
Abstract
To meet increased demand for biofuels, biorefineries need to achieve higher yields, which could be achieved by optimizing current processes by using thermotolerant microorganisms in first- and second-generation (2G) biofuel production. 2G biofuel production is comprised of two basic steps before fermentation of the reducing sugars: lignocellulosic biomass pretreament and enzymatic hydrolysis, which is typically performed between 40–55°C. However, Saccharomyces cerevisiae, the most commonly used microorganism, performs fermentation at temperatures up to 35°C, which reduces the enzymatic activity when this yeast is combined in processes such as simultaneous saccharification and fermentation (SSF) and consolidated bioprocessing (CBP). In this context, the use of thermotolerant yeast isolates in SSF and CBP could promote enzymatic hydrolysis at temperatures closer to the optimal of enzymes, increasing the sugars and biofuel yields. Moreover, increased temperatures will reduce cooling costs and contamination by other microorganisms that compete with S. cerevisiae for the released sugars. Therefore, we used adaptive laboratory evolution to evolve two industrial Brazilian S. cerevisiae strains, PE-2 and SA-1, for thermotolerance. Compared to their parental strains, AMY35 (SA-1) and AMY12 (PE-2) had 63% and 61% higher cell growth at 40°C, respectively, and showed an ethanol yield of ∼0.44 g ethanol/g glucose. In addition, the reduced trehalose content in the evolved isolates compared with the respective parental strains at 40°C suggests that other mechanisms are responsible for the thermotolerance. Results showed that our adaptive evolution approach was sufficient to evolve diploid industrial S. cerevisiae strains and that the selected strains had great potential for high-temperature fermentation.
Introduction
There is a global consensus that biofuels can partially or totally substitute fossil fuels, and bioethanol holds great promise of becoming a commodity biofuel. 1 However, to meet worldwide demand, biorefineries need to achieve higher yields and productivities. 2 As research advances and new fermentation processes emerge, the need for robust microorganisms that perform well under stressful conditions becomes evident. Although several bacteria and yeasts are capable of converting fermentable sugars into ethanol, S. cerevisiae is the most widely used in industrial applications due to the ability to produce and tolerate high ethanol concentrations and resist inhibitors from pretreated biomass 3 such as phenolic compounds (e.g., ferulic acid) and furans (e.g., furfural). Although S. cerevisiae usually performs fermentation at 25–35°C in industrial processes, 4 higher temperatures reduce medium viscosity and oxygen solubility, facilitating agitation and the maintenance of anaerobic conditions during fermentation, 5 preventing microbial contamination, and reducing cooling costs by decreasing water and energy consumption. 6 The overall costs of ethanol production from starch could be reduced by US$30,000 per year for a 30,000 kL plant if fermentation temperature is increased 5°C, which could also benefit sustainable initiatives that use lignocellulosic biomass, such as 2G biofuel production. 6
2G biofuels production is comprised of initial pretreatment of lignocellulose (e.g., steam explosion, ionic liquids), enzymatic hydrolysis, and fermentation of the released sugars for biofuels production. 7 The enzymatic hydrolysis step could be combined with microorganisms in processes such as simultaneous saccharification and fermentation of hexoses (SSF); simultaneous saccharification and co-fermentation of pentoses and hexoses (SSCF), where the enzyme cocktails are added to the bioreactor; or consolidated bioprocessing (CBP), where the pretreated biomass is mixed with a microorganism capable of producing its own enzymes. In all combinations, higher efficiency would be obtained if microorganisms that could tolerate temperatures closer to the optimal of the enzymes were used, 8 reducing the amount of enzymes loaded and/or the time for process to complete. 9
Temperatures higher than those regularly applied for yeast growth disorganize the cytoplasmic membrane, which results in reduced ethanol yields and tolerance. 10 Several studies search for isolates of thermotolerant yeast strains for ethanol production, including those of Pichia kudriavzevii, Candida tropicalis, 5 S. cerevisiae, 11,12 and Kluyveromyces marxianus. 13 –15 However, some studies found that these isolates showed either lower ethanol yield or low recycling viability when compared to industrial Saccharomyces strains. 16,17
Thermotolerance could be acquired through evolutionary engineering, which harnesses specific phenotypic changes through random mutations under selective pressure, 18 creating new strains with this multigenic trait 19,20 also found in natural 21 and industrial sources. 22,23 However, most studies focused on elucidating these random mutations were carried out in haploid laboratory strains 24 with a genetic background divergent of those well-established industrial strains used often in biorefineries, 25 such as the Pedra-2 (PE-2) 17,26 and Santa Adélia-1 (SA-1), 17,27 isolated from Brazilian sugarcane mills, where the stressful conditions imposed by yeast cell recycling, high cell densities, and ethanol concentrations selected for robust strains with high ethanol yields (92% of theoretical yields). 17
The aim of our work was to improve tolerance towards high temperature of two industrial S. cerevisiae strains, PE-2 and SA-1, highly used in Brazilian sugarcane mills (first-generation ethanol), compare the phenotypic variability emerged when these strains are subjected to the same selective pressure, and evaluate the kinetic and physiological parameters of the selected strains.
