
Abstract
The use of microorganisms in industrial fermentations requires robust strains tolerant to stresses that challenge its performance during the bioprocess. One approach to obtain such a strain, adaptive evolution methodology, is carried out in this work with an emphasis on the biochemistry of stress tolerance. This work evaluated the robustness and cellulosic ethanol efficiency of an evolutionary adapted strain of Scheffersomyces stipitis NRRL Y-7124 (HAJ) obtained after successive batch cultures with increasing concentrations of acid hydrolysate lignocellulosic jojoba residue. Strain robustness was associated with its ability to tolerate stresses present along an industrial cellulosic bioethanol production process (i.e., thermal, oxidative or osmotic stress; high concentration of ethanol or phenolic compounds). Under such conditions, HAJ exhibited 4-fold higher viability and 8-fold higher vitality (metabolic performance) values than the parental strain. Whereas all stresses assayed produced a significant increase in reactive oxygen species (ROS) concentrations in Y-7124 (up to 15-fold higher than controls), in HÁJ only ethanol induced a significant rise in ROS levels, associated to variations in superoxide dismutase (SOD) and catalase (CAT) enzymatic activities. The highest increase in SOD activity was associated with ethanol stress, the most oxidative stress assayed, being 3.5-fold higher in HAJ versus Y-7124. Intracellular concentrations of cell protectants trehalose and glycogen increased significantly after stresses related to hydric deficiencies (sorbitol and ethanol), with HAJ showing a higher increase than the parental strain. Ethanol production efficiency on a non-detoxified, non-supplemented acid-hydrolyzed lignocellulosic medium was 40% higher for HAJ versus Y-7124. Our results propose that stress cross-tolerance of this yeast is associated to its oxidative stress tolerance, and that high levels of molecules like trehalose should be a goal for obtaining a robust strain that can be used industrially.
Introduction
Scheffersomyces (Pichia) stipitis is an aerobic, Crabtree-negative yeast, well known for its capacity to ferment C5 and C6 sugars to efficiently produce ethanol and having one of the highest native abilities for xylose fermentation, with substrate yields between 0.35–0.44 g/g. 1 -3 In S. stipitis, ethanol production is favored under oxygen-limited (microaerobic) conditions. 4 Due to its ability to ferment both C5 and C6 sugars, S. stipitis can be efficiently used for bioethanol production from lignocellulosic substrates containing large amounts of these sugars. 5,6 Standard acid hydrolysis pretreatments of lignocellulosic biomass result in hydrolysates rich in hexoses (principally glucose and galactose) and pentoses (mainly xylose and arabinose). 7 As a consequence of this treatment, several toxic and growth-inhibiting compounds are also produced, including furaldehydes, aliphatic acids, and phenolic compounds. 8 For its industrial application, it is essential for S. stipitis to have the capacity to tolerate those inhibitors, as well as a wide variety of stress conditions. Simultaneously, it should maintain an adequate metabolic performance during the fermentative process to increase the bioconversion yield of cellulosic sugars to ethanol.
Enhancing yeast performance in the presence of inhibitors or stressors requires the selection of yeast populations with better vitality (overall physiological and metabolic fitness). Adaptive evolution allows the selection of yeast populations resistant to different stress conditions and with an expanded tolerance range.
9
–14
Tolerance to oxidative stress-generating reactive oxygen species (ROS) seems to be a common mechanism of multiple stress tolerance.
9
ROS are generated as byproducts of cellular aerobic metabolism,
15
with the three most common being superoxide (O2
•-), hydroxyl radical (OH•), and hydrogen peroxide (H2O2). These can damage a variety of cellular components, including nucleic acids, proteins and lipids, eventually leading to yeast cell death.
16,17
Under normal physiological conditions, yeast can respond to ROS, overcoming oxidative stress by diverse mechanisms combining ROS scavengers such as glutathione, thioredoxins and dehydroascorbic acid,
18
–22
and antioxidant enzymes such as catalases (CAT), superoxide dismutase (SOD) and glutathione peroxidase. CAT and SOD are considered the most active antioxidant enzymes in response to environmentally stressful conditions, due to direct decomposition of ROS, and SOD, usually considered the first line of defense against oxidative stress.
23
-25
Together with the defensive mechanisms mentioned above, the non-reducing disaccharide trehalose has been shown to play a main role in yeast cell protection against a variety of stresses, including exposure to oxidative agents.
26
-29
Trehalose (a non-reducing disaccharide) appears to be important during both the direct stress challenge and subsequent stress recovery
30,31
by stabilizing cellular structures.
