
Abstract
The maize seed expression system is a powerful platform for recombinant protein production. It has been utilized for high-yield production of industrial cellulases such as endo-1,4-β-D-glucanase (E1) and 1,4-β-D-glucan cellobiohydrolases (CBHI & CBHII). However, inhibitory compounds that could significantly reduce the cellulolytic efficiency were detected even in purified E1 and CBHII samples. After several steps of purification and analysis, the inhibitory compound found in the purified E1 sample had characteristics consistent with a thermostable polysaccharide with a molecular size over 10 kDa. Further polysaccharide analysis confirmed the molecular size of the detected polysaccharide and suggested that it is an arabinoxylan with a high arabinose/xylose ratio. Even though the strict column purification process should have removed water-soluble non-protein molecules during protein purification, the inhibitor remained. A possible explanation is that recombinant cellulase can bind to the inhibitor, carry it through all purification steps and finally release it when denatured by heat.
Introduction
Transgenic maize seed is a high-performance expression system for recombinant cellulase. Recombinant cellulases, including endo-1,4-β-D-glucanase (E1), originally from Acidothermus cellulolyticus (GenBank Accession #U33212); 1,4-β-D-glucan cellobiohydrolase I (CBHI) originally from Trichoderma reesei (GenBank Accession #X69976); and 1,4-β-D-glucan cellobiohydrolase II (CBHII) also originally from Trichoderma reesei (GenBank Accession #M16190), have been produced in transgenic maize seeds for industrial purposes. 1,2 To make these expressed recombinant cellulases into commercial products, a highly effective purification process, including buffer crude extraction, ammonium sulfate (AS) precipitation and ion-exchange chromatography designed by Hood et al., 3 has been applied.
Despite this, inhibitory compounds were found in the recombinant protein production system. According to unpublished data by Hood and Davis, an inhibitor of cellulase was found in the crude extract of recombinant E1 enzyme. Although AS precipitation was able to remove some of the inhibitor, inhibition of cellulase still remained in the final purified protein fractions. Based on literature that reported polysaccharides such as xylans are inhibitors of cellulase, 4 –7 and on clues gathered from previous unpublished work, water-soluble macromolecular compounds were believed to be the main inhibitors. Therefore, the goal of this work was to identify the inhibitory compound and find answers as to why this cellulase inhibitor would remain throughout the entire protein purification process.
In this research, we investigated fractions derived from purified recombinant E1 and CBHII. The results strongly suggest that an arabinoxylan with a high Ara/Xyl ratio is the potential inhibitor of cellulase and that it binds to the maize-seed produced recombinant cellulase and is carried to the final purified protein fractions.
Materials and Methods
Materials and Chemicals
Transgenic maize seeds producing either recombinant E1 or CBHII 1,2 were used for recombinant E1 and CBHII extraction and purification. Xylanase 1 (Sigma-Aldrich, X3254, St. Louis, MO) and Proteinase K (New England Biolabs, P8107S, Ipswich, MA) were applied for inhibitory sample treatment. Commercial cellulase cocktail (Sigma-Aldrich, C2730) was utilized for the cellulolytic efficiency assay. Rye arabinoxylan (Megazyme, P-RAXY, Bray, Ireland) was tested as a control for the identified maize arabinoxylan.
Preparation of Inhibitory Samples
Inhibitory samples were derived from the purification process of either recombinant E1 or CBHII from transgenic maize seeds (Fig. 1A). The purification method was based on the protocol described by Hood et al: 3 a 500 g sample of transgenic seeds went through grinding, 0.05 M sodium acetate buffer, pH 5.0, extraction and 70% AS precipitation. The AS-precipitated protein was separated by filtering through a paper filter. The precipitated protein was re-dissolved in 0.05 M sodium acetate buffer, pH 4.0, and desalted through a 10 kDa molecular weight cut off biomax membrane (polyethersulfone) fixed in a tangential flow filtration (TFF, Millipore, Billerica, MA) system. Subsequent cation-exchange and anion-exchange column chromatography were applied for further protein purification, as described by Hood et al. 3 After these steps, final purified recombinant E1 and CBHII (16.0 and 11.5 mg/mL of total soluble protein concentration respectively) were prepared in 0.05 M sodium acetate buffer, pH 5.0 with 20% glycerol and stored at −20°C.
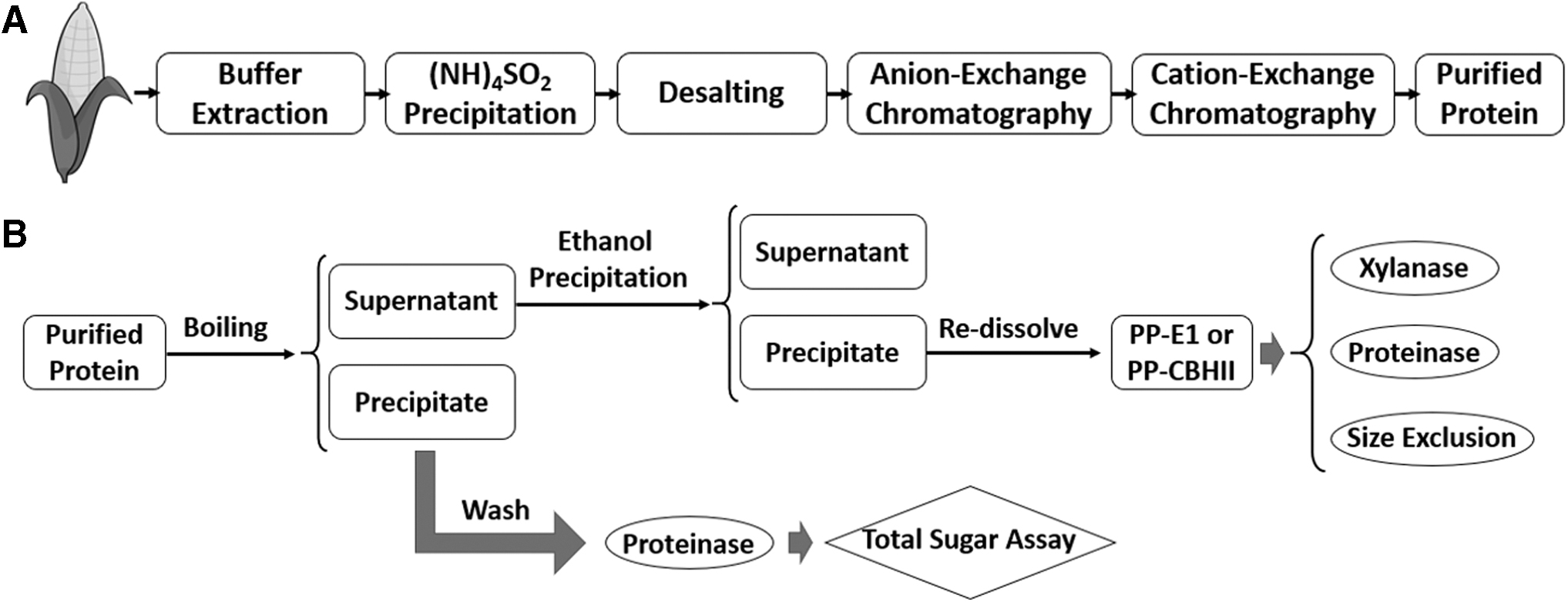