Materials and Methods
Strains and Media
Two Brazilian industrial strains of S. cerevisiae were subjected to evolutionary engineering, PE-2 (diploid) and SA-1 (diploid), which were originally isolated from the Pedra Plant and the Santa Adélia Plant,
17
respectively, and stocked in the bank of cultures of the Chemical, Biological and Agricultural Pluridisciplinary Research Center (CPQBA, Brazil). The thermotolerant industrial S. cerevisiae strain AZJ001 (diploid), kindly provided by Dr. Anderson Ferreira da Cunha from Federal University of Minas Gerais (Brazil), was used as a tolerant control, while the
Evolutionary Engineering Approach
Adaptive evolution was conducted separately for each strain according to Alkim, Turanli-Yildiz and Cakar 28 and colony selection was subsequently performed for each of the evolved strains. The parental strains SA-1 and PE-2 were evaluated for growth at 30°C, 38°C and 40°C, 200 rpm for 24 h in Schott glass bottles with 80% volume filled with YPD medium (Table 1). The inoculum cultures were prepared with the parental strains at 200 rpm and 30°C in YPD medium overnight and the appropriate volume recovered, and centrifuged at 5,000 rpm for 3 minutes. The supernatant was removed, washed with sterile distilled water, resuspended in 500 μL sterile distilled water and used to start the cultures to an initial optical density at 600 nm (OD600) of 0.5. Adaptive evolution of the two strains were carried out simultaneously, as previously cited, in Schott glass bottles, and consecutive batch fermentations were grown for 24 h at 38°C and 200 rpm (Fig. 1). At the end of each cycle, a 2-mL sample was removed to measure the OD600 and ethanol concentration, evaluate for contamination under the microscope, and the appropriate volume used to start the subsequent batch fermentation to an initial OD600 of 0.5. The same process was repeated after each cycle of 24 h until the parental SA-1 showed stable growth with OD600 above that of the initial test, 3.64. We cultured the PE-2 strain longer to also obtain stable growth with cell density above that of the initial test at 38°C, however it stabilized with an approximately OD600 of 2.8, and the evolutionary process was continued with both strains at 40°C. The process continued at 40°C as previously cited until the strains showed stable growth, which was reached with an approximately OD600 of 2.5 for both strains.
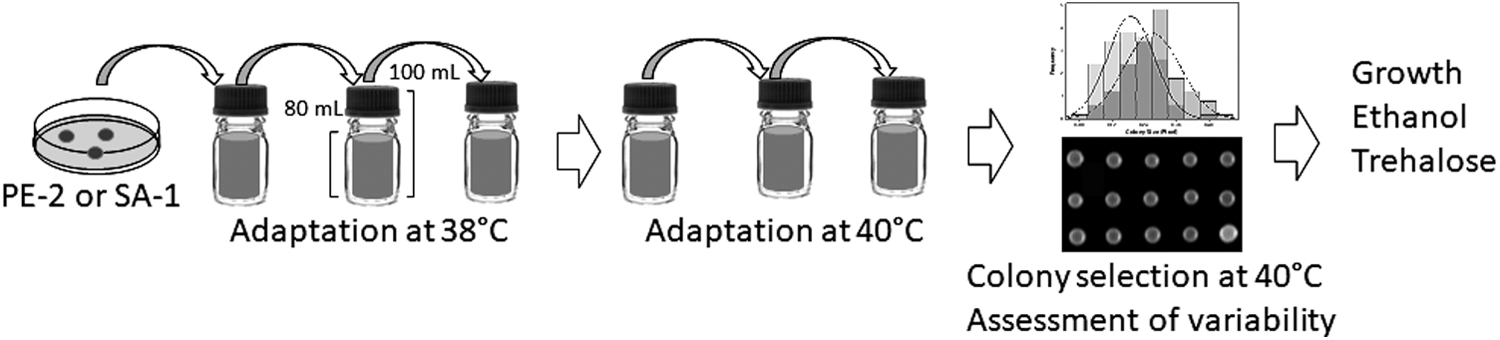
Schematic design of the evolutionary engineering approach used in this study.
Growth Parameters of the Parental Strains PE-2 and SA-1 Cultured for 24 h in YPD Medium
Mean values and standard deviations of three replicates.
