
Abstract
In the food industry, the β-galactosidase enzyme is widely used, being applied mainly in the elaboration of products for the lactose intolerant, can carry out the hydrolysis of lactose. To date, it has been immobilized by different methods to expand and improve its application. In this work, β-galactosidase was produced from yeast Kluyveromyces marxianus and immobilized on Duolite® A-568 ion exchange resin. Optimization of the immobilization conditions was performed using Central Composite Rotational Design with the variables: pH, buffer concentration, and activity offered in immobilization. Immobilization was also evaluated for commercial Lactozyme® enzyme. The produced enzyme showed thermal stability at temperatures between 30 and 35°C, pH 7.3, and storage stability under refrigeration (8°C) for 7 weeks. The best results for the immobilization optimization for β-galactosidase produced (activity recovered in the range of 19.24%) were obtained for pH values between 6.8 and 7.3, buffer concentration between 0.1 and 0.8 M, and activity offered between 18.5 and 27 U. Results, when compared to the use of a commercial enzyme, that reached recovered immobilization of 12.22% in pH 6.0 and 19.47% in pH 7.0 shows, that enzyme β-galactosidase immobilization in Duolite A-568 resin is a possibility.
Introduction
The enzyme β-galactosidase is classified as a hydrolase and it catalyzes lactose hydrolysis to form galactose and glucose. This is of significance in the food industry and has been much studied in terms of biosynthesis and application processes. Its main application is in the manufacture of low lactose products, making dairy products suitable for consumption by lactose-intolerant people. Another application that has become of great interest is the application of such enzymes in the synthesis of galactooligosaccharides (GOS), which are considered prebiotics that help in the maintenance of intestinal flora. 1 –3 β-galactosidases are widely distributed in nature and can be found in plants, animals, and microorganisms such as bacteria, filamentous fungi, and yeast. However, production by microorganisms is most common. 3,4
The yeast Kluyveromyces marxianus has been used in the production of β-galactosidase and also in ethanol production. 5 –7 In addition to having rapid growth, it has great potential for biotechnology application, has Generally Recognized as Safe (GRAS) status, is considered safe, and is capable of assimilating lactose and inulin, which it uses as its carbon source. Currently, there is a particular interest in lactose as a source of carbon to increase the use of whey and whey permeate, because they have high concentrations of lactose and are considered by-products of the dairy industry, consequently an environmental problem. 8 –10
The use of free enzymes in industrial processes is difficult due to instability, high cost, and difficult recovery at the end of the processes. Immobilized enzymes are an efficient alternative. Immobilization allows the reuse of enzymes, enables use in continuous processes, facilitates downstream, and enables increased process stability and storage. 11
With regard to immobilization, the choice of support is crucial for good process efficiency. Ion exchange resins should be mentioned as they allow rapid isolation and separation at the end of the reaction, the immobilization process is simple, and the interactions between enzyme and support are ionic and electrostatic. 12 Duolite® A-568 is a commercial resin classified as a weakly basic anion exchanger, phenol-formaldehyde cross-linked, has high porosity and is used as organic support for enzyme immobilization. 13 –15 This resin requires little treatment, is easy to handle in processes, and can also be reused.
In this context, the objective of this study was to produce the K. marxianus β-galactosidase enzyme by submerged cultivation, using whey permeate as a carbon source, in addition to immobilize the enzyme produced by adsorption on Duolite A-568 ion exchange resin.
Materials and Methods
Material
The microorganism used was the yeast K. marxianus ATCC 46537 from the Tropical Cultures Collection of the André Tosello Foundation of Campinas. Whey permeate acquired from SOORO was used as a substrate for cultivation. The carrier used for immobilization was the Duolite A-568 ion exchange resin (Rohm Haas, Philadephia, PA, USA), donated by Dow (Midland, MI). All other reagents used were of analytical grade.
Inoculum
The inoculum was cultured in a 250 mL conical flask using a culture medium with the composition (g L−1): yeast extract (6.0), (NH4)2SO4 (6.0), KH2PO4 (5.0), MgSO4 7H2O (0.6) and lactose (from whey permeate) (50) prepared in 0.2 M sodium phosphate buffer, pH 5.5. The useful volume for fermentation was 75 mL, being inoculated from the culture maintained in an inclined tube. The cell concentration was 2x108 cells mL−1. The culture was maintained at 30°C, 120 rpm for 14 h in a rotary incubator (Incubator 430 - Ethiktechnology) according to Santiago et al. 16
β-Galactosidase Production
The production of β-galactosidase was by submerged cultivation conducted at 30°C, under 300 rpm agitation, and 1 vvm aeration for 24 h, with an initial cell concentration of 5 g L−1. The experimental unit used was composed of a B. Braun Biotech International fermenter Biostat B, with a volume of 2.0 L, with a useful cultivation volume of 1.5 L.
Enzyme Extraction
For extraction of the β-galactosidase enzyme from K. marxianus which is intracellular, ultrasonic disruption was performed in an ultrasonic processor (High-Intensity Ultrasonic Processor, 500-watt model) with a frequency of 20 kHz and a maximum power of 150 W using a 40% titanium microelectrode tip. The fermented broth was centrifuged in a Beckman Coulter centrifuge model Avanti Centrifuge J-25 (27,200 g, 10 min), the decantated was resuspended in 0.2 M sodium phosphate buffer pH 7.3 to obtain a cell suspension with 2.62 mg mL−1. 17,18 The cell suspension was subjected to 30 min of ultrasonic pulses (total time of the 60-min process), with the replacement of the ice bath every 10 min. 19
To obtain the clarified enzyme extract, the cell suspension obtained after each bursting process was centrifuged (27,200 g, 10 min), the decanted liquid discarded, and the supernatant was used to determine the enzymatic activity.
Analytical Methods
Determination of cell concentration, lactic acid, lactose and ethanol
The cell concentration was obtained from the absorbance at 650 nm and converted to dry mass of cells using the standard dry biomass curve. Concentrations of lactic acid, lactose, and ethanol were determined by high performance liquid chromatography (HPLC). The sample was diluted, filtered, and injected into the chromatographic system, High Performance Liquid Chromatography-HPLC, Shimadzu model LC-20A Prominence, SUPELCOGEL C-610H column (Kyoto, Japan). Phosphoric acid (0.1%) was used as the mobile phase, with a flow of 0.5 mL min−1, oven temperature at 32°C and an injection volume of 20 μL. Concentrations of lactic acid, lactose, and ethanol were calculated using standard curves previously performed.