32
On the other hand, high trehalose levels could lead to glycolysis dysfunction, decreased pyruvate kinase activity (affecting the redox balance/homeostasis of the cell
Several studies have reported that previous yeast exposure to one mild stress condition can increase tolerance against challenges by the same, or different stress, even when applied at a higher level—a phenomenon known as stress cross-tolerance. 14,34 –37
Traditional ethanologenic industrial yeasts do not ferment C5 sugars and are not able to grow in concentrated lignocellulosic hydrolysates due to their high sugar concentrations (>100 g/L) and the toxic byproducts generated during lignocellulosic substrate pretreatments. 6 As mentioned, S. stipitis is one of the most viable natural pentose-fermenting yeasts available for cellulosic ethanol production. 3 However, its stress tolerance needs improvement to be able to resist osmotic pressure and high ethanol concentrations. 38
In view of this evidence, the objectives of this study were, first, to improve the robustness of a strain of S. stipitis in industrial bioethanol production from lignocellulosic feedstock through an adaptive evolution strategy; second, to explore whether it is possible to use this methodology to obtain a yeast cell population of S. stipitis co-tolerant to the different stress conditions that may be found during bioethanol industrial production; and finally, to gain insight into the relationship between oxidative stress resistance mechanisms, synthesis of stress protectant molecules, and multiple stress tolerance.
Herein we present an evolutionarily adapted S. stipitis strain in which the concomitant action of antioxidant enzymes and cell membrane-protecting molecules makes it tolerant to, among others, high ethanol and osmotic stress. These traits, together with its increased thermotolerance, make it a suitable yeast strain for economically viable industrial cellulosic bioethanol production.
Materials and Methods
Strains
Scheffersomyces (syn Pichia) stipitis NRRL Y-7124 (USDA, Northern Regional Research Laboratory Culture Collection) was provided by Dr. C. Kurtzman and used for the adaptive evolution procedure. Both the parental wild-type strain and the adapted clone were maintained on petri dishes containing YPD medium (1% w/v yeast extract, 1% w/v meat peptone, 2% w/v glucose, 2% w/v agar) at 4°C and stored in glycerol 25% v/v at -80 ± 1°C.
Unless indicated otherwise, all measurements were carried out at 28 ± 1°C. Cell counting was performed in a Neubauer chamber mounted in saline. All submerged cultures were carried out in 250 mL Erlenmeyer flasks containing medium at a 1:5 medium volume to flask volume ratio, in an orbital shaker at 150 rpm. Each experiment was done in triplicate and the data presented are the average.
Preparation of Jojoba Cake Hydrolysate
Residual cakes from chemical extraction of waxes from jojoba (Simmondsia chinensis) seeds—containing 11.1% w/w cellulose, 12.0% w/w hemicellulose, 16.7% w/w total non-structural carbohydrate, 28.0% protein and 24.9% w/w lignin—were grinded to less than 1 mm particles. The ground residual cake was hydrolyzed in 0.5 N sulfuric acid with a dry weight/volume ratio of 1:5, for 3 h at 100°C, and filtered successively through gauze and filter paper. The pH of the acid hydrolysate was brought to 4.8 with 10 N NaOH and sterilized by autoclaving. This lignocellulosic hydrolysate, identified as jojoba cake hydrolysate (JCH), contained ∼4.0 g/L of glucose (measured using an enzymatic kit from Wiener Lab, Rosario, Argentina), ∼40.0 g/L of total pentoses (determined by the method described in Deschatelets and Yu), 39 ∼1.81 mg/L of total phenols (method adapted from Singleton et al.), 40 and ∼1.35 g/L furans (as described in Martinez et al). 41
Adaptive Evolution Of S. Stipitis in Hydrolysate
The parental strain S. stipitis NRRL Y-7124 was grown on YPD medium in an orbital shaker at 150 rpm for 24 h. The culture was used to inoculate a 250-mL Erlenmeyer flask containing 50 mL of an adaptation medium composed of yeast nitrogen base (YNB, BD Difco, Franklin Lakes, NJ) and 10 % v/v JCH. After 24 h incubation, this culture was used as inoculum for a new, 24 h culture with the same concentration of hydrolysate. This procedure was repeated in successive passages using increasing concentrations of hydrolysate, at 5% increments up to 90 % v/v, implying 34 culture transfers and approximately 300 generations. Cultures were started with low biomass concentration (OD600nm ≈ 0.1) to select a cell population with better adaptability or detoxification capacity on this hydrolysate. Growth was followed spectrophotometrically (OD600nm) with an UV-Visible Spectrophotometer T60 PG Instrument. This growth determination method was routinely employed (unless indicated otherwise). Samples were withdrawn from the last culture in which growth occurred and stored at −80 ± 1°C in 25% v/v glycerol solution for further assays. The clone adapted to the JCH was named S. stipitis HAJ.
