
Abstract
The heterologous expression of Vitreoscilla hemoglobin (VHb) is known to enhance short-chain-length (scl) and short-chain-length-medium-chain-length (scl-mcl) polyhydroxyalkanoate (PHA) production in Escherichia coli, Aeromonas hydrophila and Pseudomonas putida, but has not been evaluated for the production of medium-chain-length (mcl) PHA in pseudomonads. Three recombinant strains of P. putida KT2440 were constructed, harboring pBBR1MCS-2 expression plasmids containing 0, 1 and 2 copies of the vgb gene. The vgb gene was placed under the control of the lac promoter, which conferred constitutive expression of vgb, since no lac repressor was produced. Mcl-PHA production was assessed in nonanoic acid-limited, fed-batch fermentations with exponential feeding. Although the VHb levels in the engineered strains were comparable to those observed in vgb-bearing pseudomonads, the recombinant strains had lower biomass production, PHA accumulation, volumetric productivity and product yield when compared to the wild-type strain. The performance of the plasmid-harboring strains decreased with increasing vgb copy numbers, presumably due to an increased metabolic burden arising from plasmid expression. All the fermentations ended at similar times due to insufficient oxygen supply and subsequent uncontrollable foaming, demonstrating that VHb production did not alleviate oxygen limitation. These results indicate that the effectiveness of VHb engineering must be assessed on a case-by-case basis.
Introduction
Polyhydroxyalkanoates (PHA) are biodegradable biopolymers produced from renewable resources with potential to substitute traditional petroleum-based plastics, which are persistent and accumulate in the environment. 1 The thermomechanical properties of PHA make them suitable for a diverse range of applications, such as in compostable packaging, as well as in the biomedical and pharmaceutical fields. 2,3 Medium-chain-length PHA (mcl-PHA) are a class of PHA having repeating units of six to fourteen carbon atoms. Their diversity and elastomeric properties are better suited to high value-added applications. However, their commercialization is hindered by high production costs. Before its widespread use, PHA must be economically competitive with established petrochemical plastics. The economic drawbacks of PHA are mainly attributed to expensive carbon feedstocks, low process productivity and inefficient downstream strategies. 4 Considerable research efforts have been directed towards increasing PHA market competitiveness through process optimization, aiming mainly at increasing polymer yield and volumetric productivities. As PHA accumulate intracellularly, high cell density is required to achieve commercially viable PHA production, and oxygen limitation is the main constraint to obtaining high cell density. This is usually addressed by increasing the bioreactor's oxygen transfer rate and/or managing the substrate uptake rate.
One strategy to influence oxygen uptake is to metabolically engineer microorganisms for the heterologous expression of bacterial hemoglobins. Among these, Vitreoscilla hemoglobin (VHb) is the most widely studied. VHb is a soluble, homodimeric, cytoplasmic hemoprotein capable of reversible oxygen binding. It allows its strictly aerobic native host, Vitreoscilla stercoraria, to thrive in oxygen-deprived environments. VHb-based bioengineering has been shown to be beneficial to the bioremediation of persistent organic pollutants, 5,6 to the synthesis of industrially relevant microbial products, 7 –10 in addition to providing resistance to submergence in higher plants 11 and hypoxia tolerance in vertebrates. 12
The expression of vgb was found to be beneficial to PHA synthesis by different heterologous hosts. VHb-producing Escherichia coli strains had 3.4-fold higher cell growth and 4.2-fold higher poly(3-hydroxybutyrate) (PHB) accumulation at low dissolved oxygen concentrations. 13 Using an arabinose-inducible construct, a 1.2-fold increase in cell density and a 1.9-fold increase in PHB concentration was observed. 14 VHb production was also observed to have a positive influence on the production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx) in Aeromonas hydrophila 4AK4 with an increase of 33% in biomass concentration and 39% in PHBHHx content. 15 –17 Recombinant Halomonas strains also benefited from vgb expression, presenting increased cell growth of up to 100% and intracellular PHB content of up to 85%. 18