Determination of enzymatic activity
Initial lactose hydrolysis reaction rates were used to determine the enzyme activity of the enzyme β-galactosidase free and immobilized forms. 16 It was performed in a batch reactor using 75 mL of solution and 50 g L−1 lactose (PA), prepared in pH 6.5 lactic buffer at 30°C as described by Santiago et al. 16 In each experiment, 5 mL of the supernatant obtained after the enzyme extraction process was added to the reactor to determine the activity of the soluble enzyme. To the immobilized enzyme, a stainless steel basket containing 0.5 g of immobilized biocatalyst was added to the mixing reactor containing said lactose solution under the same conditions used for the free enzyme. The activity unit (U) was defined as μmol of glucose produced per minute (μmol min−1). The glucose oxidase method was used to measure the formed glucose. 20
Thermal Stability of free β-Galactosidase
Samples of the enzymatic extracts were incubated in a thermostatic bath (Thermomix B. Braun Biotech International) at different temperatures according to preliminary tests. The conditions were 30°C (24 h), 35°C (12 h), 40°C (4 h) and 50°C (2 h). The residual enzyme activity as a function of the incubation time for each temperature was adjusted to the first-order thermal deactivation models (Equation 1) and to one-stage thermal deactivation modelling (Equation 2). The adjustments were performed using the Levenberg-Marquardt numerical method using Statistica 7.0 software, determining the kinetic parameters through the best adjustments.
21
where A/A0 = Relative Activity and
To determine the activation energy of the thermal deactivation process, the results of the thermal deactivation constants (kd) as a function of temperature were adjusted to the Arrhenius (Equation 3) model using Software Origin® 7.0.
where A = frequency factor for reaction; Ea = thermal deactivation process activation energy; T = absolute temperature and R = contained in the gas law.
Stability of free β-Galactosidase In relation to the pH
The crude enzyme extract samples were lyophilized in a Liotop L101 freeze dryer and stored at −15°C until use. The lyophilized samples were resuspended in 0.2 M sodium phosphate buffer solutions at the following pH values: 6.0; 6.5; 7.0; 7.3; 7.5 and 8.0 and incubated in a thermostatic bath at 30°C for 12 h. At the end of this time, the residual activity of each sample was determined and related to the initial activity of the enzyme solution. Enzymatic activities in all experiments were determined according to Enzymatic Activity Determination.
Enzyme Immobilization
The immobilization process consisted of the adsorption of the enzyme on the Duolite A-568 ion exchange resin. Initially, the resin was activated according to the procedure described by Guidini et al. 22 A mass of 0.5 g of Duolite A-568 resin previously activated was added to enzyme extract solution (with volume and enzyme concentration defined for each experiment) prepared from the lyophilized enzymatic extract in sodium phosphate buffer varying the pH (according to the value defined for each experiment). The samples were kept under agitation at 150 rpm in a rotary incubator at 25 ± 1°C for 1 h. The immobilization conditions employed in relation to temperature, agitation and resin mass were followed according to the conditions proposed by Falleiros et al. 15
The recovered activity shown in Equation 4 was defined as a ratio between the activity observed in the immobilized enzyme and the activity of the initial soluble enzyme, according to Sheldon and Van Pelt.
23
The unit used in the calculations was U (μmol glucose min−1), considering that for all immobilization experiments a mass of 0.5 g of Duolite A-568 resin was used.
Optimization of K. marxianus β-galactosidase immobilization
To analyze the process of immobilization of K. marxianus β-galactosidase in Duolite A-568, a study of the influence of pH, immobilization buffer concentration, and Offered Enzyme Activity were carried out through a Central Composite Rotational Design (CCRD), with enzymatic activity of the immobilized biocatalyst the response evaluated. The CCRD was performed with two original levels, using Statistica 7.0 software, with 23 factorial points, 6 axial points, and 3 repetitions in the central point, totaling seventeen experiments.
Table 1 shows the values that were used in the experimental design for the three independent variables.
Real and Coded Values Used in the Central Rotational Composite Design
The initial pH and buffer concentration bands were determined according to Campello et al., and the offered enzyme activity was defined based on preliminary β-galactosidase immobilization tests. 24 A validation experiment performed in triplicate was conducted using the maximized condition.
Immobilization with the most concentrated enzymatic extract at pH 6.0 and 7.0
To obtain more concentrated crude enzyme extracts, the fermentation broth during 24 h of fermentation was centrifuged in a Beckman Coulter model Avanti Centrifuge J-25 centrifuge (27,200 g, 10 min). The decanted liquid was then resuspended in 250 mL phosphate buffer of 0.2 M sodium pH 7.0. The sample was disrupted in the ultrasonic processor according to method described above and centrifuged again. The supernatant was used for immobilization, and the decanted liquid was resuspended in sodium phosphate buffer and the rupture process resumed.
With the enzyme extract obtained, immobilizations were performed at pH 6.0 and 7.0 using a fixed enzyme volume of 20 mL, but the activity offered was quantified for each sample. The offered enzyme activity for immobilization were 139 and 181 U for pH 6.0 and 153 U for pH 7.0. The enzymatic activities and the recovered activities of each immobilized biocatalyst obtained were calculated.
Serial immobilizations of β-galactosidase enzyme
Serial immobilizations were performed, and three sequential immobilizations were made with the same enzyme solution to immobilize all the enzyme present in the solution. Thus, 0.5 g Duolite A-568 resin was used for 20 mL of crude enzyme extract prepared in 0.8 M sodium phosphate buffer pH 7.0. The strength of the buffer was determined at 0.8 M, as it was the best range for immobilization found in the optimization. The immobilization was performed for 1 h, 150 rpm, 25 ± 1°C. After the first immobilization, the resin was separated from the supernatant. The supernatant resulting from the first immobilization was again offered to a new resin of the same mass (0.5 g). The second immobilization was performed under the same conditions, then the same was done for the third immobilization. The resins of the three steps were washed separately with 0.8 M sodium phosphate buffer at pH 7.0, and the enzymatic activity of the biocatalyst obtained in each step was determined.
Reuse of Immobilized β-Galactosidase
To assess the reusability of the immobilized β-galactosidase, the immobilized biocatalyst obtained by the process described above was reused in seven consecutive lactose hydrolysis assays of 25 min each, being the biocatalyst sample was washed with 0.8 M sodium phosphate buffer pH 7.0 between hydrolysis reactions. The enzymatic activity determined in the first lactose hydrolysis was considered as 100% and the enzymatic activity achieved after each reuse was calculated in relation to the initial one.
Immobilization with the Commercial Enzyme
The commercial enzyme was immobilized in order to compare with results obtained in the immobilization process using produced and non-purified enzyme. The enzyme K. lactis β-galactosidase, Lactozyme® 2,600 L was purchased from Sigma Chemical Co. (Sigma-Aldrich, St. Louis, MO). The enzyme was immobilized under defined conditions (1 h, 150 rpm, 25 ± 1°C, pH 6.0 and 7.0). The offered enzyme activities were 34, 68, and 414 U for pH 6.0 and 112, 170 U for pH 7.0. The enzymatic solutions were prepared in 0.8 M sodium phosphate buffer. The enzymatic activities and also the recovered activity of the immobilized enzyme were calculated.