The final purified recombinant cellulase was investigated for inhibitory compounds. A 1 mL aliquot of purified recombinant E1 or CBHII was incubated in boiling water for 30 min and centrifuged at 13,000 relative centrifugal force (rcf) for 10 min. The supernatant was collected and precipitated with 80% ethanol at room temperature for 20 min. After centrifuging at 13,000 rcf for 5 min, the pellet was washed twice with the same volume of 80% ethanol and dried on a heating block at 90°C for 30 min. The dried pellets derived from 1 mL of purified cellulase stock were re-dissolved in 0.5 mL double distilled water as the purified precipitate sample (PP-E1 and PP-CBHII; Fig. 1B).
Further Treatment of Inhibitory Samples
Further treatments including xylanase, proteinase and size-exclusion fractionation were applied to PP-E1 (Fig. 1B). In the xylanase treatment, a 0.2 μL/mL (v/v) of purchased xylanase was applied using the recommended conditions (70°C, pH 5.0, 24 h) to eliminate potential xylan. After incubation, the solution was incubated in boiling water for 30 min to denature the applied xylanase. In the proteinase treatment, proteinase K with a final concentration of 0.1 μg/μL was applied to samples in 0.05 M sodium acetate buffer, pH 5.0, containing 5 mM CaCl2 in a 2-mL Eppendorf tube with 2 glass beads (0.5 mm) and was incubated at 37°C with 180 rpm shaking for 24 h. After incubation, the solution was boiled for 30 min to denature the applied proteinase. In the size-exclusion fractionation, samples were centrifuged at 13,000 rcf to pass sequentially through a 50 kDa and a 10 kDa size-cutoff Corning® Spin-X® UF Centrifugal Concentrator PES membrane (Corning®, 431480 and 431478, Corning, NY). The resulting size-exclusion fractions (>50 kDa and 10–50 kDa) were reconstituted with 0.05 M sodium acetate buffer, pH 5.0, to their initial volume.
Proteinase treatment was applied to boiling-denatured precipitates of purified recombinant E1 and CBHII as well (Fig. 1B). The pellet of denatured recombinant cellulase from 1 mL final purified fraction was collected and washed three times with 1 mL washing buffer (0.05 M sodium acetate buffer, pH 5.0, containing 5 mM CaCl2). The washing buffer from the third wash was collected. The washed pellet was resuspended in 1 mL washing buffer and underwent the same proteinase K treatment described above. After the proteinase K treatment, the entire solution was boiled for 30 min to denature the proteinase K and prepared for a total sugar assay and a cellulase efficiency assay.
Total Sugar Assay
A phenol-sulfuric acid total sugar assay 8 as a glucose equivalent was applied to quantify polysaccharides in the target sample. A 30-μL aliquot of sample with appropriate dilution was well mixed with 30 μL 5% phenol solution in a 96-well plate. A 120 μL aliquot of 98% sulfuric acid was added into the sample-phenol mixture followed by immediate stirring. After incubation at room temperature for 20 min, the 96-well plate was read in a Synergy HT microplate reader (BioTek, Winooski, VT) at 490 nm. A standard curve made of a standard glucose solution (concentration ranges from 0.0 to 3.0 mg/mL) was used for quantification.
Polysaccharide Analysis
The PP-E1 sample was condensed through a 10 kDa size-cutoff Corning® Spin-X® UF Centrifugal Concentrator PES membrane to a final total sugar concentration of 20 mg/mL. A 300-μL sample of condensed PP-E1 was sent to the Complex Carbohydrate Research Center at the University of Georgia for glycosyl composition, linkage analyses, and molecular weight determination.
Cellulase Efficiency Assay
The cellulose-DNS (3,5-Dinitrosalicylic acid) assay was utilized to evaluate the activity and inhibitory effect of the derived samples. The enzymatic efficiency (ηe) in the cellulose-DNS assay was represented by:

In the cellulose-DNS assay, a reaction mixture with a total volume of 200 μL containing 1.0 mg of Sigmacell cellulose (Sigma-Aldrich, S3504) and 60.0 μg commercial cellulase cocktail (Sigma-Aldrich, C2730) was prepared with 0.05M sodium acetate buffer, pH 5.0, and incubated in a sealed 96-well plate with 1 glass bead at 50°C for 24 h with 100 rpm reciprocal shaking. Potential inhibitory samples were tested in this reaction system. After the reaction was complete, a 50 μL aliquot of reacted solution was collected for the DNS assay. 9
Since the inhibitory samples possibly contained reducing compounds that would be detected by the DNS assay or may be digested by cellulases, an additional cellulose-free control (substrate-free control) group was set up to account for the background absorbance.
Statistical Analysis
Statistical analysis was applied to the cellulase efficiency assay. Experiments were performed with at least triplicates for the efficiency assay. All data were averaged with experimental standard error. A double tailed t-Test was applied for statistical significance with p < 0.05.