Mcl-PHA are mainly synthesized by fluorescent Pseudomonas species. The expression of vgb has been demonstrated in P. aeruginosa and P. putida, 19,20 and was shown to slightly enhance cell viability in P. aeruginosa and Xanthomonas maltophilia. 21 The biodegradation of benzoic acid was favored by VHb production in X. maltophilia 22 and P. aeruginosa, 23,24 and VHb-producing P. aeruginosa grew to higher cell densities when fed with benzene, toluene and xylene. 5 P. putida KT2442 with chromosomally integrated vgb exhibited increased biotransformation of benzene, toluene and chlorobenzene. 25 The modification of P. putida KT2440 with vgb allowed growth on 1,2,3-trichloropropane under oxygen-limited conditions, a condition in which the vgb-free control was unable to grow. 6 Applications of VHb engineering in Pseudomonas aiming at PHA production are scarce. P. putida KT2442 with the pha operon partially replaced by the vgb operon exhibited an enhancement of PHB-co-mcl-PHA productivity of up to 35% that was associated with vgb expression, although the cellular production in both the vgb-positive strain and the vgb-negative control strains were inferior to half of that of the parent KT2442 strain. 26
The present study is based on the hypothesis that vgb expression in P. putida KT2440 can be beneficial to mcl-PHA production by increasing cell growth and polymer production or by delaying the onset of oxygen limitation. This hypothesis was tested in carbon-limited, high-cell-density, fed-batch fermentations, which have been shown to achieve high mcl-PHA productivities, although requiring costly oxygen supplementation. Wild-type P. putida KT2440 and recombinant constructs harboring 0, 1 and 2 copies of the vgb gene were evaluated as mcl-PHA production strains.
Materials and methods
BACTERIAL STRAINS AND PLASMIDS
The bacterial strains and plasmids used in this study are listed in Table 1. 8,27 –29 Escherichia coli DH5αF’ was used for routine cloning and Pseudomonas putida KT2440 was the mcl-PHA producer which was genetically modified with the vgb gene. E. coli DH5αF’ was grown in Luria-Bertani (LB) medium at 37°C and P. putida KT2440 was grown in LB medium or a defined mineral salts medium at 30°C. Kanamycin sulfate (100 μg mL−1) was added as required. pGEM®-T Easy was used to subclone and confirm the sequence of the vgb gene. pBBR1MCS-2 was used as the expression plasmid for the vgb gene in P. putida under control of the lac promoter.
Strains and Plasmids Used in This Study
ATCC, American type culture collection.
CONSTRUCTION OF RECOMBINANT P. PUTIDA STRAINS HARBORING PLASMID PMCS2-VGB1 OR PMCS2-VGB2
All genetic techniques followed the procedures as described by Sambrook and David.
30
The DNA fragment was sequenced using the chain termination method and an automatic sequencer (Genome Quebec Innovation Centre, Montreal, Canada). A 508-bp DNA fragment containing the vgb gene with a ribosome binding site was amplified by PCR employing a pair of primers with anchored Kpn I restriction sites which are underlined. The primer sequences were derived from the Vitreoscilla sp. C1 hemoglobin gene deposited in the GenBank database under the accession number L21670.1, vgK-F2: 5′-ATT
The vgb gene was amplified from pGEM-vgb by PCR, digested with Kpn I and ligated into Kpn I-hydrolyzed pBBR1MCS-2, resulting in two plasmids pMCS2-vgb1 and pMCS2-vgb2 (Fig. 1). The two plasmids were distinguished by molecular weight after verifying with Pst I and analyzed on agarose gel. The transcriptional direction of the vgb gene(s) under control of the lac promoter in pMCS2-vgb1 and pMCS2-vgb2 was verified by digesting both plasmids with the restriction enzyme Mls I and analyzed on agarose gel.

Schematic representation of the construction of the plasmids pMCS2-vgb1 and pMCS2-vgb2. Italic numbers in parentheses indicate the site of respective restriction enzymes on plasmids.