We pooled the final population of each strain, washed with distilled water, adjusted to an initial OD600 of 0.1, inoculated into 96-well plates diluted and plated onto 10 petri dishes (150 × 15 mm) plates containing solid YPD medium using a Boekel microplate replicator with 200 colonies per plate, and incubated at 40°C for 24 h. The 83 largest and fast-growing colonies were selected, cultured overnight in YPD liquid medium, washed with distilled water, inoculated into a 96-well plate to an initial OD600 of 0.1, and plated and cultured as previously described at 30°C and 40°C. Twelve randomly distributed wells (to avoid position effects) were inoculated with the controls: 3 wells were filled with YPD medium (blank), 3 with the thermotolerant strain AZJ001, 3 with the susceptible laboratory strain
Fermentations at 30°C and 40°C
We evaluated the growth, fermentation performance and trehalose content of the evolved strains to validate our previous results. Initially, the yeast strains were cultured in 96-well plates containing 200 μL of YPD medium at 200 rpm for 8 h at 30°C and 40°C, and the OD600 was measured using a plate reader (Tecan Infiniti® 200 PRO). The same strains were also cultured in 500-mL shake flasks with 300 mL YPD medium to reduce the rate of oxygen transfer, at 200 rpm for 12 h at two different temperatures, 30°C and 40°C, while the glucose concentrations were 20 or 60 g/L at 40°C. Fermentations with 60 g/L were also performed to evaluate growth and ethanol production under a higher osmotic pressure. Two mL samples were removed to measure OD600 and trehalose content, and the remaining were centrifuged and the supernatant collected to evaluate ethanol concentration. Kinetic parameters and cell dry weight concentrations were calculated according to procedures published elsewhere. 30,31 All fermentations were inoculated to an initial OD600 of 0.2 and performed in duplicates.
Metabolite Concentration
Glucose and ethanol were determined as previously described 32 by high-performance liquid chromatography (HPLC, AccelaTM, Thermo Scientific, Waltham, MA) using a refractive index (IR) detector with an HyperREZTM XP (Thermo Scientific) Organic Acids column (100 × 7.7 mm) at 35°C, with 0.6 mL/min eluent of 5 mM H2SO4.
Determination of Intracellular Trehalose Accumulation
Intracellular trehalose was determined using the modified anthrone method previously described. 33 Five mL cultures of each strain were inoculated to an initial OD600 of 0.1 in YPD liquid medium at 200 rpm and 30°C or 40°C overnight (∼10 hours), collected in the late exponential phase, the OD600 measured, cell quantity of each strain adjusted, centrifuged at 5,000 rpm for 3 min, the cells washed in sterile distilled water, and the pellet resuspended in 400 μL of 0.5 M trichloroacetic acid and mixed at room temperature for 40 min in an ultrasonic bath. The supernatant was separated by centrifugation at 18,000 × g for 2 min, and an aliquot of 200 μL were mixed with 1 mL of cold anthrone reagent (0.005% anthrone and 1% thiourea in 66% (v/v) H2SO4), incubated for 10 min at 100°C, cooled immediately in water, and the absorbance was measured at 620 nm in a spectrophotometer (Genesys 20, Thermo Scientific).
Results and Discussion
Long-Term Evolutionary Engineering
Evolutionary engineering of the two strains was carried out with different periods—SA-1 for 158 days and PE-2 for 183 days (Fig. 2)—due to often bacterial contaminations and unstable growth in the latter. The SA-1 strain was stable in the final quarter of the evolution at 38°C, which showed several overnight fermentations with cell concentration higher than those of the parental strain at 38°C and 30°C. The increased stability observed in the growth of the SA-1 population demonstrates that this strain could generate stable variants adapted to thermotolerance faster or that once individuals with this phenotype emerged in the population, they had specific growth rates higher than those arising in the PE-2. It could occur due to genetic mechanisms that foster this phenotypic variability found in this strain that may be lacking in the PE-2. After the first cycle of selective pressure at 38°C, the strains were cultured at 40°C to further increase thermotolerance, with the SA-1 batch fermentations closer to the parental optimal at 30°C and one fermentation with cell density above it, while the PE-2 had all overnight batches under its parental growth at 30°C.