Results and Discussion
Inhibition Exhibited by the Product of the Recombinant E1 Purified Protein Implied an Inhibition Model based on Partially Denatured E1
Inhibitor samples derived from both purified recombinant E1 (PP-E1) and purified recombinant CBHII (PP-CBHII) were investigated. When introduced into cellulolytic reactions, PP-E1 showed significant inhibition of the commercial cellulase cocktail. A dosage test of PP-E1 was conducted and indicated that a low concentration of 5% (v/v) in the reaction system reduced cellulolytic efficiency to 71% (Fig. 2A). However, 10% (v/v) of PP-CBHII did not show any significant inhibition (Fig. 2B). Since PP-E1 was derived from final purified E1 fractions, it appeared that the inhibitory compound is protein and unique to the E1 transgenic maize line. Considering our highly effective recombinant protein purification system and the limited difference in final protein concentration between the E1 and CBHII enzymes, it was hypothesized that the inhibition of cellulase might be attributed to properties of recombinant E1.
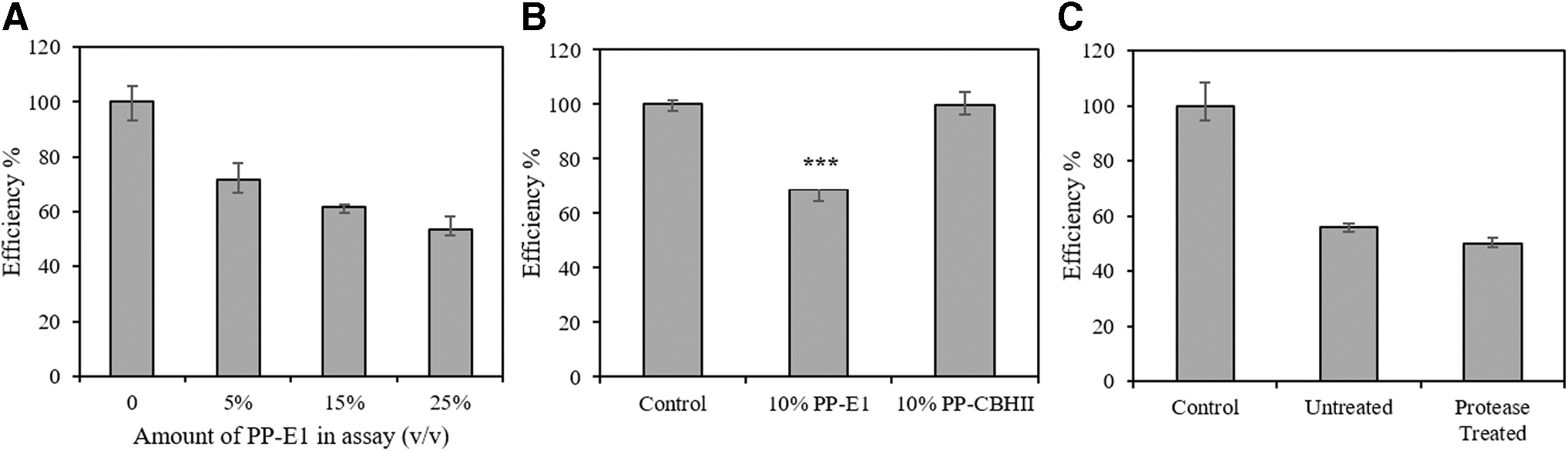
Inhibitory effect of PP-E1 and PP-CBHII on commercial cellulase cocktail.
Endoglucanase E1 is a thermostable enzyme with an optimal reaction temperature of 80°C. 10 In addition, as an endoglucanase, E1 can randomly bind to and hydrolyze glycoside linkages within (rather than at the ends of) cellulose chains (Fig. 3A). 11 With these two special properties, this heat-treated recombinant E1 in our case may show some possibility of becoming an inhibitor of cellulases. After boiling, as a thermostable enzyme, E1 may retain partial function or partially renature at room temperature like other thermostable proteins. 12 Hence, the possibility exists that after boiling, some recombinant E1 was partially denatured but remained soluble and could bind to cellulose without any cellulolytic activity. When bound to a cellulose fiber, partially denatured E1 would occupy reaction sites on the cellulose fiber and thus prevent the interaction between active cellulases and cellulose (Fig. 3B). CBHII, unlike E1, will be thoroughly denatured by heat. Even if some CBHII molecules survived boiling or were only partially denatured by boiling, because CBHII can only bind to and hydrolyze at non-reducing ends of a cellulose fiber, 13 they will not block the activity of other cellulases on cellulose (Fig. 3C).