GROWTH MEDIA
The inoculum for the batch fermentations was grown in two 500-mL Erlenmeyer flasks containing 150 mL of medium, inoculated from cultures kept in glycerol at −80°C. The flasks were maintained in an orbital shaker at 30 ± 1°C and 200 rpm for approximately 12 h before inoculation. The inoculum medium contained (per L): 6.36 g Na2HPO4, 2.70 g KH2PO4, 4.70 g (NH4)2SO4, 0.80 g MgSO4·7H2O, 3.00 g nutrient broth and 9.00 g glucose. The initial fermentation medium contained (per L): 6.36 g Na2HPO4, 2.70 g KH2PO4, 4.70 g (NH4)2SO4, 0.80 g MgSO4·7H2O and 10 mL of trace element solution. The trace element solution was formulated according to Sun et al. 31 Kanamycin was added to the fermentation medium for the recombinant strains at a concentration of 100 mg L−1.
BIOREACTOR CONDITIONS AND CONTROL
The fed-batch fermentations were performed in a Minifors stirred tank bioreactor (Infors HT, Bottmingen, Switzerland) with a 1.6-L working volume. The agitation was maintained at 700 rpm and the temperature was controlled at 30 ± 1°C. A 14% (w/v) NH4OH solution was used to maintain the pH at 6.85 and also served as a nitrogen source. The dissolved oxygen concentration was measured with an Ingold polarographic probe. The aeration rate was maintained at 1 L min−1 through the use of a mass flow controller. The oxygen and carbon dioxide contents of the inflow and off-gas streams were continuously monitored with a Tandem PRO Gas Analyzer (Magellan Instruments, Borehamwood, United Kingdom). The reactor feeding was controlled based on the mass of the nonanoic acid reservoir as described previously. 31 An initial 5-h batch phase was conducted before starting the feeding phase. Carbon-limited growth was achieved by exponentially feeding nonanoic acid to support a specific growth rate of 0.15 h−1. Each set of experimental conditions was performed at least twice in independent experiments. Data acquisition and reactor control were performed with LabVIEW™ 6.1 (National Instruments, Austin, TX).
ANALYTICAL METHODS
Biomass concentration was measured gravimetrically as cell dry mass (CDM). Known volumes of broth samples were centrifuged at 9,000 × g for 15 min, washed once with distilled water and lyophilized. Residual biomass (Xr) was calculated as the difference between the total CDM and the measured PHA concentration. The sample supernatants were analyzed for phosphate concentration by the molybdenum blue method; 32 for ammonium concentration by the phenol-hypochlorite method; 33 and for residual nonanoic acid concentration by GC-FID analysis according to Ramsay et al. 34 The intracellular PHA content was determined by GC-FID after sample methanolysis using benzoic acid as an internal standard, according to Braunegg et al. 35 External standards for mcl-PHA quantitation were prepared by repeated solvent extraction and precipitation in cold methanol. 36 The monomeric composition of the standards was determined by 1 H and 13 C nuclear magnetic resonance spectroscopy.
The production of VHb by the recombinant strains was measured as heme protein by fluorescence analysis. The iron present in the heme group was removed with an oxalic acid solution, resulting in fluorescent protoporphyrin according to the method described by Sinclair et al. 37 The specific oxygen uptake rate was evaluated throughout the fermentations using the inflow and off-gas oxygen contents and assumed a pseudo-steady state condition.
Results and Discussion
CONSTRUCTION OF RECOMBINANT P. PUTIDA STRAINS HARBORING PLASMID PMCS2-VGB1 OR PMCS2-VGB2
Recombinant plasmids pMCS2-vgb1 containing one copy of vgb gene and pMCS2-vgb2 containing two copies of vgb genes were constructed by ligating the Kpn I-treated vgb gene with the Kpn I-treated pBBR1MCS-2 plasmid. The reaction of ligation was transformed into E. coli DH5αF’ strain, then spread on solid LB medium containing kanamycin sulfate (100 μg mL−1) and X-gal. Candidate plasmids in fifty-eight white colonies appearing on the solid LB medium were extracted and pre-analyzed for their molecular weight on agarose gel by comparing to the mobility of pBBR1MCS-2 plasmid as control. Successful ligations would create recombinant plasmids with higher molecular weights (5658 bp and 6172 bp), while unsuccessful ligations would create self-circled plasmids with molecular weight being equal to control (5144 bp) (data not shown). Consequently, seventeen successful recombinant plasmids were selected for further verification by digesting with the Pst I restriction enzyme and analyzed on agarose gel.