Tolerance to Stress Conditions
To evaluate the robustness of the adapted clone to the hydrolysate of jojoba cake compared to its parental strain, tolerance to a set of stress conditions was evaluated. The stressors studied consisted of inhibitors present in lignocellulosic hydrolysates, and others that a yeast might face in a bioethanol production process typical of alcoholic fermentation. Briefly, cultures of S. stipitis Y-7124 and HAJ (in YPD and JCH, respectively) were diluted to obtain a final OD600nm of 1.0. Then, cells were washed three times with saline solution (0.9% v/v NaCl), centrifuged at 7,500 g for 10 min, and resuspended in 0.1 mol/L potassium phosphate buffer (pH 6.0) before being subjected to the following stress conditions: ethanol (20% v/v, 30 min), oxidative stress (0.3 mol/L H2O2, 1 h), osmotic shock (2.5 mol/L sorbitol, 4 h), thermal shock (55°C, 5 min) and phenolic acid (0.6 g/L vanillin, 1 h). After treatments, stressors were removed by washing the cells with 0.1 mol/L potassium phosphate buffer (pH 6.0) or by ending the experiment in a water bath at 25°C. The stressors were selected due to their presence in the acid hydrolysate (phenols); for the potential use of the adapted strain in very high gravity (VHG) fermentation (osmotic stress) or in simultaneous saccharification and fermentation (SSF) (thermal stress); for their known oxidative capacity (hydrogen peroxide); or because they are products of fermentative activity (ethanol). To study the effect of these stressors, the following determinations were carried out on the treated cells: viability; vitality; antioxidant enzyme activities; intracellular concentrations of ROS, trehalose, glycogen, NAD/NADH and NADP/NADPH; and pyruvate kinase activity.
Cell Viability
Serial dilutions of the stressor-treated cells were carried out with distilled water from 10−1 up to 10−7. Viability was determined in triplicate by plating 10 μL of each dilution on YPD agar. After a 24 h incubation at 28 ± 1°C, colonies were counted and averaged for each dilution. Cell viability was reported as the survival percentage 42 in comparison to the unstressed cells (control).
Cell Vitality
Vitality of the treated cells was assessed to determine their metabolic performance by using the acidification power test devised by Opekarová and Sigler. 43 Briefly, cells were harvested after stressor exposure by centrifugation, washed three times with saline solution, and quantified in a Neubauer chamber. Dilutions were made with sterile distilled water to reach a final concentration of 5 × 107 cell/mL, and 5 mL of each dilution was transferred to 15 mL Falcon tubes. pH was measured every 1 min for 10 min, after which a glucose solution was added to a final concentration of 1% v/v, and pH measurement continued for another 10 min. Acidification power was determined as AP = pH t=0min – pH t=20min.
Oxidative Parameters
Cell free extracts were prepared using the method described by Gurdo et al. 14 Intracellular ROS concentration, and CAT and SOD activities were quantified in the supernatants of the cell homogenates using methods described by Zhang, 44 Aebi, 45 and Beauchamp and Fridovich, 46 respectively.
Protein Content
Total soluble protein content was determined in the supernatants according to Bradford, 47 using bovine serum albumin as standard (Sigma Aldrich, St. Louis, MO).
Trehalose and Glycogen Measurement
Intracellular trehalose and glycogen were quantified as described in Parrou et al. 48 Briefly, 1.0 × 108 cells of each treated culture were centrifuged in 1.5 mL tubes at 20,000 x g for 1 min. After discarding the supernatants, pellets were washed with 1 mL saline solution and centrifuged again. Pellets were resuspended in 125 μL of 0.25 mol/L Na2CO3 solution and incubated 3 h at 95 ± 1°C. After cooling to room temperature, 300 μL of 0.2 mol/L CH3COONa (pH 5.2) and 75 μL of 1 mol/L acetic acid were added, bringing pH to 5.5. The content of each tube was then divided into 2 tubes (250 μL/tube). In one of these tubes, trehalose concentration was determined by adding 15 μL of 0.2 mol/L CH3COONa (pH 8.0) and 3 μL of porcine trehalase (Sigma-Aldrich). Following overnight incubation at 37 ± 1°C, glucose was measured using an enzymatic kit (Wiener Lab, Rosario, Argentina). Glycogen content was determined by addition 10 μL of an α-amyloglucosidase solution (∼70 U/mg) to the remaining tube. After overnight incubation at 57 ± 1°C glucose concentration was measured using the mentioned enzymatic kit.
Pyruvate Kinase Activity and Intracellular Oxidoreductase Coenzymes Levels
Pyruvate kinase (PK) activity, and levels of NADH/NAD and NADPH/NADP were measured spectrophotometrically as described in the corresponding kits protocols (Sigma-Aldrich MAK072, MAK037 and MAK312, respectively) in cell-free extracts of Y-7124 and HAJ grown in YPDX or JCH, respectively. PK activity is reported as milliunit/mL, where one milliunit (mU) of pyruvate kinase is defined as the amount of enzyme that will transfer a phosphate group from PEP to ADP to generate 1.0 nmole of pyruvate per minute at 25°C.
