrolysis of plant biomass, 111 with Novozyme 188 commonly employed with Celluclast® (Novozymes A/S) that consists of cellobiohydrolases and endo-1,4-β-glucanases. 112 –114 Considering the current predilection for processes whose hydrolysis step occurs simultaneously with fermentation, 103 some features presented in Table 1 are better fits for SSF than Celluclast. According to Herlet et al., 113 Celluclast retains 50–65% of its maximum activity at 35°C and pH 5.0 depending on the cellulosic analyzed substrate. On the other hand, as shown in Table 1, the cellulase described by Hatefi et al. 99 shows maximum activity at pH 5.0 and almost 90% of it at 30°C, and the endo-1,4-β-glucanase Bh-EGase II 105 retained ∼70% and ∼90% of its maximum activity at 30°C and pH 5.0, both pH and temperature levels being optimum for yeast cells. Therefore, various mixes of insect β-glucosidases and cellulases may show better performance in terms of hydrolysis of vegetal biomass than the commonly used commercial mix Novozyme 188 + Celluclast, especially in SSF processes.
Cellobiose- and Xylose-Fermenting Yeasts in Plant-Associated Insects
Research in pursuit of new microbial strains is relevant primarily because of their predisposition to produce enzymes of high commercial value, thereby constituting alternatives that reduce biotechnological costs. The screening of new strains usually occurs via isolation of microorganisms of interest in the most varied habitats and hosts. 115,116 In fact, the microbiological biodiversity existing in the natural ecosystem is huge; however, identification of new strains with industrial value generates significant impact on the development of globally-relevant bioprocesses, including 2G ethanol production. 117,118
In the case of biodiversity, insects can be considered intermediates in a large microbial community forming consortia of symbiotes among these constituents, including bacteria, yeasts, filamentous fungi and protists. Indeed, the environment of the insect intestinal microflora is extremely abundant in organisms that produce enzymes capable of metabolizing lignocellulosic material that are ingested as food. 116,119,120
Therefore, the insect gut may also be considered a source of various yeast species. 121 –123 Yeasts are linked to environments that have sugar sources, and that makes herbivorous insects a perfect niche, because these invertebrates feed on rich-carbohydrate biomasses. 124 –126 Studies with wasps, beetles and termites have shown that most yeast species consumed by these insects survive and remain in the gastrointestinal system and in the cuticles, thereby making the insects vectors of new yeasts among the most varied ecosystems. Therefore, isolation of yeast from the intestine of insects is a promising alternative for discovering new strains with potential for various applications. 126 –129
Insect-isolated yeasts present the possibility of producing a series of enzymes essential for a range of industrial processes, including those that allow metabolism of xylose (the main product of hemicellulose hydrolysis and the second-most abundant sugar in biomass hydrolysates) as well as hydrolysis of cellobiose (a disaccharide found on cellulose hydrolysates). 130,131 For the fermentation of the pentose, yeast cells must harbor a xylose reductase (XR) and a xylitol dehydrogenase (XDH) that share the same coenzyme—i.e., that allows NADH/NAD+ recycling—as well as an overexpressed xylulokinase. These three enzymes work in sequence, allowing the conversion of xylose into xylulose-5P that then accesses pentose-phosphate pathway before generating glycolysis intermediates to enable the conversion of pyruvate into ethanol. 43 Regarding cellobiose, normally the third-most abundant sugar in biomass hydrolysates after glucose and xylose, 132,133 yeasts need to express a β-glucosidase that can be secreted by the cells or located either in the periplasm or in the cytosol. β-glucosidases can also be part of a cellulase complex; however, in such cases, it has been shown that it may be insufficient for the amount of cellobiose formed during the hydrolysis of cellulose. Therefore, this disaccharide accumulates and may inhibit the action of endo- and exoglucanases by negative feedback, reducing the saccharification rate. 134 –136 In this scenario, a direct metabolization of cellobiose by yeasts during the production of 2G ethanol is crucial for the optimization of the process. 137
Depending on the yeast species, its use in the 2G ethanol production can be as only an enzyme supplier—because their enzymes can be heterologously expressed in industrial yeast strains already employed in first-generation ethanol plants—or directly as the fermenting microorganism itself. In fact, yeasts of the genera Blastobotrys, Candida, Pachysolen, Hamamotoa, Meyerozyma, Nematodospora, Pichia, Scheffersomyces, Schizosaccharomyces, Spathaspora and Sugiyamaella have been found in insect guts (especially beetles and termites) and were shown to be able to ferment xylose. 116,118,127,138 –144 Some of them showed high efficiency in converting this sugar into ethanol (Table 2). Furthermore, most of the aforementioned genera (especially Candida and Spathaspora) have representatives that can hydrolyze and ferment cellobiose. 138,145–146 Table 2 summarizes the most notable yeast fermentative performances in xylose- and cellobiose-containing media.