After the Pst I reaction, recombinant plasmids containing one and two copy(ies) of vgb gene(s) would give DNA fragments of 3902 bp and 1756 bp, and of 3902 bp and 2270 bp, respectively; while the control plasmid pBBR1MCS-2 would give DNA fragments of 3902 bp and 1242 bp (Fig. 2A). As a result, twelve constructed plasmids were chosen as plasmids containing one copy of vgb gene (lanes 4–7, 9, 11, 13–18) and four constructed plasmids were selected as plasmids containing two copies of vgb genes (lanes 3, 8, 10, and 19).
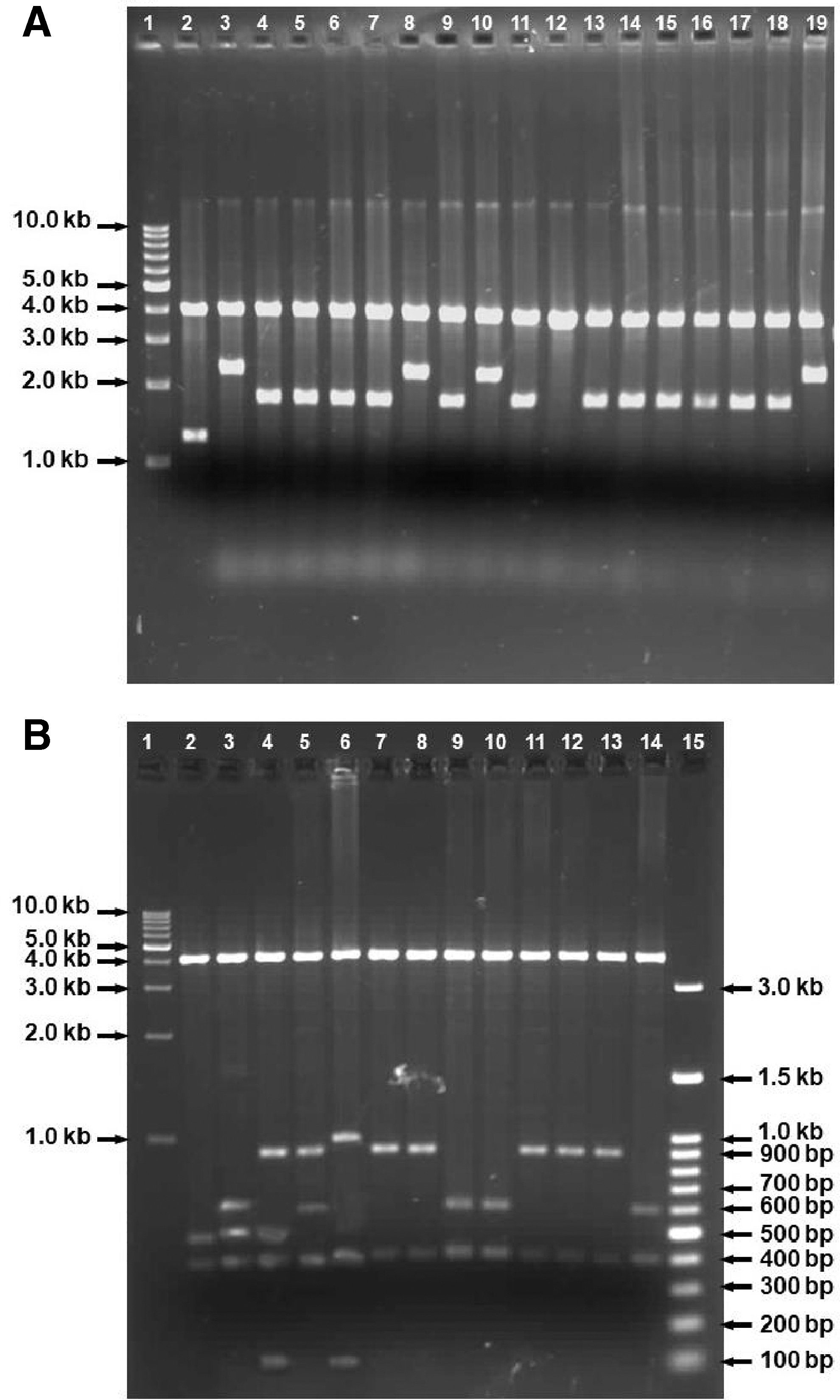
Agarose gel analysis of reactions catalyzed by restriction enzymes
Subsequently, the transcriptional direction of the vgb genes under control of the lac promoter on constructed plasmids was verified by performing reactions catalyzed by the restriction enzyme Mls I and analyzed on agarose gel. In plasmids containing two copies of the vgb gene, when both copies have the same transcriptional direction as the lac promoter, one would expect DNA fragments of 4248, 599, 514, 410, and 401 bp. When one or both copies are in the opposite transcriptional direction as the lac promoter, one would expect DNA fragments of 4248, 906, 514, 401 and 103 bp; or 4248, 906, 599, 401 and 18 bp; or 4248, 1010, 410, 401 and 103 bp. Plasmids containing one copy of the vgb gene in same transcriptional direction as the lac promoter would give DNA fragments of 4248, 599, 410, and 401 bp; while copies in the opposite transcriptional direction as the lac promoter would give DNA fragments of 4248, 906, 401, and 103 bp. The control plasmid pBBR1MCS-2 would give DNA fragments of 4248, 495, and 401 bp (Fig. 2B). As expected, one desired plasmid containing two copies of the vgb gene was selected (lane 3) and designated as recombinant plasmid pMCS2-vgb2. Three desired plasmids containing one copy of vgb gene were selected (lanes 9, 10, and 14) and designated as recombinant plasmid pMCS2-vgb1. Plasmids pMCS2-vgb1 and pMCS2-vgb2 were then transformed into P. putida KT2440 to create the recombinant P. putida strains Vgb-1 and Vgb-2, respectively.
Absence of repression of the lac promoter was confirmed by culturing the recombinant cells with and without addition of the inducer isopropylthiogalactopyranoside (IPTG, 100 mmol L−1). The vgb gene was expressed in both conditions, indicating that both recombinant constructs exhibited constitutive vgb expression.