Bioethanol Production Efficiency
The ethanologenic efficiencies of S. stipitis Y-7124 and HAJ clone were compared using two fermentation media: 100% JCH and semisynthetic medium YPDX (0.5% w/v yeast extract, 0.5% w/v peptone, 0.4% w/v glucose, 4.0% w/v xylose). 100 mL Erlenmeyer flasks containing 80 mL of either JCH or YPDX were adjusted to pH 4.5 and inoculated to an initial cell concentration of 1.0x106 cell/mL. After incubating in an orbital shaker at 28 ± 1°C and 100 rpm for 72 h, samples were withdrawn from the cultures and then centrifuged at 7,500 g for 10 min. Ethanol was quantified from the culture supernatants using an enzymatic kit (R-Biopharm AG, Darmstadt, Germany), and total reducing sugars were measured by the DNS method to determine volumetric production of ethanol, sugar consumption, volumetric productivity, and fermentation efficiency (Fe). The latter was calculated according to the following formula:
where E is the final ethanol concentration in the fermented medium [g/L]; S is total reducing sugars consumed [g/L], and 0.511 g/g is the maximum theoretical yield of ethanol from 1 mol of glucose or xylose.
Statistical Analysis
All experiments were performed in triplicate, unless stated otherwise. Data were analyzed by two-way ANOVA followed by Fisher's LSD (Least Significant Difference) test, using GraphPad Prism 6.01 (GraphPad Software, La Jolla, CA). Logarithmic transformation of CAT, SOD and ROS data was needed to comply with homoscedasticity. Values in graphics are shown as mean ± standard error.
Results and Discussion
Strain Adaptation in JCH
The adaptation of S. stipitis NRRL Y-7124 was performed in successive batch cultures with increasing concentrations of JCH. This procedure allowed the adaptation of the S. stipitis strain to the jojoba hydrolysate through metabolism adaptation or spontaneous mutations that could be driven by the inhibitors and toxic compounds present in this hydrolysate. As a result of this evolutionary adaptation strategy, the adapted clone HAJ was obtained, capable of attaining growth levels in 90% JCH of >4.0 absorbance units (starting from OD600nm ≈ 0.5) within the first 24 h of culture, whereas the parental strain Y-7124 showed no growth until 48 h in the same culture medium.