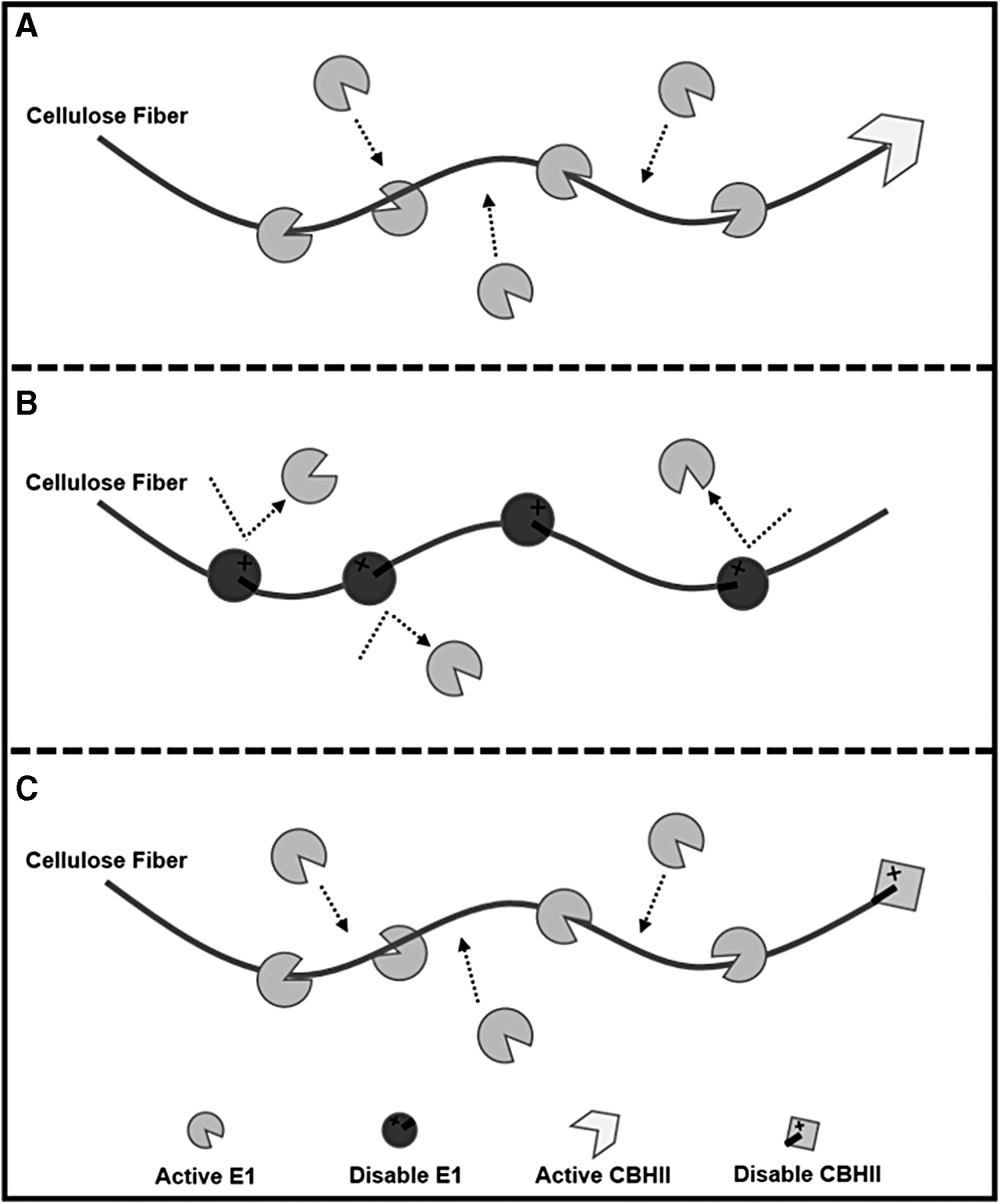
A hypothesized inhibition model of partially denatured recombinant E1.
Based on this inference, we hypothesized that the partially denatured E1 enzyme derived from boiling treatment could block the binding of cellulases to cellulose fibers and therefore inhibit cellulase activity. In order to test this hypothesis, proteinase K treatment was applied to PP-E1. Proteinase K was expected to thoroughly hydrolyze any protein including the partially denatured E1. However, after 24 h of treatment with proteinase K at 37°C, treated PP-E1 at 15% (v/v) still retained significant inhibition of the commercial cellulase cocktail compared to the untreated control, and the proteinase K treatment did not rescue the cellulolytic efficiency or make any difference in the inhibition level (Fig. 2C). This retained inhibition of proteinase K-treated PP-E1 disproved our hypothesis and induced us to consider alternatives.
Characterization of Inhibitory Compounds Found in Purified Recombinant E1
Even though our hypothesis on the inhibition of partially denatured E1 was wrong, the existence of inhibitory compounds in the purified E1 enzyme fraction was confirmed. Because polysaccharides were suspected, a phenol-sulfuric acid total sugar assay was performed. 8 Surprisingly, fraction PP-E1 was found to contain 10.6 mg/mL total sugar as glucose equivalent.
To estimate the molecular size of these discovered polysaccharides, PP-E1 was fractionated sequentially through 50 kDa and 10 kDa size-cutoff spin columns. Large (>50 kDa) and medium (10 ∼ 50 kDa) fractions were brought back to their original volume with double distilled water. Total sugar of each size exclusion fraction was estimated by the phenol-sulfuric acid method (Fig. 4A). Among the three size-exclusion fractions, the large and medium fractions contained about 3.7 mg/mL and 3.6 mg/mL of total sugar, respectively, while the small fraction (<10 kDa) contained almost no sugar. The polysaccharides in PP-E1 are likely a mixture and their molecular sizes varied because of their permeability to the different size-exclusion membranes. The inhibitory effect of each size-exclusion fraction on the commercial cellulase cocktail was evaluated in a cellulose-DNS assay. Both large and medium fractions with a concentration of 20% (v/v) showed significant inhibition on the commercial cellulase cocktail, while the small fraction did not show any inhibition (Fig. 4B).