FED-BATCH FERMENTATIONS WITH P. PUTIDA STRAINS
Carbon-limited, fed-batch fermentations were performed with wild-type P. putida KT2440 and the recombinant strains Vgb-1, Vgb-2 and a control strain harboring the expression plasmid but no vgb gene (Vgb-0). For all of the strains investigated, the fermentation was terminated after 25 h of feeding due to oxygen limitation which caused the cessation of growth, as marked by a sudden drop in the carbon dioxide production rate. Similar biomass profiles were obtained for all strains, and the data from replicate fermentations was highly reproducible (Fig. 3). The wild-type produced slightly more biomass, achieving 27 g CDM L−1 by the end of the experiment, compared to 24, 23 and 21 g CDM L−1 for Vgb-0, Vgb-1 and Vgb-2, respectively. The intracellular PHA content increased constantly during each fermentation. The wild-type produced the most PHA, 61% of CDM at 25 h. The Vgb-2 strain achieved a maximum PHA content of 42%, noticeably lower than the 54% and 55% observed for the Vgb-1 and Vgb-0 strains, respectively. The PHA monomer composition was independent of the production strain. 3-hydroxyheptanoate (C7) and 3-hydroxynonanoate (C9) were the only monomers detected. An average monomer composition of 44:56 (C7:C9, %mol) was observed for all strains.

Biomass production, intracellular PHA content, dissolved oxygen and specific oxygen uptake (qO2) rate profiles obtained during carbon-limited, fed-batch fermentations with exponential feeding of nonanoic acid using the recombinant strains Vgb-0 (), Vgb-1 ( ), Vgb-2 (
), Vgb-2 ( ) and the wild type (WT,
) and the wild type (WT,  ) strain P. putida KT2440. Error bars represent one standard deviation from the mean of two independent fermentations. Color images are available online.
) strain P. putida KT2440. Error bars represent one standard deviation from the mean of two independent fermentations. Color images are available online.
A marked decrease in the dissolved oxygen profile occurred concomitant with a more intense biomass activity, between 13 and 25 h in all fermentations. The specific oxygen uptake rate (qO2) profiles were nearly indistinguishable between the three recombinant strains, presenting little variation during the experiment, with an average value of 3.9 mmol gXr −1 h−1.