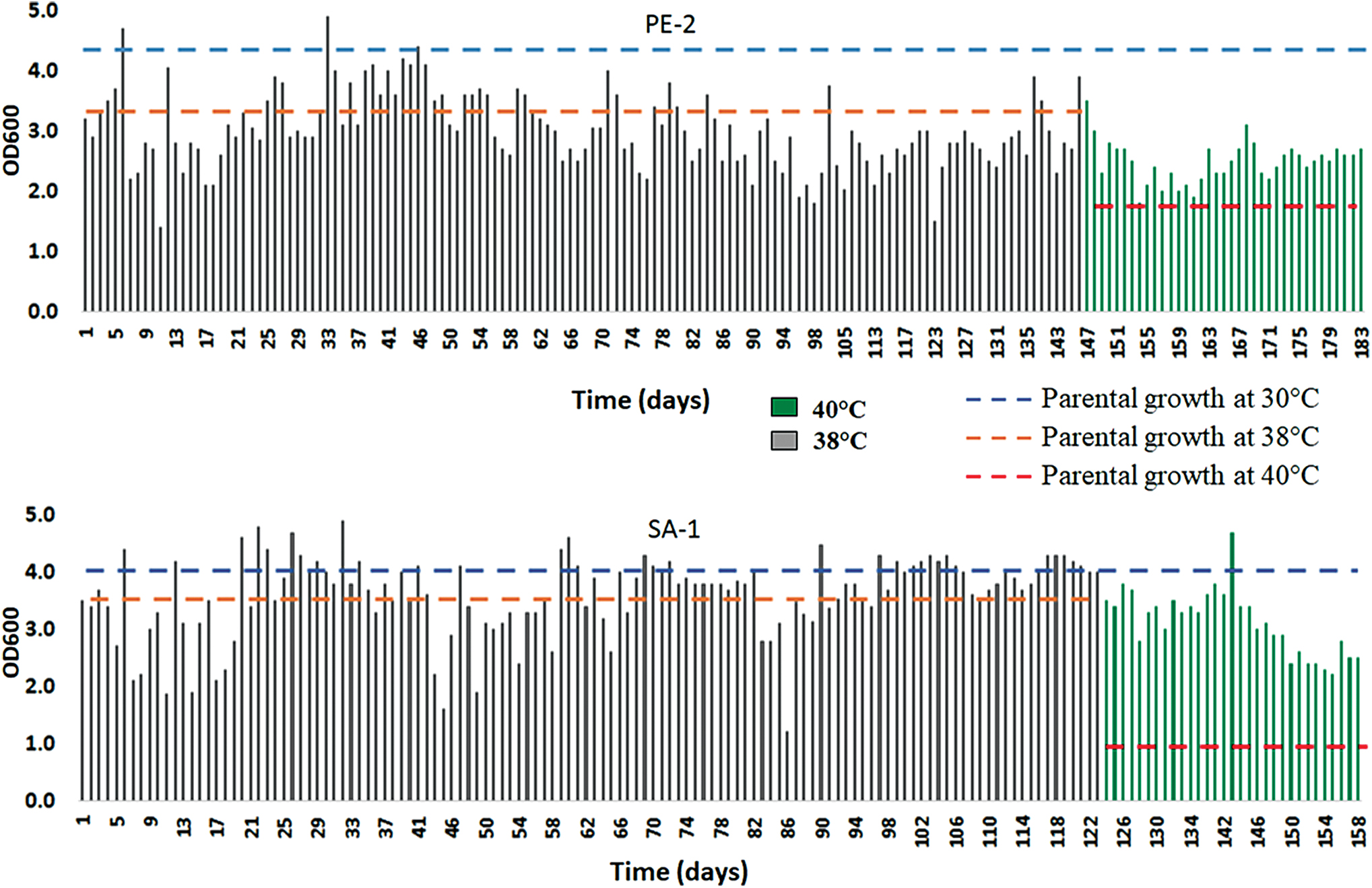
Follow-up of the evolution profile of the parental strains PE-2 and SA-1 at 38°C and 40°C. Color images are available online.
Selection of Thermotolerant Isolates
In the first step of colony selection for thermotolerance, the last batch of evolution at 40°C of the two populations were plated onto solid YPD media and cultured at 40°C for 24 h. Two thousand colonies of each strain were semi-quantitatively evaluated for colony formation, and the 83 best-performing colonies were quantitatively reevaluated and compared with controls using ImageJ (Fig. 3). 29,34 The strains showed a normal distribution of colony size, i.e., most colonies exhibited intermediate growth, with lower percentages showing poor or excellent growth (Fig. 4). SA-1 colonies showed, as expected, improved growth and variability with the mean colony size and standard deviation 25% and 20% higher, respectively, compared to the PE-2 population. These results followed those in the evolutionary engineering in the glass bottles and indicates that, although the SA-1 parental had an OD600 lower than the PE-2 parental at 40°C (Table 1), it could cope faster with higher temperature, generating more variants and higher frequency of colonies with an improved growth capacity at 40°C.