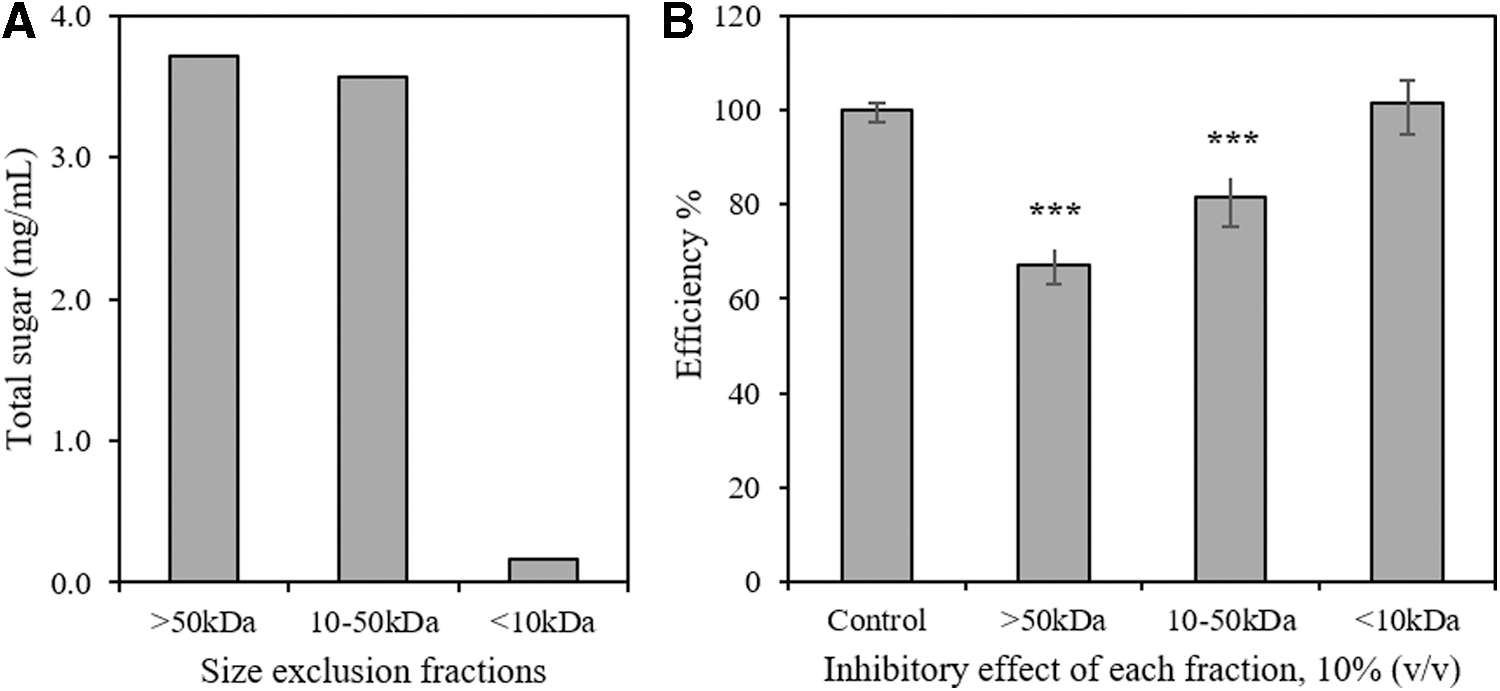
Size exclusion fractions of PP-E1 and their inhibitory effect in the cellulolytic efficiency assay.
Identification of Inhibitory Polysaccharide Based on Polysaccharide Analysis
Although we speculated that starch, pectin or xylan could be the inhibitor, the inhibitor was resistant to amylase, pectinase, and xylanase (data not shown). Therefore, to characterize the inhibitory polysaccharide, the PP-E1 fraction was sent to the Complex Carbohydrate Research Center at the University of Georgia for glycosyl composition analysis, glycosyl linkage analysis, and molecular weight determination.
In molecular weight determination, size-exclusion chromatography of the PP-E1 fraction (Fig. 5) showed the presence of several overlapping peaks at positions corresponding to dextran molecular masses of approximately 500, 100, 40 and 10 kDa (arrows). The major peak with the highest concentration was located at 40 kDa. This result is consistent with our estimation based on 50 kDa and 10 kDa size-cutoff column fractionation.
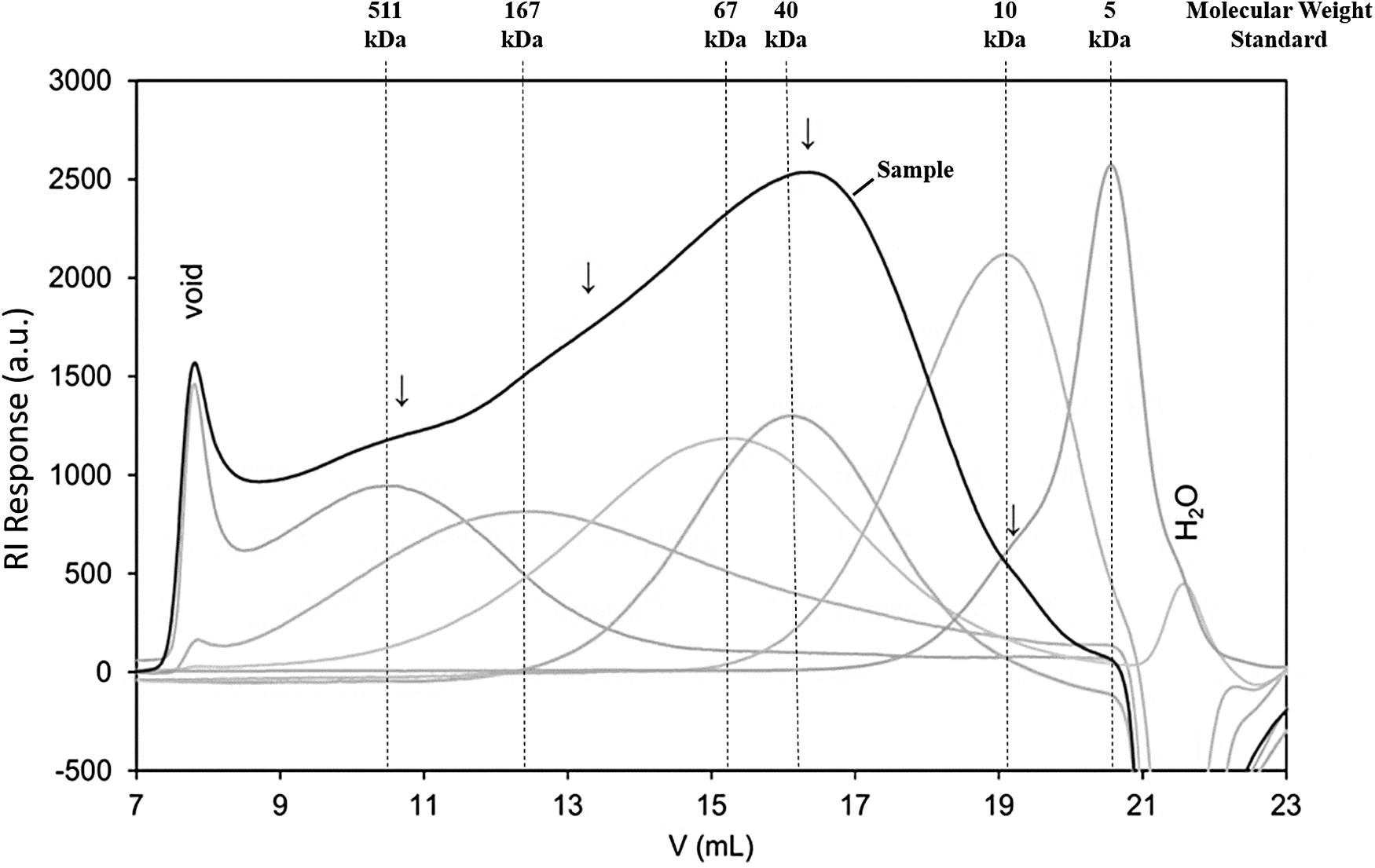
Overlay of Superose 6 chromatograms for the PP-E1 fraction and dextran molecular weight standards. Sixty μg of protein sample and 40 μg of each molecular weight standard were loaded onto a size exclusion column. Eluted fractions were monitored by a refractive index detector. The signal response intensity of each eluted fraction was presented in absorbance units (a.u.). Arrows indicate approximate peak maxima for the sample.