Stability after Storage of free β-Galactosidase and Immobilized Produced and Commercial Enzymes
Free produced β-galactosidase
Samples of the crude extract of β-galactosidase from K. marxianus in its free form were stored for seven weeks in liquid form, pH 7.3 under refrigeration at 8°C and freeze-dried at −15°C. The samples were analyzed weekly and the enzymatic activity determined.
Immobilized produced and commercial enzymes
The commercially available and produced β-galactosidase enzymes were immobilized and stored in 10 mL 0.8 M sodium phosphate buffer, pH 7.0 under refrigeration at 8°C and accompanied for 28 days.
For each analysis, enzymatic activity was determined. The initial activity of the immobilized enzyme prior to storage was considered to be 100%. After analyses, the enzymes were washed and stored again.
Results and Discussion
Cultivation Kinetics for β-Galactosidase Production
During cultivations conducted by K. marxianus, cell growth; pH of the medium; concentrations of lactose, ethanol, and lactic acid; and enzymatic activity were measured. These results are shown in Fig. 1.

In Fig. 1a, the pH of the medium decreased during cultivation, reaching a final value of 5.3, the reduction is explained by the production of lactic acid (3.3 g L−1) that leads to acidification of the medium at the end of 24 h. Figure 1b shows the complete lactose consumption within 15 h of cultivation. The enzymatic activity of β-galactosidase increased, reaching 127.09 U mLbroth −1 at 24 h of cultivation. The cell concentration reached a maximum value of 30 g L−1 in 9 h of cultivation. Interestingly, after cell growth is arrested, intracellular β-galactosidase activity continues to increase during the manutention phase. The concentration of ethanol at approximately 18 h of cultivation reached a value of 40 g L−1.
You et al. used whey milk as a carbon source for fermentation with K. lactis for β-galactosidase and ethanol production and achieved an enzymatic activity of 119.30 U mL−1 and ethanol concentration maximum 16.96 g L−1. These results corroborate those found in this study, with slightly higher enzyme activity and also concentrated in ethanol (127.09 U mL−1; 40 g L−1). 6 Murari et al. optimized ethanol production with K. marxianus using whey cheese and achieved ethanol production in the range of 12.64 to 29.78 g L−1; these results are in line with those obtained in this work. 25
Thermal Stability of Soluble β-Galactosidase
The relative activities as a function of incubation time are illustrated in Fig. 2, which shows that the temperature exerts a great influence on the stability of β-galactosidase produced by K. marxianus.
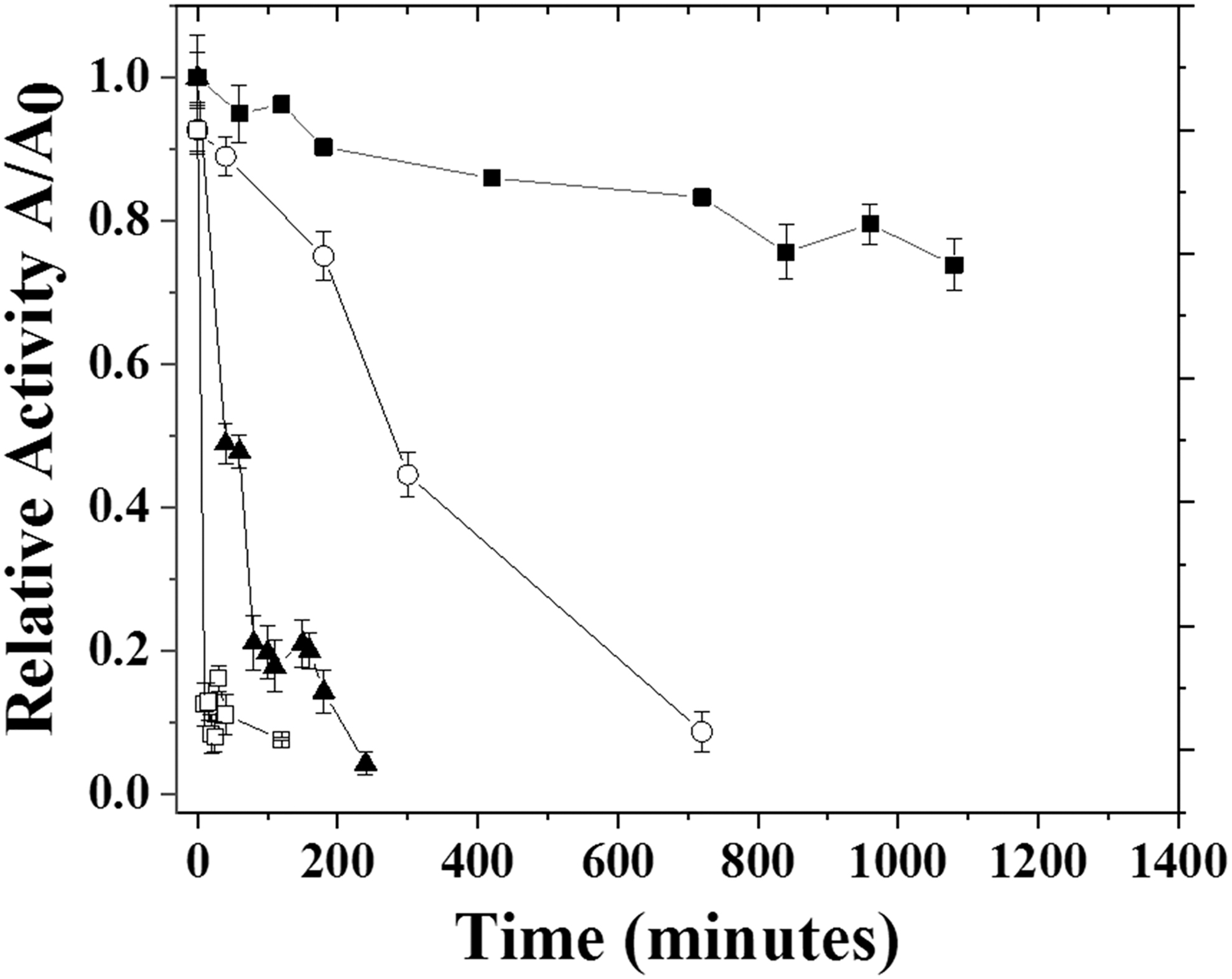
Thermal inactivation profiles for β-galactosidase soluble in ○30°C, ■ 35°C, ▲40°C, □ 50°C.