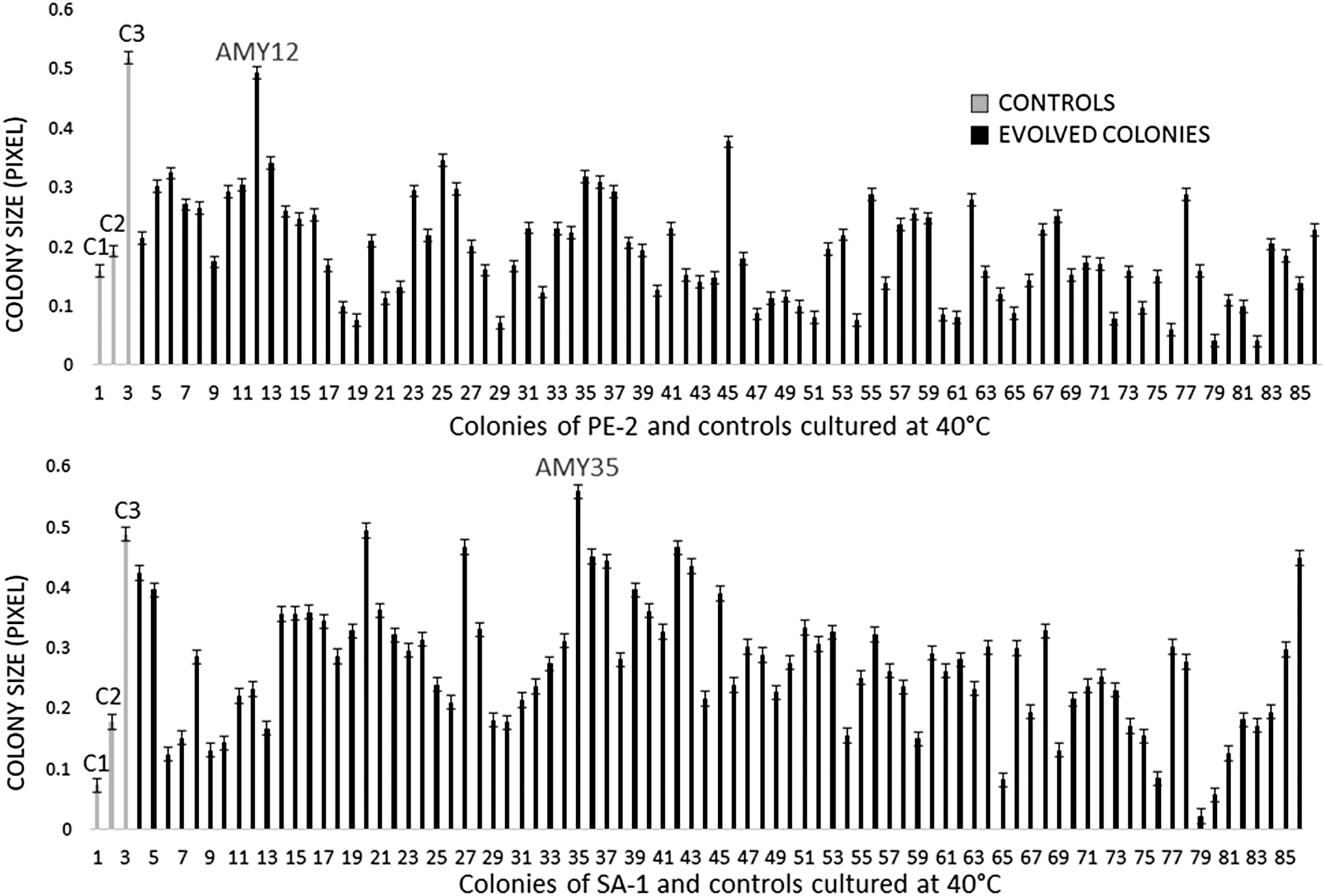
Screening of thermotolerant colonies plated onto solid YPD medium and cultured at 40°C. C1: Susceptible strain CEN.PK122, C2: Parental PE-2 or SA-1, C3: Tolerant strain AZJ001.