In the glycosyl composition assay, PP-E1 contains mainly arabinose and xylose in approximately equimolar amounts (Ara/Xyl ratio of 1.09) with small amounts of fucose, mannose, galactose and glucose (Table 1). Together with the data of glycosyl linkage analysis (Table 2 and Supplementary Fig. S1), it can be inferred that the inhibitory polysaccharide might be arabinoxylan. According to glycosyl linkage analysis, about 82.6% of arabinoses serve as terminal substitutes, while about 61.8% of xyloses serve as singly or doubly branched residues. Therefore, a total arabinosyl substitution on the xylan backbone is at least 70%, meaning that hypothetically 70% of xylopyranosyl residues hold at least one arabinofuranosyl or arabinopyranosyl substitution. Because polysaccharide sequencing data were not collected, a detailed structure of the detected arabinoxylan cannot be determined.
Glycosyl Composition of PP-E1 Fraction
Relative Molar Percentages of Linkages Detected in the PP-E1 Fraction
Arabinoxylan is a major component of hemicellulose that has a xylan backbone with arabinose side chains. As a major component of dietary fiber, it is found in many cereal grains especially in endosperm. 14 –16 The arabinoxylan detected was obtained from the maize seed expression system. However, considering that the commercial cellulase cocktail used in these experiments (Sigma C2730) has hydrolytic activity on xylan, 17 –19 it was still a mystery how an arabinoxylan with a digestible xylan backbone could maintain the inhibition on cellulases involved in this study.
The High ARA/XYL Ratio is Likely the Source of Inhibition
After determining that the inhibitory compound is likely maize-derived arabinoxylan, we tried to eliminate it with commercial xylanase (Sigma-Aldrich, X3254). A commercial rye arabinoxylan (Ara/Xyl ratio: 0.61) was used as positive control. Unfortunately, xylanase treatment was not able to hydrolyze this maize-derived arabinoxylan or to relieve its inhibition in the cellulose-DNS assay (Fig. 6A). In addition, compared to native maize arabinoxylan, rye arabinoxylan did not show any inhibition on the commercial cellulase cocktail and could be easily digested by the commercial xylanase (Fig. 6B). The poor effect of xylanase treatment on maize-derived arabinoxylan compared to rye arabinoxylan is likely due to the high Ara/Xyl ratio.
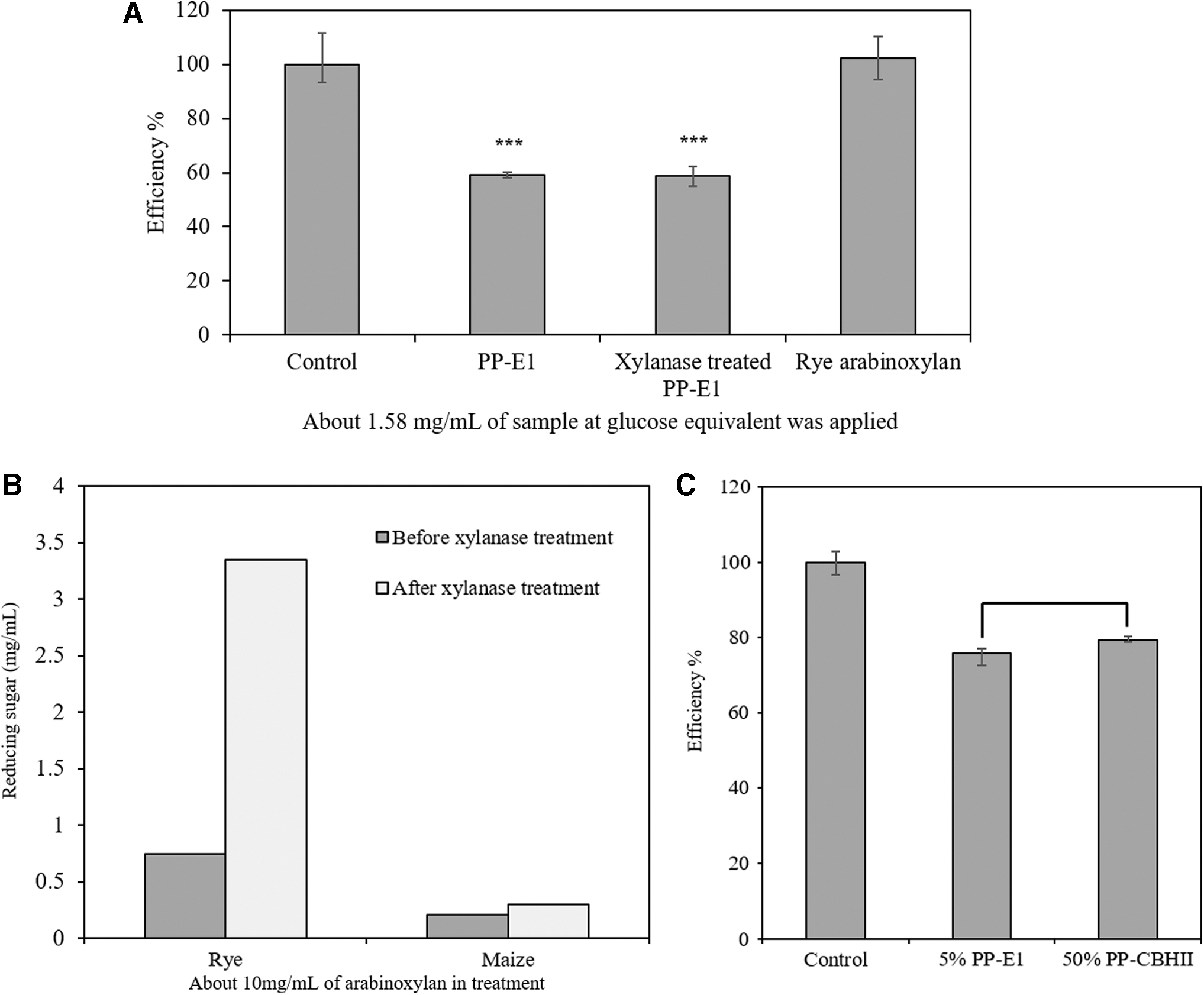
(A) inhibitory effect of xylanase treated PP-E1 on commercial cellulase cocktail compared with untreated PP-E1 and commercial rye arabinoxylan;
From the polysaccharide analysis, it was interesting to observe a high Ara/Xyl ratio (1.09) in our inhibitory fraction. The Ara/Xyl ratio varies a lot among species. In maize, especially maize seed products such as corn bran and corn fiber, the Ara/Xyl ratio of extractable arabinoxylan is usually under 1.00 (may range from 0.50 to 0.80). 20 –22 Since it has been reported that the Ara/Xyl ratio is negatively correlated with the percentage of arabinoxylan removed by xylanase treatment, 24,25 it makes sense that such a high Ara/Xyl ratio of our inhibitory arabinoxylan may contribute to its resistance to the xylanase in our study. At this point, it is quite likely that with a relatively high Ara/Xyl ratio, maize-derived arabinoxylan (Ara/Xyl ratio: 1.09) acts as the inhibitor of cellulases.