In Fig. 2, it can be observed that at temperatures of 40 and 50°C, there was a significant reduction of activity (greater than 80%), just after 1 h of incubation. At 35°C, the enzymatic inactivation was somewhat slower; the enzyme underwent thermal inactivation only after 12 h of incubation, and at 5 h, the loss of activity was about 60% relative to the initial activity. However, at 30°C, the enzyme maintained about 80% of its initial activity after 24 h of incubation.
According to the results of the thermal deactivation modeling at the evaluated temperatures, it is verified that the first-order thermal deactivation model (Equation 1) has been adjusted for all temperatures. However, the one-stage thermal deactivation model (Equation 2) was efficient for adjusting the results relative to temperatures of 30, 40, and 50°C. Using the thermal deactivation constants (kd), adjusted for the deactivation models, the half-life times for each temperature were calculated. The results are shown in Table 2.
kd Values and Half-Life Times Calculated by the First-Order Thermal Deactivation Model
The enzyme incubated at 30°C had the longest half-life (3,387 min). At 40°C, a considerable reduction in enzyme stability was observed, since the half-life was only 41.9 min; after 50°C, the reduction was at an extremely fast speed. There is a decrease in activity with temperature, with a half-life of just 2.1 min. With the values obtained and using the Arrhenius equation, it was possible to predict that at 25°C the half-life of the enzyme would be even greater—around 7,904.53 min. These results show that the enzyme β-galactosidase produced from K. marxianus is more stable at lower temperatures (30; 35°C), among those evaluated. It is therefore suitable for cold temperatures and can be useful, as milk processing takes place at low temperatures (around 5°C).
The linear adjustment was obtained with the correlation coefficient of 0.99. The value determined for the activation energy of the thermal deactivation was 64.13 kcal mol−1.
Stability of Soluble β-Galactosidase in Relation to the pH
The pH of the medium is very important and exerts a significant influence on the activity and catalytic stability of enzymes in general. The stability of the enzyme at pH is a factor that should be known for both proper storage and application in industrial processes. Figure 3 illustrates residual activity (%) after incubation for 12 h at 30°C in the pH range 6.0 to 8.0.
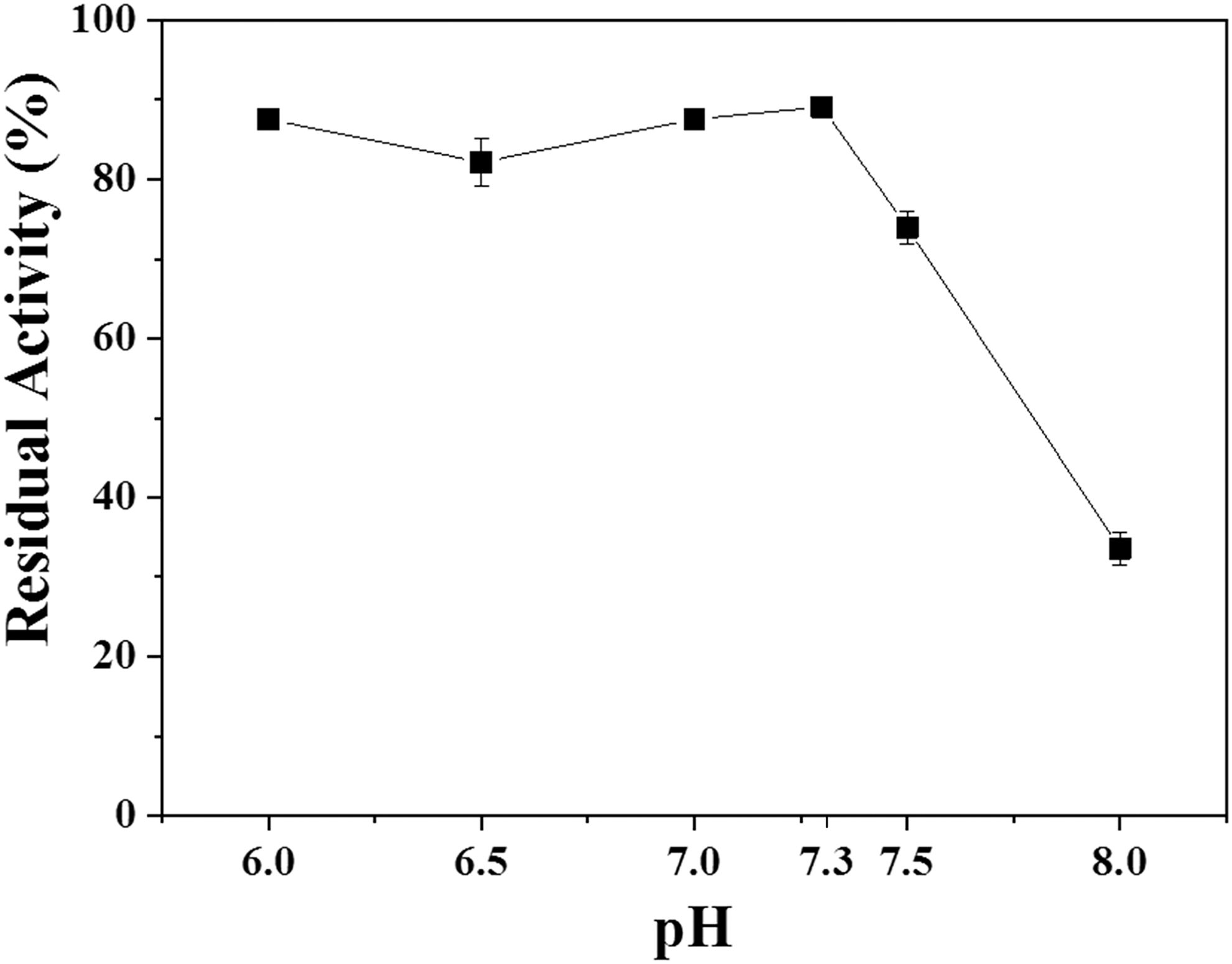
Influence of pH on the stability of soluble β-galactosidase at 30°C and 0.2 M sodium phosphate buffer in the pH range 6.0 to 8.0.
From Fig. 3 shows that the β-galactosidase K. marxianus in its soluble form was stable in the pH range between 6.0 and 7.3, maintaining its activity above 80%. The highest stability was obtained at pH 7.3, where the enzyme maintained approximately 90% of its initial activity even after 12 h of incubation. From pH 7.5, a decrease in enzymatic stability can be observed, and at pH 8.0 the enzyme showed a reduction of about 70% of its initial activity.