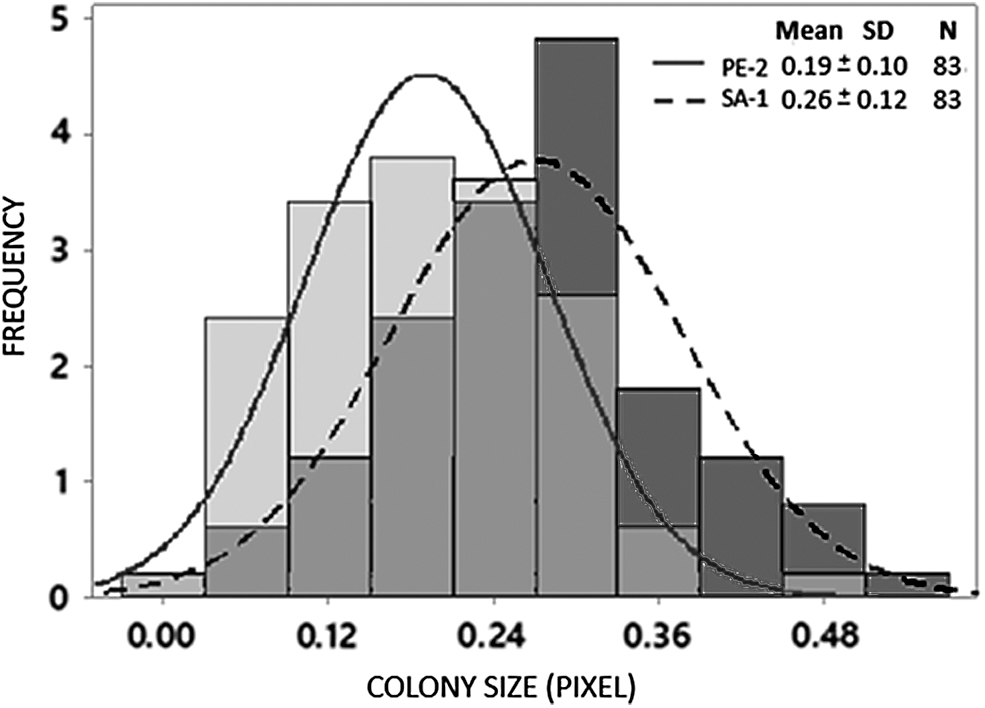
Frequency distribution of colony sizes of the evolved strains. The two means are significantly different with p < 0.0001 according to Student's t-test.
Three colonies of the SA-1 population, 20, 27 and 42 (Fig. 3), grew equally to the tolerant control AZJ001, while the colony 35 (AMY35), was 13% and 63% larger than the AZJ001 and the parental strain, respectively. Although the mean colony size of the PE-2 population was smaller than that of the SA-1, the colony 12 (AMY12) grew equally to the AZJ001 and was 61% larger than the respective parental strain, thus we choose it for further evaluations.
Evaluation of the Adapted Strains AMY35 and AMY12
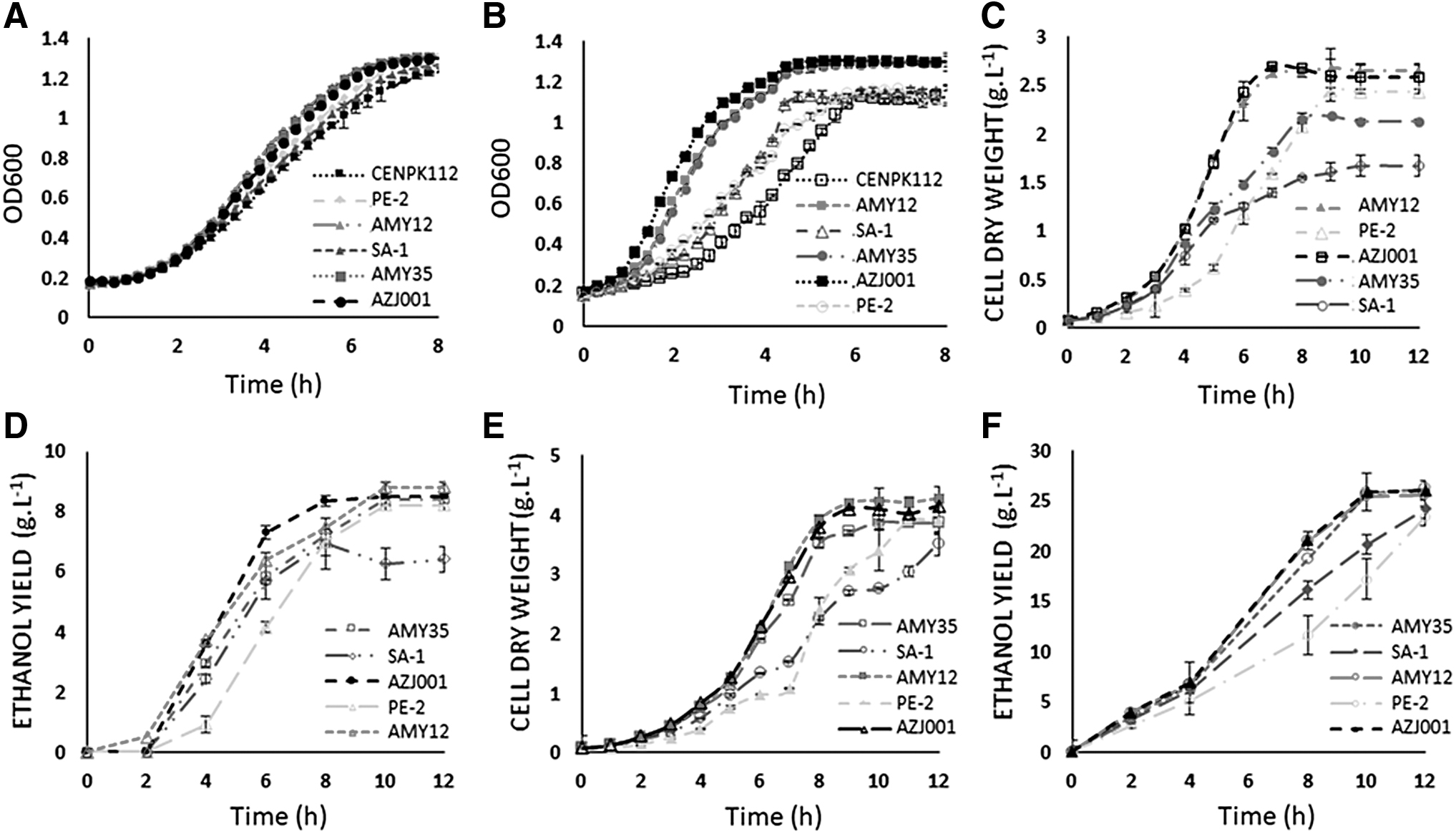
The small-scale fermentation in the plate reader showed that the evolved and AZJ001 strains had the highest growth compared to the parental and
Growth curves and ethanol yield at 30°C (20 g/L glucose) and 40°C (20 g/L and 60 g/L glucose);
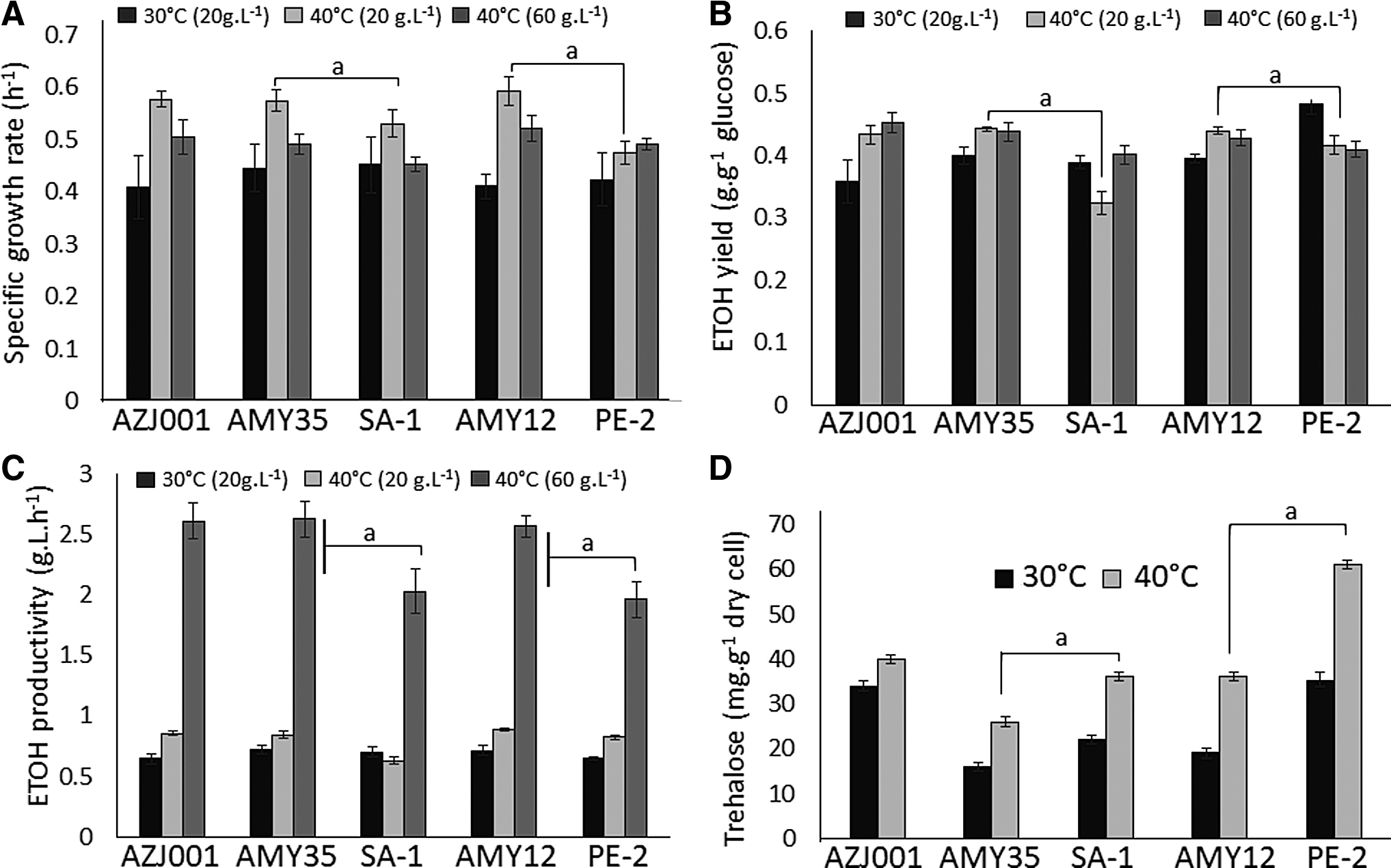
Kinetic parameters of growth and ethanol production after 12 h, and trehalose content at 30°C (20 g/L glucose) and 40°C (20 g/L and 60 g/L glucose);
The evolved and AZJ001 strains also showed a significant increase in ethanol productivity compared to the parental strains in the fermentation with 60 g/L of glucose, which could also indicate that the evolved strains tolerate higher ethanol concentrations, in addition to ethanol production at higher temperatures. Ethanol productivity at 40°C (60 g/L) in our conditions increased approximately 24% and 31% when using AMY35 and AMY12 compared to the respective parental strains, suggesting that our strains could also increase ethanol production in bioprocesses such as SSF, SSCF and CBP, considering the increased release of sugars by the enzymes working at this temperature and the improved phenotype for fermentation showed by our strains. Results supporting our hypothesis include Suryawati et al. 9 and Hu et al. 38 , who used K. marxianus at 45°C and S. cerevisiae at 38°C, respectively, to increase ethanol yield and productivity in SSF.