Even though the Ara/Xyl ratio has been discussed in relation to enzymatic resistance, 24,25 in order to explain the exact mechanism of inhibition, other factors including branching level, molecular size and substitution of other functional groups must be considered.
Polysaccharide Found in PP-CBHII
If the cellulase inhibitor detected in PP-E1 is a native maize arabinoxylan, then it should be contained in CBHII transgenic maize lines as well. Total polysaccharides with a concentration of 1.07 mg/mL in PP-CBHII were detected by a total sugar assay. Although in the final purified recombinant cellulase sample, total soluble protein content of E1 is about 1.5-fold higher than CBHII (16.5 mg/mL of total soluble protein in purified E1 and 11.5 mg/mL of total soluble protein in purified CBHII), the total sugar content of E1 is about 10-fold higher than CBHII (10.6 mg/mL of total sugar in PP-E1 and 1.07 mg/mL of total sugar in PP-CBHII) (Fig. S2). To determine whether the polysaccharide detected in PP-CBHII has an inhibitory effect on cellulase as well, the same polysaccharide amount (0.25 mg/mL total sugar) of PP-E1 and PP-CBHII were compared in the cellulolytic efficiency assay. These two samples at the same level of total sugar displayed the same level of inhibition (75% decreased efficiency for PP-E1; 79% decreased efficiency for PP-CBHII; p < 0.05) on a commercial cellulase cocktail (Fig. 6C). This result implies that the polysaccharide detected in PP-E1 and PP-CBHII may be the same inhibitory arabinoxylan.
Further Tests to Explain the Presence of Inhibitor Polysaccharide in Purified Recombinant Cellulase Fractions
It is still a mystery how polysaccharide could be present in the protein sample derived from a specific protein purification process which included buffer extraction, ammonium sulfate precipitation, 10 kDa size-cutoff filtration, cation exchange chromatography and anion exchange chromatography. One explanation is that sufficient amounts of this polysaccharide that could inhibit the enzyme still remained after the applied protein purification process. However, the fact that no polysaccharide was detected in the flow through fractions prior to elution in both cation- and anion- exchange chromatography (Fig. S3) refuted this explanation.
Efforts were made to detect the inhibitory polysaccharide remaining in the denatured recombinant proteins as well. During the preparation of purified fractions PP-E1 and PP-CBHII, recombinant E1 and CBHII were denatured by boiling. Denatured E1 and CBHII as the precipitate were collected. After multiple washes with sodium acetate buffer (to eliminate remaining free polysaccharides generated during boiling), precipitated cellulases were digested with proteinase K. The protein solution after proteinase K digestion was condensed and washed through 10 kDa size-cutoff spin columns and assayed for total sugar content. Very limited amounts of total sugar were detected from both digested E1 and CBHII. In digested E1, about 0.17 mg/mL total sugar was detected whereas in digested CBHII, about 0.60 mg/mL total sugar was detected (Fig. 7A). In addition, polysaccharides detected in both denatured E1 and CBHII performed significant inhibition on commercial cellulase cocktail at the same concentration of about 0.25 mg/mL (Fig. 7B) and were proven to be the same inhibitory compounds as detected in PP-E1 and PP-CBHII.