Thus, the enzyme β-galactosidase produced by K. marxianus has pH stability in the range of neutrality between 6.0 and 7.3. This pH range was in line with the literature. 26 The results obtained in the present work were similar to those obtained by Chanalia et al., which produced β-galactosidase from Pediococcus acidilactici and evaluated the stability of the enzyme in relation to pH at 37°C for 10 min at pH 4 to 10. The results obtained showed an activity greater than 97% in the pH range of 5.8 to 7.0 and 75% in the range of 5.5 to 7.5. 27
Optimization of K. marxianus β-Galactosidase Immobilization
When simultaneously analyzing the variables pH, buffer concentration, and activity offered, it was found that the enzymatic activities recovered in immobilization ranged from 2.51 to 19.24% presented in Table 3. The analysis of the significance of the model parameters was performed from the p-values found by the Student t-test, where X1 represents the pH, X2 the buffer concentration, and X3 the enzymatic activity offered in the immobilization. The CCRD was considered at a significance level of 95%. Using the results for immobilized activity recovered (AR) after multiple regression, Equation 5 was obtained with significant parameters.
Matrix with the Results Obtained from the Activity Recovered from the Immobilization Process of K. marxianus β-galactosidase
According to Equation 5, it can be seen that an increase in pH (X1), buffer concentration (X2), and activity offered (X3) negatively influence the recovered activity. The activity offered is the effect that most influenced the response of recovered activity, followed by buffer concentration and lastly the pH in the studied range.
The impact of negative effect obtained for variables (pH, buffer concentration and activity offered) on the studied process is reflected in recovery activity of immobilized biocatalyst, as can be observed when compared the experiments 9–10, 11–12 and 13–14 in pairs, in which extreme conditions (- α and + α) are used for each variable and the others maintained at the central level.
Analyzing the effect of pH (experiments 9 and 10), when pH 6.46 (-α) is used instead of pH 8.14 (+α), the recovery activity achieved is 4-fold higher. Regarding the buffer concentration (experiments 11 and 12), the increase employed between the conditions (0.10 M (-α) 1.50 M (+α)) implied a reduction of approximately 30% in the response. Finally, comparing experiments 13 and 14, which offering almost twice the enzyme activity in the immobilization solution (18.59 U (-α) to 35.41 U (+α)) the recovery activity decreased by half. Accordingly, within the studied range and evaluating in isolation, lower pH values (X1), buffer concentration (X2), and activity offered (X3), resulted in higher activity recovered, as predicted by the model.
The coefficient of determination (R 2 ), after adjusting the experimental data in the model, was 0.83, which indicates good agreement between the experimental values and those predicted by the model. From the model used, it was possible to create contour curves, showing the influence of pH, concentration, and buffer offered on the recovered activity in the process of immobilization of K. marxianus β-galactosidase.
In Fig. 4, the contour curves indicate the regions of greatest activities recovered, analyzing the influence of the studied variables. The best results were obtained for pH values between 6.8 to 7.2, buffer concentration between 0.1 to 0.8 M, and activity offered between 18.5 and 27 U. Campello et al. evaluated the immobilization of K. lactis β-galactosidase in Eupergit® and obtained as a negative influence of pH and positive influence of the buffer ionic strength and volume of enzyme offered. These authors reported the increase in pH from 6.6 to 7.4 represented a reduction of 18.19% in the immobilization yield, this result corroborates with those obtained in the present work. When comparing experiments 1 and 5, the increase of pH from 6.8 to 7.8 implied a reduction of activity recovered (∼16% lower), when the other variables were maintained constants (buffer concentration of 0.4 M and offered enzyme activity of 22 U). Analyzing experiments 1 and 3 in which the same enzyme concentrations (22.00 U) and pH values (6.8) were used, the increase of buffer concentration of 0.40 (−1) to 1.2 (+1) implied a reduction of approximately 20% of recovery activity obtained by the biocatalyst. Similar behavior can be observed at pH 7.8 by analyzing experiments 5 and 7, when the increase in buffer concentration from levels −1 to +1 resulted in a 50% lower recovery activity. 24
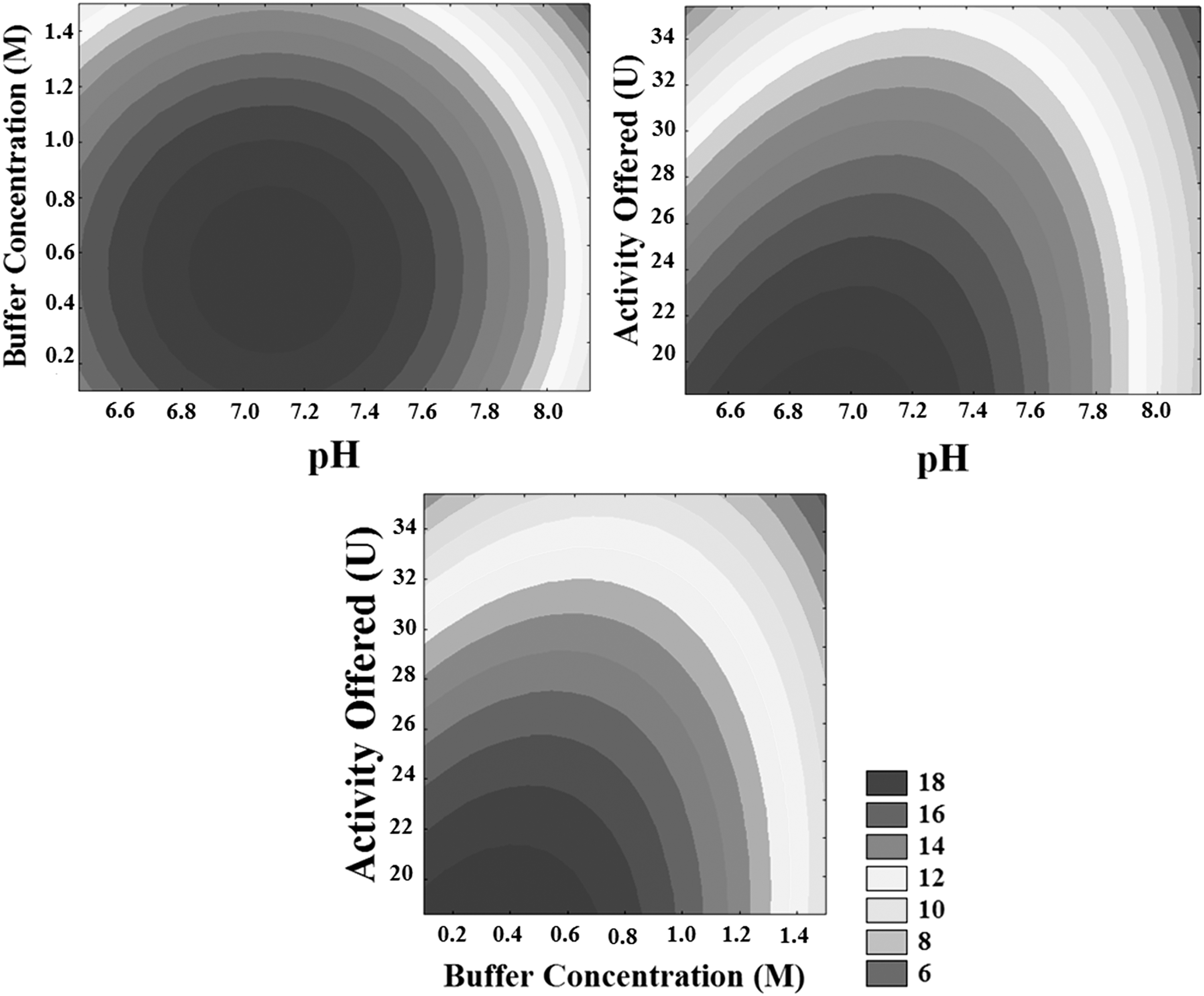
Contour curves of the influence of pH, buffer concentration and activity on the recovered activity of β-galactosidase immobilization.