Other works used more complex evolution processes, such as genome shuffling and UV-mutagenesis to evolve S. cerevisiae, with the maximal ethanol yields also obtained at 40°C, 39,40 including the cell viability at higher temperatures. 40 However, our work obtained significant improvements in ethanol production with a simpler approach in two industrial strains used in Brazilian ethanol industry.
Trehalose Accumulation in Response to Thermal Stress
Accumulation of intracellular trehalose is one of the response mechanisms to thermal stress 41 -43 and strongly correlates with heat resistance in yeasts. 44 However, Nwaka et al. 45 suggested that trehalose accumulation in S. cerevisiae is an initial response to thermal stress and that better adapted individuals have other response mechanisms. The trehalose content was higher in both parental strains compared to the respective evolved strains at 30°C and 40°C (Fig. 6D), with an accumulation around 2-fold more trehalose when cultured at 40°C. This suggests that thermal stress could also trigger trehalose accumulation in these strains, which has a role in protecting cells against heat by activating the environmental stress response. 46 Although the trehalose content was also higher in the evolved strains at 40°C, compared to growth at 30°C, the increased accumulation in the parental strains is contrary to results found by Benjaphokee et al., 33 which suggests that our evolved strains could have genomic mechanisms that emerged for additional protection reducing the trehalose content and the need for this disaccharide. Omics technologies (genomics, transcriptomics, proteomics and metabolomics) applied to these strains could provide more clarity.