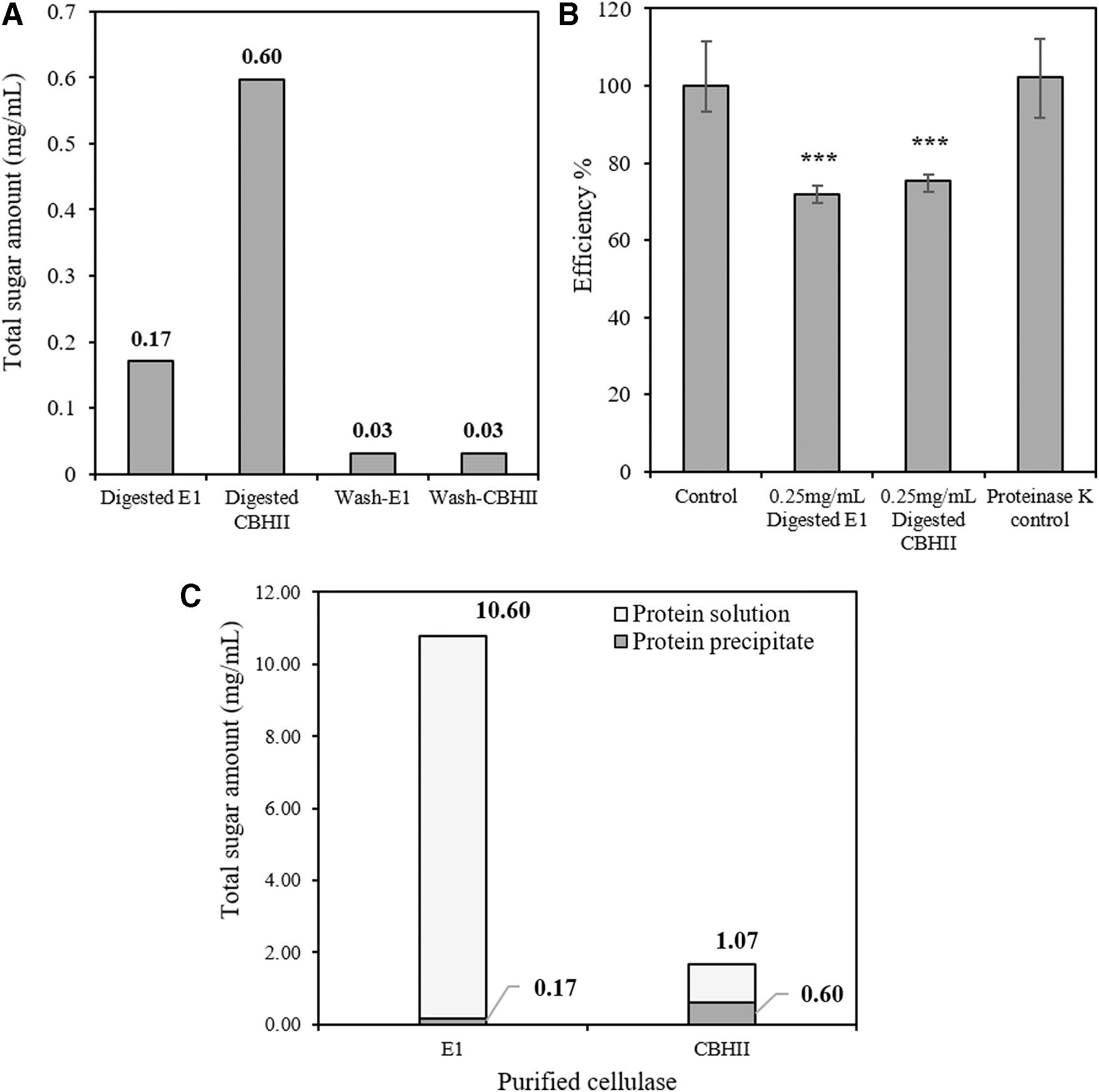
Detection of inhibitory polysaccharides in proteinase k digested recombinant E1 and CBHII.
If the inhibitory polysaccharides presented in free form, adequate washing following boiling treatment should eliminate them and both denatured E1 and CBHII precipitates should be inhibitor-free. In contrast, if the inhibitory polysaccharides presented in an E1 or CBHII-bound form, then it makes sense that some of it remained bound to the recombinant E1 and CBHII even after boiling denaturation. From this point, it can be concluded that purified recombinant E1 contains 6.4-fold more total sugar than purified recombinant CBHII and released about 98.4% of total sugar after boiling treatment, whereas purified recombinant CBHII released only 64.2% (Fig. 7C). However, the difference in the ratio of protein content to bound polysaccharide and the difference in polysaccharide releasing rate between recombinant E1 and CBHII remain unexplained. Maybe these differences can be attributed to protein properties such as spatial conformation, molecular size, charge level and hydrophilicity. The difference in the amount of carried polysaccharide between recombinant E1 and CBHII might also be attributed to the parent inbred lines of maize to which the transgenic lines were bred. E1 is in a germplasm background of SP114 and SP122 from Holden Foundation seeds (now Monsanto/Bayer) whereas the CBHII is a background of Stine Seeds (Adel, IA). The grain from these diverse lines could likely have widely variable arabinoxylan content.
Arabinoxylans, the putative inhibitor of cellulase in this study, are major components of hemicellulose in cell walls, especially in endosperm. 14,15 Recombinant CBHII was targeted to be expressed in maize endosperm cells and deposited in the cell wall matrix. Thus, there is no doubt that recombinant CBHII could reach and carry the inhibitory arabinoxylans. For recombinant E1, although it was expressed in maize germ and targeted to the cell vacuole, during maize seed grinding and buffer extraction, recombinant E1 was presented the chance to interact with the inhibitory arabinoxylans located in cell walls and bring them to the final purified fraction. No matter how the inhibitory arabinoxylan encounters the recombinant proteins, the maize native inhibitory polysaccharide presents an interesting situation in utilizing maize seeds as the expression platform for commercial recombinant cellulase production. Clearly, the inbred lines producing the enzyme have an impact on inhibitor concentration and thus the method of formulation of the final product can minimize the inhibitor's effect.
Conclusion
Preliminary inhibition data suggested the existence of unknown compounds in maize grain that can inhibit commercial and recombinant cellulases. After significant investigation, thermostable inhibitory macromolecular compounds were found in the final purified fraction of maize seed-produced recombinant E1 and CBHII enzymes and were suggested to be arabinoxylan with a high Ara/Xyl ratio of about 1.09, which may explain why this inhibitory arabinoxylan is resistant to commercial xylanase and is inhibitory to cellulases. After further experiments, the data suggest that the inhibitory polysaccharides may bind to recombinant cellulases, be carried through the entire protein purification process, and be released after boiling treatment of recombinant cellulases.
Footnotes
Author Disclosure Statement
Hong Fang declares no competing interests. Elizabeth E. Hood is owner of Infinite Enzymes, LLC, which provided the corn lines analyzed in this study.
Funding Information
This work was supported by USDA-NIFA Program Grant (2015-70001-23423), the Arkansas Biosciences Institute, and the Chemical Sciences, Geosciences and Biosciences Division, Office of Basic Energy Sciences, U.S. Department of Energy grant (DE-SC0015662) to Parastoo Azadi at the Complex Carbohydrate Research Center.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.