Viability and Vitality Under Stress Conditions
As shown in Fig. 1, all stressors tested significantly decreased viability and fermentative performance (vitality) of both strains in different percentages. Thus, viability values for strain Y-7124 ranged from below 7% survival for temperature and 13% for ethanol, up to 40% for vanillin, indicative that the most severe stresses, among those assayed concerning cell duplicative capacity, were thermal shock and ethanol, with phenolic stress being the less severe. On the other hand, compared to Y-7124, clone HAJ showed higher viability under all the stress conditions tested, from ∼2.0 fold for ethanol and temperature stress up to ∼4.0 fold for osmotic stress (Fig. 1a).
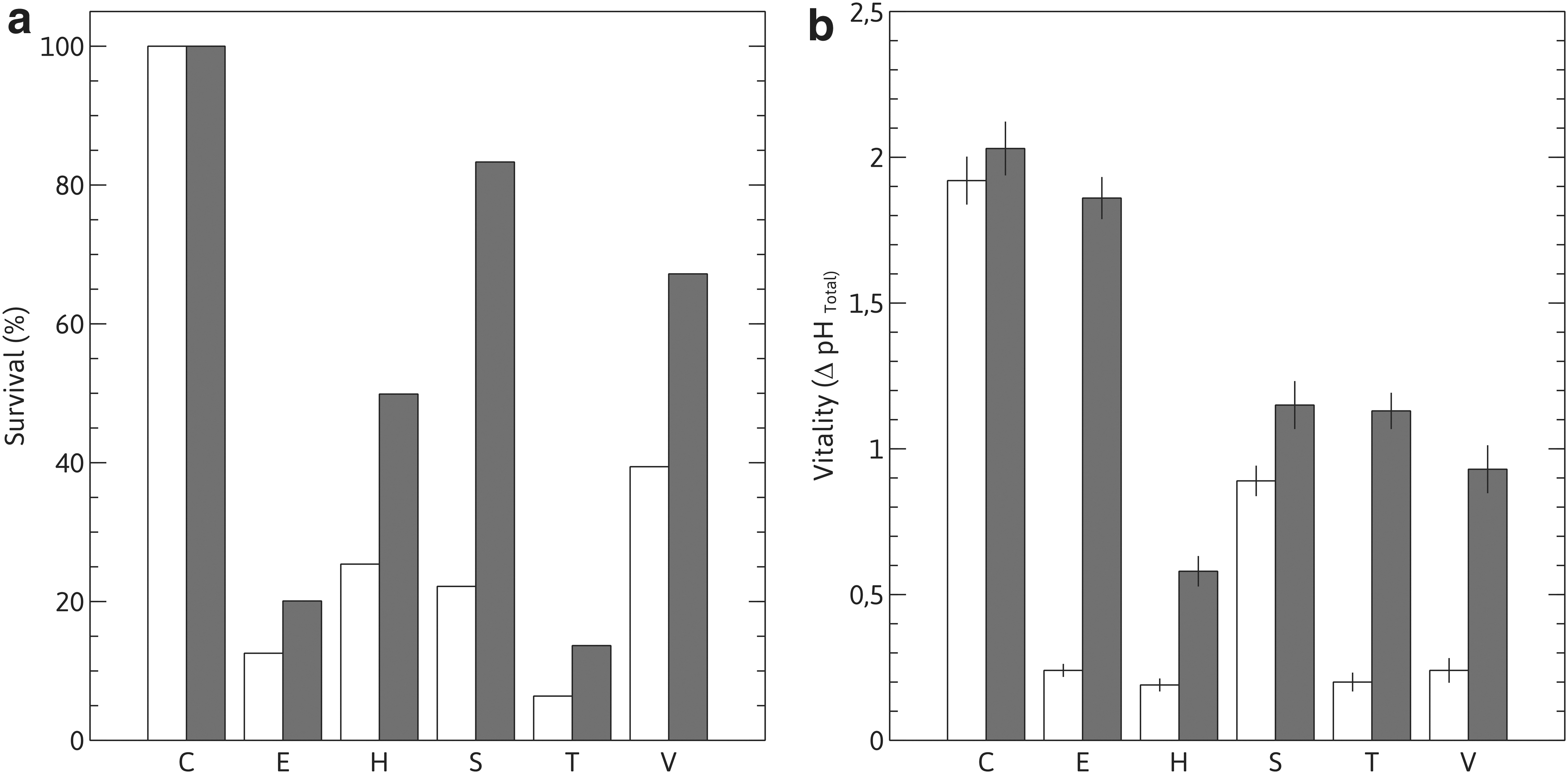
Viability and vitality under stress conditions.
Preconditioning of HAJ by exposure to toxic compounds present in JCH seemed not to affect its metabolic performance, since vitality control values were similar in magnitude to those of Y-7124 (Fig. 1b). After stress treatment, the difference between strains became more pronounced, with HAJ exhibiting significantly higher vitality than Y-7124 under the same conditions, and most pronounced for ethanol (∼8.0 fold higher).
HAJ exhibited significantly higher viability and vitality values than Y-7124 where stresses seemed to affect the fermentative performance more than its survival capacity. These results indicate that the adapted HAJ clone has a higher stress tolerance, evaluated as survival and mainly as metabolic robustness, than its isogenic parental strain. HAJ would thus be preferred to Y-7124 for ethanologenic fermentation, because the different stressful situations that yeasts could face during industrial cellulosic bioethanol production are less detrimental to its biocatalytic performance.
Oxidative Parameters: ROS and Antioxidative Enzymatic Activity
The stressful conditions applied produced an imbalance in the redox state of the cells, as depicted in Fig. 2a, where the intracellular ROS content of both strains are shown before and after being submitted to the different stresses assayed.