Conclusion
The evolutionary engineering approach used in this study was able to select two thermotolerant individuals, AMY35 and AMY12, from robust industrial strains highly used in the first-generation biofuel industry. These strains were able to grow faster and produce ethanol with higher productivity at 40°C with increased glucose concentration (60 g/L), showing that product release increases with higher amounts of substrate compared to the parental strains, which had an ethanol production rate ∼23% lower at equal conditions. The higher phenotypic variability and colony sizes found in the SA-1 population at 40°C indicates that this strain could have genetic mechanisms to generate diversity faster when adaptive evolution is performed for thermotolerance and possibly to other phenotypes such as tolerance to inhibitors present in pretreated lignocellulose. These results indicate that these strains are excellent chassis for ethanol production when using high-temperature processes such as those performed in tropical countries. Although our isolates were obtained using a simple methodology, it could have been generated faster if genome shuffling or mutagenic (UV, EMS) approaches were used. Also, phenotypes such as resistance to ethanol and inhibitors found in the pretreated biomass could be pursued in future works with these isolates. The AMY12 and AMY35 isolates are interesting examples for whole-genome sequencing to analyze markers associated to thermotolerance and ethanol productivity. In this context, our group is working on the heterologous expression of cellulases in both strains to perform CBP, and also in the whole-genome sequencing of a pool of thermotolerant isolates from the PE-2 parental.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported in part by FAPESP [Grant N° 04602-3/2016; 20630-4/2015] and by CAPES finance code 001.