Unlike pH, the effect of ionic strength obtained for Campello et al. did not agree with the present work because the authors concluded that the increase in ionic strength improved the immobilization yield. This result can be explained by the fact that Campello et al. used a hydrophobic support for immobilization and the increasing salt concentration created higher ionic forces that favored the hydrophobic surface. Thus, the difference between their study and the work in question can be associated with the use of supports Eupergit® and Duolite A-568 resin, respectively. 24
Buffer concentration promoted an increase in activity recovered up to 0.8 M. However, buffer concentrations greater than 0.8 M negatively affected the recovery activity of β-galactosidase immobilization. Adsorption is sensitive to variations in ionic strength; according to Leaver et al. the higher the ionic strength of the buffer (higher concentration), the smaller the amount of adsorbed substance, as there is a competition between the proteins and the ions present in the buffer solution for the resin adsorption site. On the other hand, high ionic strength can also lead to protein denaturation of the enzyme. Thus, it is clear that everything will depend on the protein adsorption process and also on the type of support used in the immobilization. It is necessary to carefully choose the variable to obtain the correct balance between maximum enzyme binding and support, and that denaturation is minimal. 28,24
Regarding pH, values above 7.3 negatively interfered with the activity of the immobilization process, a fact possibly related to the stability of the enzyme. This is consistent with the results of free enzyme stability in relation to pH. Figure 3 shows the most stable pH is close to 7.3. The activity offered negatively affects the activity recovered of β-galactosidase immobilization, as can be verified by Equation 5, but the optimal range was 18.5 to 27.0 U.
The average value of repetitions in the central point (pH 7.3, buffer concentration of 0.8 M and Offered Enzyme Activity of 27 U) resulted in 14.91% of activity recovered, too close (less than 1% variation) to the result obtained in experiment 13 using 18.59 U of Offered Enzyme Activity. Comparing these results with experiment 14, the increase of Offered Enzyme Activity to 35.41 U implied a reduction of activity recovered of about 46%.
A similar effect of initial enzyme concentration was observed by Liu et al., who evaluated the immobilization of K. fragilis β-galactosidase in magnetic nanometric epoxy for galactooligosaccharide synthesis, varying the enzymatic concentration from 1 to 6 mgprotein mL−1 in the immobilization. These authors concluded that the activity recovered from immobilization was maintained at 72% when employed enzymatic concentration from 1 to 3 mg of protein mL−1, after which a reduction of activity recovery, reaching 64%, was observed. 31 This phenomenon reported by Liu et al. and observed in the present work can be explained by the saturation of the support used, considering that the multiple layers of enzyme molecules present on the support surface (at conditions of high initial enzyme concentration) can block the active sites of the enzyme and cause diffusion limitation of substrate. 29
A model validation experiment was carried out in triplicate using the conditions of the best test results (pH 7.0, 0.8 M buffer concentration and 27 U activity offered). The average activity recovered in this experiment was 15.03 ± 0.12%, which was close to that of predicted values by the model (15.10%). The deviation between the values of the activities recovered was only 0.035%.
From the results of immobilization, it can be observed that the recovered immobilization activities ranged from 2.51 to 19.54% for the β-galactosidase enzyme of K. marxianus on Duolite A-568 ion exchange resin. In general, the recovered immobilization activity can be improved to achieve higher recovery values. These results may contribute to improving new studies regarding the support enzyme considering the great potential of commercial resin Duolite A-568, which has already been studied for the immobilization of β-galactosidase from different sources. 13,15,30,31
Immobilization with the most Concentrated Enzymatic Extract at pH 6.0 and 7.0
Tests were carried out with more concentrated enzymatic extracts to find high enzyme activities (Table 4).
Results Obtained from Immobilization with the Most Concentrated Enzymatic Extract
In these experiments, a higher enzyme supply was used at pH 6.0 and 7.0 to analyze the influence of enzyme concentration, offering an Enzyme Activity much higher than those evaluated in the CCRD. Results showed that, for pH 7.0 with an initial supply of 153 U, the enzymatic activity of the immobilized biocatalyst was 6.12 U, and the activity recovered of immobilization was only 4.00%. There was no increase of the immobilized enzymatic activity nor an increase in the value of the activity recovered compared with results obtained in the section entitled Optimization of K. marxianus β-galactosidase immobilization with an enzymatic offer of 27 U.
Experiments were performed at pH 6.0 to verify possible ionization forms of the resin, considering that the enzyme in its free form also presented stability at this pH. According to the obtained results, it can be observed that at pH 6.0 there was also no increase of immobilized enzymatic activity nor an increase in the value of the activity recovered. The enzymatic activities evaluated both at pH 6.0 (139 U) and at pH 7.0 (153 U) showed similar values.
Analyzing the enzymatic offer of 181 U and pH 6.0, we observed a small increase of the enzyme activity of the immobilized enzyme (7.17 U). However, the activity recovered was low, being only 3.96%, which shows that even with a very high enzyme offer, the ion exchange resin is not able to retain all enzyme by the adsorption process under the conditions employed. The above results showed a possible saturation of the immobilization support, in this case Duolite A-568 resin. After immobilization, it was also observed that the supernatant still had a high activity since little enzyme was adsorbed by the support. In this way, a series of immobilizations was made to reuse the supernatant.