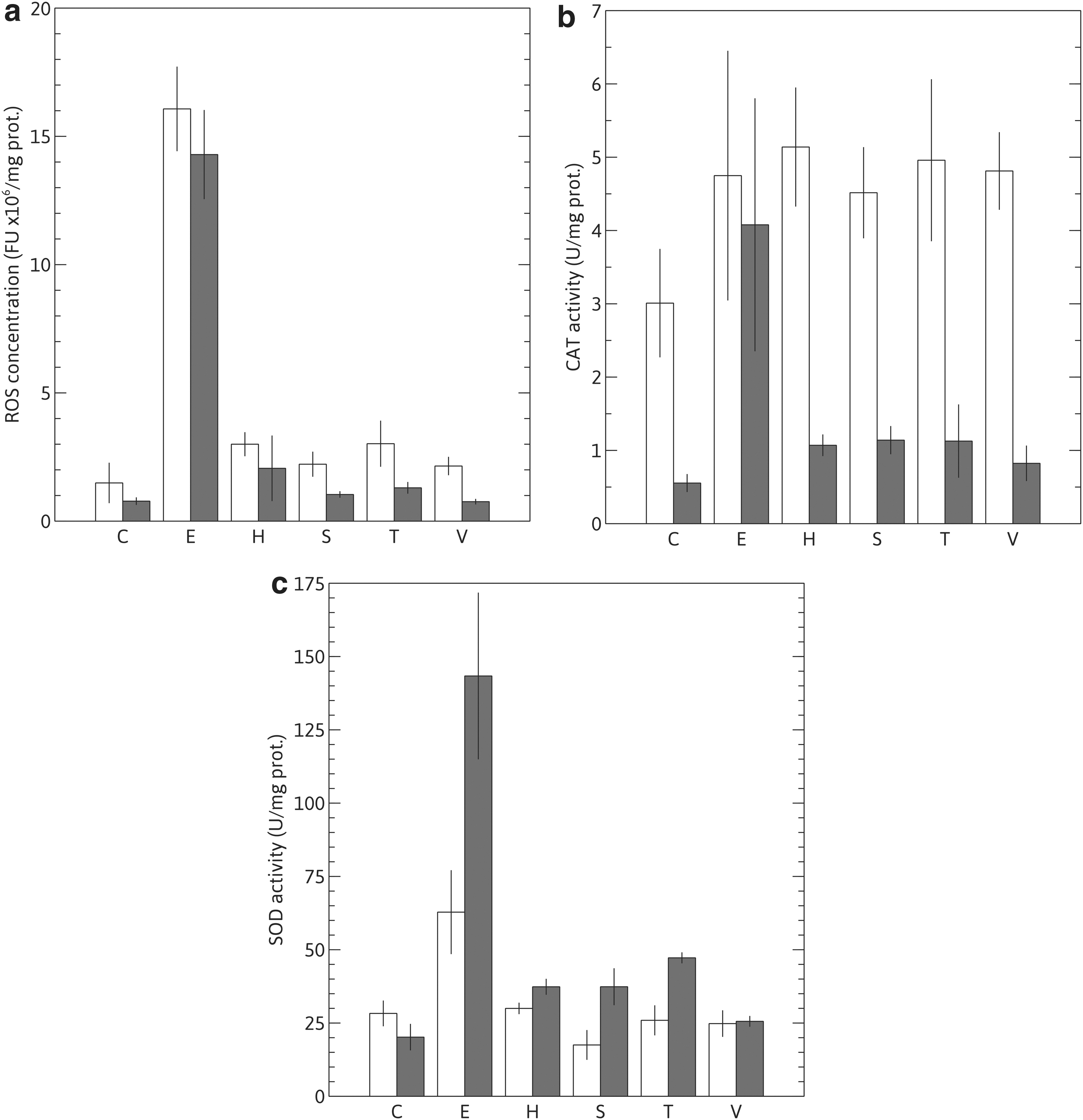
Oxidative parameters.
Upon exposition to the different stressors, ROS levels increased (Fig. 2a), in varying amounts depending on the stress applied and the yeast population being tested. While for Y-7124 intracellular ROS concentrations increased significantly after each treatment, the adapted clone ROS concentration increments were only statistically significant for ethanol, which was the most oxidative condition among those tested, with a 15-fold increase over control (non-stressed) levels (Fig. 2a). Ethanol, followed by temperature—the most damaging stressing conditions assayed (Fig. 1)—were the highest ROS-generating stresses assayed, even when compared to the highly oxidative treatment with H2O2. They were at the same time the most harmful ones, generating the lower viability and vitality values reported for both strains (mainly for Y-7124, Fig. 1). All the stresses assayed increased ROS intracellular levels in both yeast populations, supporting the idea that oxidative stress could be a cell-damaging mechanism common to all stresses tested. Compared with controls, ROS increase in HAJ after being subjected to each stress was 30–50% lower than in Y-7124. This indicates that the defenses against oxidative attacks may be expected to act more efficiently in the adapted clone, possibly because this strain has developed a more effective ROS-detoxifying mechanism than the one present in the parental strain. To evaluate this hypothesis, we measured intracellular CAT and SOD enzymatic activities in both strains and their relationship with intracellular ROS levels after being subjected to the different stress conditions.
As shown in Fig. 2b, CAT activity in HAJ was negatively affected by the stress conditions faced during its adaptation in JCH, since its activity in control cells (non-stressed) shows lower values than that of Y-7124 unstressed control. However, when both cell populations were submitted to stress, the relative increase in CAT enzymatic activity (compared to each control) was higher for the adapted strain. This was most notable for the more severe oxidative (ROS-generating) stresses assayed, i.e., ethanol (>7-fold increase), sorbitol and temperature (∼2-fold increase).
Concurrently with the observed values for CAT, SOD activity increase was more pronounced in HAJ, particularly after temperature and hydric stress treatments, e.g., ethanol and sorbitol, which were at the same time oxidative. As shown in Fig. 2c, SOD activity increased significantly in HAJ for all stresses, with this increase being higher with ethanol (∼7 fold), temperature, and sorbitol (∼2–3 fold). For Y-7124, only exposure to ethanol, the most oxidative stress assayed, seemed to elicit a rise in SOD levels (a 2-fold increase).