The behavior of the wild-type P. putida KT2440 strain in carbon-limited, fed-batch fermentations with nonanoic acid exponential feeding has been well characterized. 31 The total biomass yield on nonanoic acid (YX/NA) and the PHA yield on the substrate (YPHA/NA) obtained in the present study (Table 2) were comparable to those reported by Sun et al., 31 who obtained a YX/NA of 0.83 g g−1 and a YPHA/NA of 0.60 g g−1 with the same specific growth rate of 0.15 h−1. The biomass and PHA production performance parameters obtained in the present study, however, were not comparable to those reported by Sun et al., 31 mainly due to the use of pure oxygen supplementation in that study.
Comparison of Carbon-Limited, Fed-Batch Fermentations for mcl-PHA Production from Nonanoic Acid at a Specific Growth Rate of 0.15 h−1
Pure oxygen supplementation was employed in that study.
The recombinant strains did not present observable advantages in any of the evaluated parameters. The performance of the recombinant strains in PHA production was noticeably inferior to that of the wild type. The control strain Vgb-0 presented an 11% reduction in biomass production and a 10% reduction in PHA accumulation when compared to the wild-type strain. While the Vgb-0 and Vgb-1 strains had similar trends, a marked difference was observed between the Vgb-0 and Vgb-2 strains. The Vgb-2 strain exhibited a 12% decrease in biomass production, a 24% decrease in PHA accumulation and a 39% decrease in PHA yield.
The VHb levels were measured during the fed-batch fermentations with the strains Vgb-1 and Vgb-2 (Fig. 4). Increasing the number of vgb genes in the expression plasmid did not result in higher VHb production, as similar temporal profiles were observed for both strains. A maximum level of 12 nmol g−1 of dry residual biomass was observed for the Vgb-1 strain.
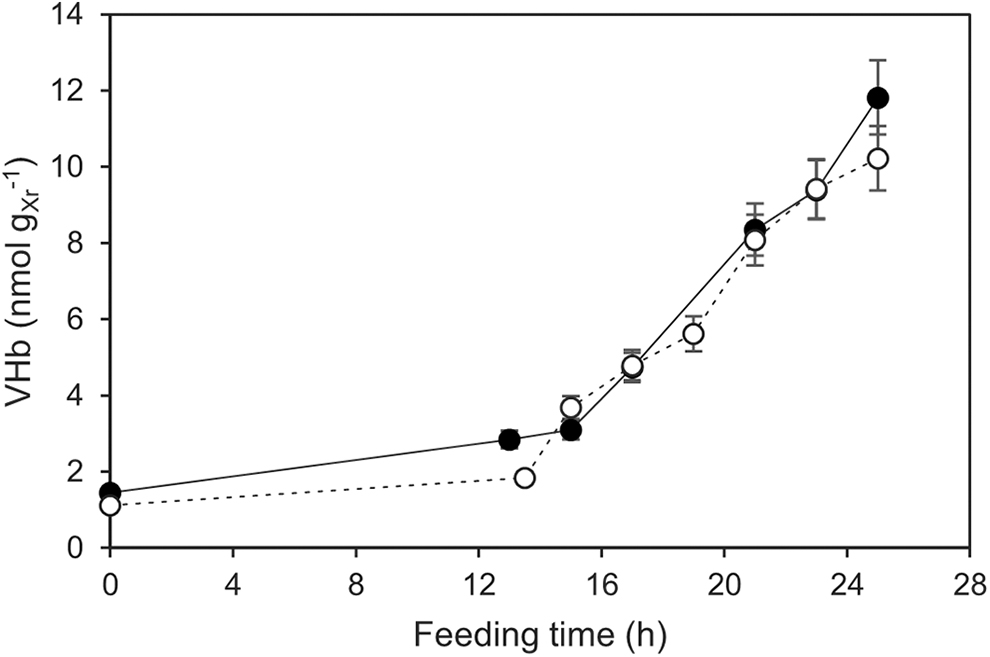
Specific VHb production throughout the fermentations with the recombinant strains (•) Vgb-1 and (○) Vgb-2. Error bars represent one standard deviation from the mean of replicate experiments.