Xylose- and Cellobiose-Fermentative Performances of Yeasts Isolated from Different Insect Guts
Percentage of the total sugar available to the yeast cells; bRatio between ethanol concentration (g L−1) and xylose and/or cellobiose consumed (g L−1); cPercentage of the maximum theoretical yield of ethanol, considering 0.46 gethanol/gxylose and 0.538 gethanol/gcellobiose 149–150 ; dCofermentation of xylose and cellobiose.
In the Brazilian ethanol industry, musts present on average 150 to 200 g L−1 of fermentable sugar. These concentrations can cause osmotic stress in many yeasts, thereby decreasing fermentation yields, 151 –156 because the carbon source is eventually diverted to glycerol synthesis. 157 In fact, it has been demonstrated that current Brazilian industrial yeasts have adaptations that make them more apt to these conditions, including greater copy numbers of SNO/SNZ genes involved in the sequential synthesis of pyridoxine and thiamin. 152 Therefore, a high resistance to osmotic stress caused by high sugar concentrations becomes a desirable feature of the fermenting microorganism. The yeasts presented in Table 2 were subjected to various concentrations of xylose, from 50 g L−1 to 150 g L−1. Especially for S. passalidarum, an increase in the concentration of this pentose from 90 g L−1 to 150 g L−1 did not cause any detrimental effect on the fermentative yield of this yeast 116,118,138,148 ; to the contrary, ethanol yields in the context of xylose concentrations were approximately 10% higher (Table 2).
Economic Potential of Insect Enzymes and Yeasts
Currently, endogenous and symbiotic insect enzymes are not yet commercialized. However, insects have high economic potential, because of the growing demand for enzymes capable of degrading lignocellulosic materials. In addition to their uses in the food and beverage, household care, and pharmaceutical industries, the growing demand for renewable fuels, especially 2G ethanol, has driven the market for enzymes aiming for higher efficiency of the production process combined with reduction in costs. 158 The global enzymes market was valued at $7.08 billion in 2017, and is projected to reach $10.5 billion by 2024, with an annual growth rate of 5.7% over this period. 159 The biofuel enzymes market was worth over $900 million in 2018, and is forecast to grow annually at around 9.0% to reach $1.5 billion by 2024. 160
However, enzymes should never be considered in isolation, but rather as a part of a biocatalyst technology. Microorganisms also play an important role in the ethanol production. Currently, Fermentec (Brazil) 161 has the largest bank of Saccharomyces cerevisiae in the world, with more than 2,800 yeasts isolated from industrial fermentation in several different regions. Fermentec and LNF Latin America 162 have commercially available three-selected yeasts responsible for 70% of all ethanol produced in Brazil: CAT-1, PE-2 and FT858. Between 1997 and 2007, Fermentec showed a net profit of around $3.5 billion, only from the increase of the ethanol efficiency production in 33 plants. 163 According to the authors, in 2007, the company introduced customized yeasts that allowed greater stability of the fermentation process because of the greater adaptation (more persistence and dominance) of the yeasts isolated and reintroduced at the same plant. Over the subsequent seven years, the production of ethanol by these yeasts increased to over 3 billion L, representing a fermentation yield improvement from 75% in 1977 to 92% in 2014. 163 Customized yeast by Fermentec increased from 2 to 29 strains over the last ten years. 164 Furthermore, in addition to S. cerevisiae, very recently other yeasts 165 –169 and filamentous fungi 150,170 –172 have been shown to be efficient in the production of ethanol, either by improving hydrolysis of lignocellulosic biomass or by fermenting sugars such as glucose, cellobiose and xylose.
Enzyme and microorganism bioprospection in insect guts for 2G ethanol production may be, thus, considered a profitable biotechnology market, since one single insect organism may perfectly work as provider of enzyme complexes and yeast strains that can increase productivity and reduce costs in this biofuel production, transforming the dream of second-generation ethanol into reality.
Final Considerations
The biofuels industry has glimpsed the possibility of locating in insects a way to increase the economic viability of 2G ethanol production, because herbivorous insects emerge as a great supplier of biocatalysts capable of fully guaranteeing the production of this second-generation biofuel. The target enzymes for sugar-alcohol factories are produced inside insect guts, either endogenously or by their intestinal microbiota. Several examples of success, presented in this review, confirm the potential of their hydrolases to ensure that the hydrolytic stage of the 2G ethanol production occurs with high efficiency, considering costs, water footprint and reaction time. Moreover, the potential of insects to optimize bioethanol production is not restricted to the bioprospection of enzymes. Several yeast species have already been isolated from their guts and have demonstrated the ability to ferment the sugars found in lignocellulosic hydrolysates. Some of these yeasts have a fermentation efficiency higher than 90%. Therefore, enzymes and yeasts found in insects complement each other in such a way that a single insect species might provide all the biocatalysts necessary for optimized production of second-generation ethanol.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Brazilian agencies CNPq, CAPES and FAPERGS.