The increase of the enzymatic activities in HAJ does not elicit a complete reversal of the homeostatic redox state to the initial one. It does allow the adapted clone to withstand the increased levels of ROS, as shown by its higher values of viability and vitality compared to Y-7124, after both were challenged with the same stresses.
In this sense, our results are consistent with the principle of stress co-tolerance reported in Saccharomyces cerevisiae by different authors. 14,36,37 The improved ability of the adapted clone to resist harsh conditions caused by various stresses could be associated with its capacity for ROS detoxification, concurring with previous reports in S. cerevisiae. 14,20,22
Trehalose and Glycogen Levels
To evaluate other biochemical mechanisms reported to be involved in stress defense in yeasts,
48
glycogen and trehalose intracellular levels were determined in S. stipitis Y-7124 and HAJ. As can be seen in Fig. 3, the non-stressed adapted clone showed glycogen levels higher, although not significantly different, than Y-7124

Trehalose and glycogen levels.
Pyruvate Kinase Activity and Intracellular Oxidoreductase Coenzymes Levels
For Y-7124 control extracts, the following values were found (in μmol/g cell dry weight): 0.43/3.5 for NADH/NAD and 0.07/0.46 for NADPH/NADP. Similar values (∼ +/-5% difference) were found in HAJ control extracts. PK activity remained at ∼4.40 ± 0.18 mU/mL for both strains. No significant differences in those values were found after ethanol exposure (the most oxidative stress reported here) for either the parental strain or the adapted clone. These results suggest that the significant increase in intracellular trehalose concentration observed after stress treatment (see previous section) did not have a significant impact on the glycolytic pathway of the strains used.
Fermentation Performance in JCH
The adapted clone HAJ and the parental strain Y-7124 were compared for ethanol production in non-detoxified and non-supplemented JCH and YPDX semisynthetic medium. As shown in Table 1 and Fig. 4, the HAJ clone was well adapted to the hydrolysate fermentation medium, as shown by the kinetic parameters, exhibiting volumetric production, sugar consumption, volumetric productivity, yield values and kinetic parameters up to 3-fold higher than those of Y-7124, despite higher sugar consumption by the latter. However, bioethanol production efficiency of the parental strain in the control medium YPDX remained slightly higher than that of the adapted clone (93.02% vs 87.47% efficiency, for Y-7124 and HAJ strains, respectively).
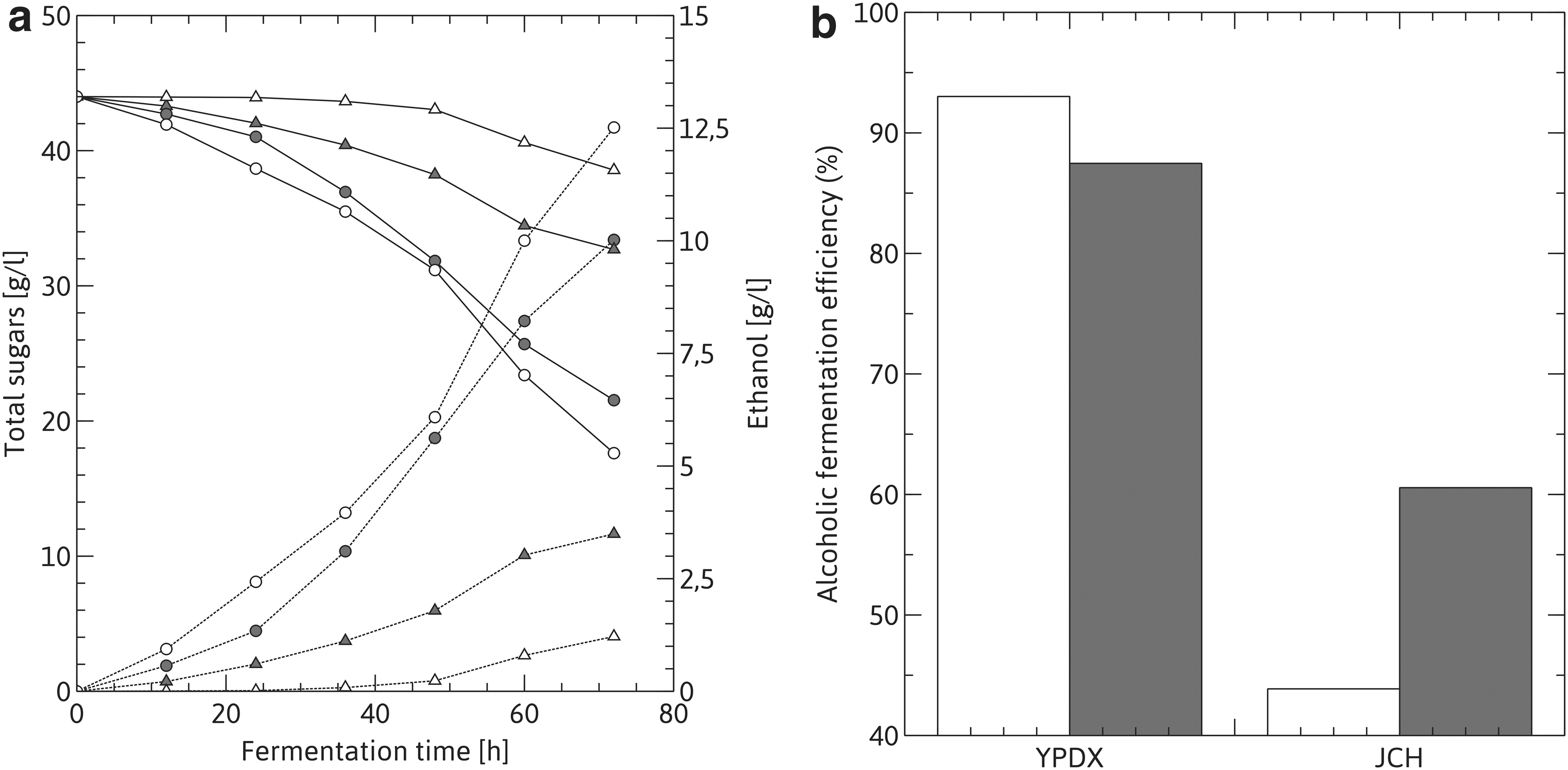
Fermentation performance in jojoba-cake hydrolysate.
Fermentation Parameters for Ethanol Production Using the Parental Strain S. stipitis NRRL Y-7124 and Adapted Clone HAJ