Enzyme concentration is important for immobilization, and several studies have addressed this variable. Suárez et al. evaluated the immobilization of Aspergillus oryzae β-galactosidase in glyoxyl-agarose, offering different enzymatic concentrations (1, 10, and 30 mg protein g support−1) in fine and macro supports. Results showed that the best protein immobilization yield for both types of the supports was higher for the concentration of 1 mg protein g support−1, being 93.4% (fine) and 94.3% (macro), and for the concentration 30 mg protein g support−1, there was a reduction, falling to 66.2% (fine) and 73.2% (macro), which was explained by the saturation of the support. 32 The results are similar to those obtained for β-galactosidase from K. marxianus in the present work, which also showed a reduction in the activity recovered from immobilization considering an increase in enzyme concentration. With the offered enzyme activity in immobilization from 139 to 181 U, about 4.0% was obtained to activity recovered, whereas, the results from CCRD, with offered enzyme activity in immobilization from 27 U obtained 14.9% activity recovered, reporting the saturation of the support, an excess of an offered enzyme that can block the enzyme's active site.
Cargnin, Gasparin, and Paulino performed a Central Composite Rotational Design (CCRD) to evaluate the influence of temperature, initial enzyme concentration, and pH on the immobilization of K. lactis β-galactosidase in pectin hydrogels. 33 The initial enzyme concentration exerted positive influences on the immobilization capacity of the hydrogels. The immobilization was optimized under conditions of 7.0% and 6.3% of initial enzyme concentration for hydrogels without pine fiber and for hydrogels with 5 and 10% pine fiber, respectively. The results were fundamental to predicting immobilization, emphasizing the importance of assessing enzyme concentration during immobilization. 33
Serial Immobilizations of β-Galactosidase Enzyme
As a strategy to maximize the utilization of the enzyme extract produced, immobilizations were performed in series, that is, three immobilizations were made in a row with the same enzyme solution. The results are presented in Fig. 5.
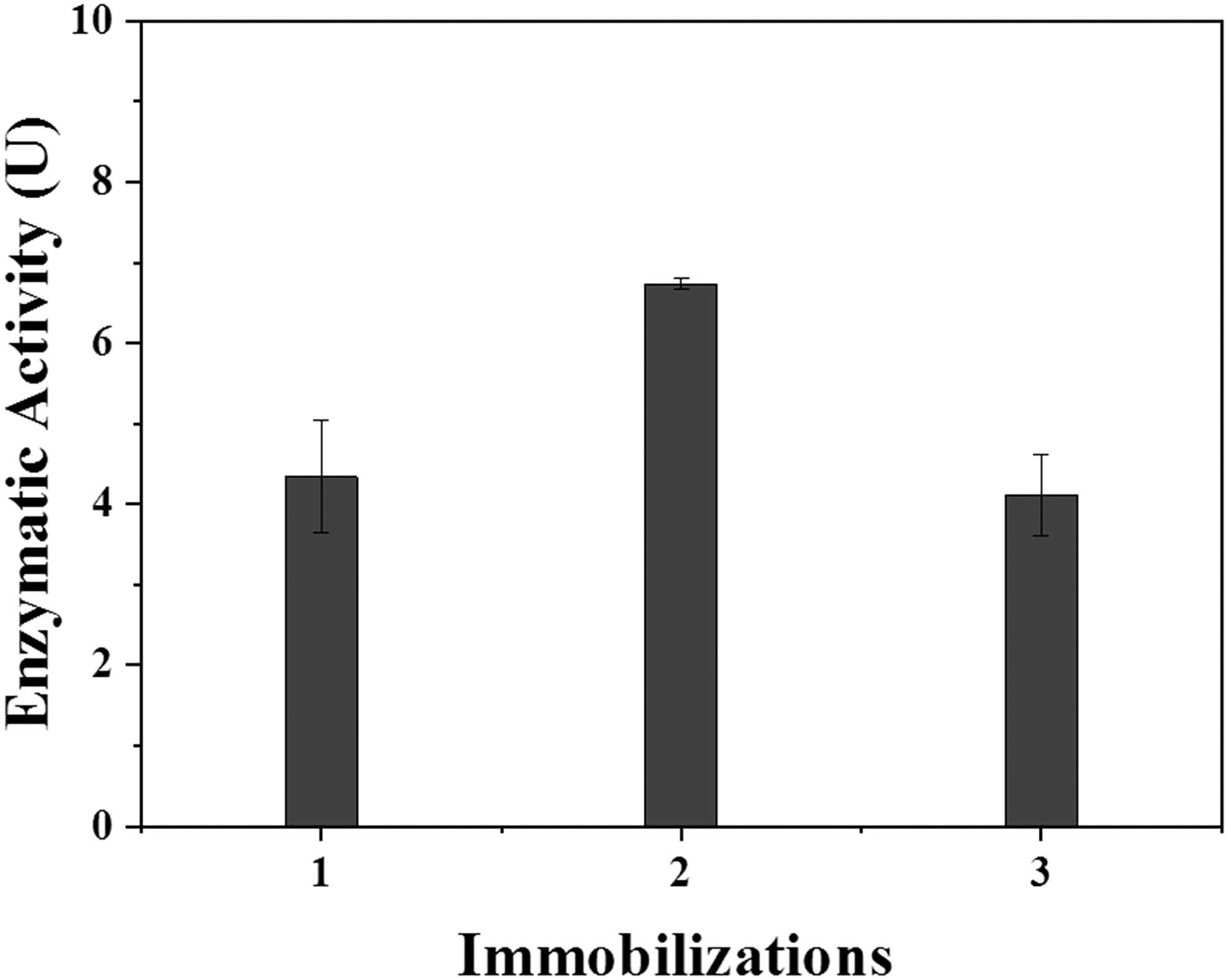
Enzymatic activity of serial immobilization of the enzyme β-galactosidase.
According to the results presented in Fig. 5, considering the initial supply of 45 U of activity offered, the three immobilizations presented similar enzymatic activities; in the first immobilization, 4.34 U and 9.64% was obtained, respectively, while the second immobilization presented 6.70 U and 14.89% and the third 4.1 U and 9.13%. This increase in enzymatic activity and activity recovered in the second immobilization may be justified by the increase in purity of the enzymatic extract, since it may contain interfering agents because it is not purified, and thus part of the impurities showed an affinity with the resin. These impurities may have been retained in the first immobilization and in the second immobilization more enzymes (β-galactosidase) were adsorbed by the resin, increasing the enzymatic activity. Thus, serial immobilization can be considered as an alternative to increasing the immobilization of all enzymes present in the enzymatic crude extract, which is extremely valuable considering the high costs of obtaining the enzymes. In addition, the immobilized biocatalysts of each of the three serial steps can be used together, containing a greater amount of enzyme to be applied to the processes and a greater recovery.