Conclusion
The VHb-producing Pseudomonas strains evaluated in this study did not present enhanced growth or PHA production when compared to the wild-type strain. In fact, PHA synthesis in the three recombinant strains was inferior to that of the unmodified strain, even when no vgb gene was present in the expression plasmid. This suggests a metabolic burden associated with the expression plasmid, and indeed the kanamycin resistance gene in this plasmid has been shown to impede P. putida growth. 38 All fermentations were terminated at similar times due to insufficient oxygen supply. Oxygen limitation remained the limiting step to achieving higher cellular densities, showing that VHb production did not alleviate its effects.
The physiological effects of VHb have been shown to occur in a dose-dependent manner in recombinant E. coli. 39 The VHb levels observed in the present work, achieving a maximum of 12 nmol g−1 of dry residual biomass, were substantially less than those reported for the native producer Vitreoscilla, which may achieve 50 nmol g−1 wet cell weight. 40 In recombinant E. coli, VHb levels are often in the 100 nmol g−1 wet cell weight range under inducing conditions. 19,39,41,42 However, VHb levels in pseudomonads are often below 15 nmol g−1 wet cell weight, 21 –24,42 and those levels were enough to elicit observable positive effects. Most of the earlier works with VHb engineering in pseudomonads had the vgb gene placed under the native vgb promoter, 21 –24,42 producing similar levels of expression of VHb to the lac promoter in the present study.
The expression of vgb did not influence oxygen usage by the recombinant strains, as comparable qO2 values were maintained during the fermentations, independently of the production of VHb. Contrary to our findings, VHb-producing strains have been reported to result in higher qO2 when compared to VHb-free controls. This behavior has been observed in the facultative aerobic bacteria Enterobacter aerogenes 43 and E. coli, 39,42,44 –46 as well as in the obligate aerobic bacterium Pseudomonas aeruginosa 42 and the aerobic yeast Pichia pastoris. 7,47 Surprisingly, there is little discussion in the literature regarding the implications of an increased qO2 in vgb-harboring strains. An increase in qO2, the microbial oxygen demand, means that the maximum oxygen transfer capacity of the bioreactor will be reached at an even lower biomass concentration. Therefore, an increase in oxygen demand by vgb-harboring cells would constitute an obstacle for achieving high-cell-density fermentations. In most cases, maximum productivity depends on the bioreactor's oxygen transfer capacity, which is unaffected by VHb content.
The actual mode of action of VHb remains largely unknown. Measurements of the oxygen-binding properties of VHb revealed an average affinity constant for oxygen and relatively slow rate of oxygen release, 48 indicating that it is likely not involved in oxygen transport. It has been proposed that, in E. coli, VHb increases the overall efficiency of aerobic metabolism, without affecting the kinetics of oxygen consumption. 49 Oxygenated VHb would influence several oxygen-regulated metabolic processes within the cell, most notably the expression of the respiratory terminal oxidases. This modulation would enhance the overall proton translocation efficiency of the cell, generating more ATP per molecule of reduced oxygen. 50 VHb also influences the oxygen-regulated switching of different metabolic modes in E. coli. Under microaerophilic conditions, VHb-producing strains are capable of maintaining fully aerobic metabolism and delay the onset of the less efficient fermentative pathways. 51
Considering the markedly distinct metabolic and regulatory backgrounds between pseudomonads and E. coli, it is difficult to establish whether the same physiological effects of VHb observed in E. coli are expected to occur in pseudomonads. Although some analogous oxygen-dependent regulatory systems are present in P. putida, 52 the potential effect of VHb on these systems remains to be elucidated.
Similar to our results, VHb engineering did not confer desirable advantages to the heterologous hosts Enterobacter aerogenes, 53 Serratia marcescens, 54 E. coli, 42 as well as in several species of higher plants (reviewed by Stark et al.). 55 These results demonstrate that VHb engineering is not universally advantageous and its utility needs to be assessed on a case-by-case basis.
Footnotes
Funding Information
The authors gratefully acknowledge the scholarship granted to Dr. Andréa Lima dos Santos Schneider by the Coordination for the Improvement of Higher Education Personnel (CAPES) and the support provided by the São Paulo Research Foundation (FAPESP, grant 2016/26034-7).
Author Disclosure Statement
No competing financial interests exist.