Y-7124: parental strain; HAJ: adapted clone; YPDX: medium containing yeast extract, peptone, dextrose and xylose; JCH: jojoba cake hydrolysate; Yp/s : Ethanol yield coefficient based on total sugar consumption (gp/g s ); Qp av: Average volumetric productivity determined as the average value of ethanol produced in function of fermentation time (gp/L·h); Qp max: Maximum volumetric productivity determined as the maximum value of ethanol produced in function of fermentation time (gp/L·h); Ethanologenic efficiency (%): sugar conversion efficiency (i.e., theoretical yield), calculated as ethanol yield/0.51) x 100. See Materials and Methods for details.
These results show that the adapted clone exhibited higher ethanologenic efficiency compared to the parental strain when using a non-supplemented and non-detoxified lignocellulosic hydrolysate as a fermentation medium. Its greater stress tolerance after adaptation would thus allow it to obtain higher ethanol production levels in such hydrolysates, turning second-generation industrial bioethanol production more economically viable by saving time and costs of medium detoxification.
Conclusions
The robustness of the evolutionarily adapted strain of S. stipitis presented in this work was associated with its higher capacity (compared to its parental isogenic strain) to tolerate the different oxidative (ROS-generating) stresses found during a standard industrial bioprocess of cellulosic ethanol production. This fact is indicative that there might be a common basis for the mechanism of stress damage and for its repair. Stress tolerance was associated, in turn, to the activation of the antioxidant enzymes catalase and superoxide dismutase. The latter's activation is quantitatively related to the level of oxidative stress applied, indicating that O2 •− plays an important role in the oxidative stress of this strain.
Cell membrane-protecting molecules, like trehalose, increased their concentration in response to the mildly-stressing adaptive process, increasing even further after more severe stress challenges. The highest intracellular trehalose levels in the adapted strain were reached under osmotic stress, indicative of the specificity between the type of stress applied and the concentration of that disaccharide.
As a whole, it can be concluded that a stress-tolerant yeast strain used in cellulosic ethanol production has to present high antioxidative defenses, and that attaining high intracellular levels of cell membrane protectants like trehalose is a goal to reach if a robust strain is desired. Our results also show that the evolutionarily adapted strain of S. stipitis could be used industrially to efficiently produce cellulosic ethanol from a non-detoxified agro-industrial fermentation medium. This is mainly due to its increased tolerance to osmotic and ethanol stresses, the two main stressful conditions that prevent the industrial use of this yeast species in ethanologenic fermentations.
Footnotes
Acknowledgments
The authors wish to thank Dr. D. Wassner for providing the jojoba cake, and Dr. Ma. T. Rimoldi for oxidoreductase coenzymes and pyruvate kinase activity measurement. We also thank Dr. Susana Passeron for the critical reading of the English manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from PIP 2012-2014 and by PIO CONICET-UNSAM 2015-2017 Argentina (Consejo Nacional de Investigaciones Científicas y Técnicas).