It is important to mention that the enzyme was immobilized in its crude form, without pre-purification, which can hinder the physical adsorption process. Processes such as serial immobilization are alternatives that can remedy difficulties in immobilizing the crude enzymes, achieving greater utilization enzyme, without requiring more expensive steps, such as purification. The purification processes are usually quite expensive; therefore many are developing one-step of the purification and immobilization. Zhao et al. evaluated the purification and immobilization of EndoS enzymes using cellulase in one step. Qin et al. used a carbohydrate-binding (CBM56-Tag) as a mediator of immobilization and purification of fusion enzymes. The serial immobilization could be used in this same sense in view of the exposed results. 34,35
This method brings a new perspective in the usage of the enzymes and can be used as a technique for purification of joint immobilization, which can reduce industrial costs and facilitate the processes.
Reuse of Immobilized Produced β-Galactosidase
The immobilized β-galactosidase enzyme was evaluated for its reuse in the hydrolysis process, being used in seven cycles of 25 min each. Reuse is an important characteristic of the industrial application of enzymes. The more the enzyme maintains its activity in the processes, the more advantageous is its application. The immobilized β-galactosidase maintained about 70% of its initial activity for 4 cycles, reaching about 30% of its initial activity after 7 cycles.
Braga et al. also evaluated the reuse of immobilized β-galactosidase from K. marxianus in Eupergit® and obtained the relative activity of about 50% after 5 cycles. Compared with results obtained from immobilization in Duolite® A-568, it is observed that the immobilized enzyme was able to maintain a higher activity for 5 cycles. 5 Albuquerque et al. studied or reused K. lactis β-galactosidase immobilized in chitosan, or reused in the conversion of lactose in cheese whey, reaching values of 50% in the second cycle and only 20% in the sixth cycle—lower values than the tests with the β-galactosidase immobilized on Duolite A-568. 36 Studies related to the reuse of biocatalysts are feasible in the process. The possibility of reusing enzymes and reducing costs is interesting, considering the high added value of the enzymes.
Immobilization with Commercial Enzyme
Immobilization was evaluated with the commercial enzyme Lactozyme® at different pH values and offered enzymatic activities. The performance evaluation of the commercial enzyme was intended to make a comparison with the immobilization of the enzyme previously produced without purification. The results are shown in Table 5.
Results of Commercial β-galactosidase Immobilization Lactozyme in Different pH and Activities Offered
The recovered activity was increased with the increment of offered enzyme activity at pH 6.0. The highest activity recovered was 12.22% obtained for an activity offered of 414.0 U. Thus, it can be concluded that the enzyme was not completely adsorbed on the resin even with a very high initial activity (414.0 U). For the experiments at pH 7.0, the highest activity recovered was 19.47% of the initial supply of 170 U.
By doing a general analysis for both pH 6.0 and 7.0, it can be observed that the activities recovered were small, all being less than 20.00%. These results are similar to those obtained in this study of optimization of β-galactosidase K. marxianus immobilization, where the highest recovered activity was 19.24%. These values for recovered activities and larger enzymatic activities were expected considering that the commercial enzyme is purified and has no interference with the medium as can occur in the unpurified produced enzyme. Thus, the potential of application of the β-galactosidase enzyme produced without any purification method is evident, considering that the enzymatic purification methods are usually expensive and laborious. It is clear from these results that both the commercially available purified enzyme and the produced unpurified enzyme presented very few differences regarding the activities recovered from immobilization.
According to the results obtained and in respect of all the variables analyzed, it can be observed that the β-galactosidase enzyme of K. marxianus presented the possibility of immobilization in a Duolite A-568 ion exchange resin even with low activity recovered from immobilization. The study has great importance for the advancement of new immobilization techniques using commercial resins.
Therefore, it is interesting to continue studies with this enzyme and the support since both have advantageous characteristics and great industrial applications. Thus, it is necessary to try other techniques of immobilization with the difference in the activation of the support to expand the application of the method and to achieve better immobilizations that satisfy the industrial applications, generating quality products and reducing costs in its application.
Stability to the Storage of free β-Galactosidase and Immobilized Produced and Commercial Enzymes
Free β-galactosidase produced from K. marxianus
It was observed that after seven weeks of storage the free enzyme and the free lyophilized enzyme maintained residual activities above 80%, showing that both forms of the enzyme are suitable for storage. Thus, the choice between lyophilized and non-lyophilized should be made to best meet the process conditions under which β-galactosidase will be applied. Jochems et al. evaluated the stability of soluble β-galactosidase of K. lactis at 5°C in tris-HCL buffer. Over a period of seven days, the soluble enzyme maintained 46% of the initial activity and after 14 days the enzyme had already lost 80% of its initial activity. Comparing these results with those obtained in the current work, it can be observed that the non-lyophilized and lyophilized enzyme maintained about 80% of initial activity over a period of approximately 50 days. 37
Immobilized produced and commercial enzymes
The produced immobilized β-galactosidase stored under refrigeration suffered a sharp reduction in its activity in the first days of storage. In 3 days, the enzyme had already reduced its initial activity by about 60%. In 12 days, the activity relative to the immobilized enzyme was practically zero.
The immobilized commercial β-galactosidase, also stored under refrigeration, underwent significant reductions over the course of days. In the first 7 days, the enzyme lost around 80% of its initial activity, and at 21 and 28 days the activities of the samples remained constant, but with a very low enzymatic activity around 3% of the initial enzymatic activity. Thus, for both immobilized enzymes, buffer storage and refrigeration were not an efficient techniques to maintain enzyme activity. It would be relevant to test other storage methods by means of increasing the shelf life of β-galactosidase immobilized on the exchange resin Duolite A-568.
Conclusions
β-galactosidase was produced efficiently by K. marxianus using whey permeate, reaching 127.09 U mLbroth −1 at 24 h of cultivation, associated with synthesizing other added-value metabolites: lactic acid (3.3 g L−1) and ethanol (40 g L−1), that can contribute to the valorization of subproduct utilized as substrate. The enzyme obtained showed greater thermal stability at 30°C, pH of 6.0 to 7.3, and storage stability for 7 weeks, maintaining about 80% of its initial activity.
The immobilization of β-galactosidase produced in ion exchange resin demonstrated strong interference from the analyzed parameters, pH, buffer concentration, and activity offered. Serial immobilization increased the enzymatic immobilization of the crude extract. In relation to reusability, the immobilized biocatalyst maintained about 70% of its initial activity for 4 cycles. All the characteristics evaluated are essential for the application of β-galactosidase in industry; however, more studies are needed to improve the immobilization of β-galactosidase in Duolite A-568 resin from K. marxianus, being a promising alternative for industrial application.
Footnotes
Acknowledgments
The authors wish to thank Dow Chemical Brazil S.A. for the resin donation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors wish to thank the CNPq, CAPES and FAPEMIG (Brazil) for the financial support.