
Abstract
Corncob is an agricultural byproduct, a result of maize grains processing. It is a lignocellulosic biomass, and its hydrolysis releases a great amount of xylose, a five-carbon sugar that is not readily assimilated by industrial yeasts for bioethanol production.
Scheffersomyces stipitis CBS 6054, however, is a yeast that can directly assimilate xylose. It is also known that aeration control has a positive effect on ethanol production during the fermentation process. However, it is rare to find research integrating the inoculum preparation with fermentation processes, and the physiological condition of the inoculum on the very moment of the culture transference can trigger off-target effects on fermentation performance. In this work, we evaluate the inoculum production of S. stipitis CBS 6054 in corncob hemicellulosic hydrolysate (CHH), containing glucose and xylose, and employing Erlenmeyer flasks with and without baffles. Baffles tend to cause an increase in oxygen transfer rate. The inoculum prepared on CHH using baffled flasks (BF) achieved faster exponential phase than unbaffled flasks (UF; 7–14 h versus 12–18 h). Cell density in this phase was also higher (0.2–2.8 versus 0.01–0.70 g L−1) compared with UF. Fermentations conducted with these inoculums presented a significant increase in ethanol production and yield (YP/S of 0.17 to 0.23), proving the important role of oxygen transfer rates on inoculum preparation.
Color images are available online
Introduction
In December 2015, the United Nations gathered 190 countries to discuss a potential agreement on global warming remediation. The relevant points raised during this conference created the Paris Agreement, where each country agreed to take steps to decrease greenhouse gas (GHG) emissions to narrow the global temperature increase to a maximum of 2°C until the end of this century, based on the Intergovernmental Panel on Climate Change (IPCC) report. 1,2 The International Energy Agency (IEA) modeled a sustainable development scenario following this thinking, based on biofuels contribution to mitigating the GHG emission. In this scenario, if in the next years the global biofuel production follows the historical pattern of development, by 2025 it will be necessary to almost double biofuel production to accomplish this goal. 3
In the biofuel scenario, Brazil stands out; it produced more than 30 billion L in 2019, second only to the United States in terms of volume. 4 Although first-generation ethanol production is a well-established process, second-generation ethanol production lacks optimized technology. Using a second-generation approach to increase ethanol production is a preferred strategy, since no increase in harvest area will be required and food security is unaffected. 5 For this approach, instead of a readily fermentable sugarcane juice, lignocellulosic biomass is used as raw material. 6 In addition to sugarcane ethanol, maize ethanol production is increasing in Brazil. Since 2014, when the first ethanol plant 100% based on corn was started up, ethanol from maize has grown to 1.34 billion L during the 2018/2019 harvest—4.2% of total ethanol produced in Brazil. 4 This is 4-fold less than the ethanol produced by the entire European Union. 7
Traditionally, Brazilian mills use sugarcane as feedstock to produce ethanol, and this process generates the lignocellulosic biomass sugarcane bagasse. 8 The increase in maize utilization as a feedstock for ethanol production in Brazil makes it necessary to increase research on using corncob, a by-product of maize processing, to produce ethanol. 9 The corncob is comprised of around 40% cellulose, 32% hemicellulose, and 17% lignin. 10 Cellulose is composed of D-glucose, which is readily assimilated by Saccharomyces cerevisiae. However, the hemicellulose fraction is composed mainly of xylose, a C5 sugar that is not assimilated by industrial yeast. 9 Some other wild strains have demonstrated the ability to efficiently ferment xylose to ethanol, making S. stipitis a promising candidate. 11
However, the recalcitrance of lignocellulosic biomass demands a pretreatment step before fermentation to fractionate it into its major components. Usually, these pretreatments are carried out using diluted acids. 12 Depending on the severity of this procedure, the sugars released by cellulose and hemicellulose fraction are accompanied by inhibitory compounds such as furfural, hydroxy-methyl-furfural (HMF), acetic acid, and phenolic compounds. 12 The impact of these inhibitors need to be minimized since fermentation yields can be compromised, especially in C5-fermenting strains. 12 –14 Several detoxifying methods are suggested to remove these compounds, with overliming being one of the most commonly adopted. 15,16
Once detoxified hydrolysate is obtained, the fermentation needs to be carefully carried out. One of the most important parameters to look for in C5 fermentation by yeasts such as S. stipitis is aeration. Ethanol production depends on the microaerophilic environment; without ideal conditions, carbon flux deviates to xylitol and biomass production. 17 Although aeration is considered for fermentation processes, to date it has not been widely explored for inoculum preparation. There are potentially effects on fermentation yields if the inoculum is prepared under an environment that is richer in oxygen than an oxygen-limited medium. To determine this, different fermentations were done using corncob hemicellulosic hydrolysate. The differences in aeration were achieved by analyzing baffled Erlenmeyer flasks (BF) versus unbaffled Erlenmeyer flasks (UF). These baffles cause a more chaotic regimen, which increases the oxygen mass transfer to the medium. 18 Usually these baffles are handmade, and the results are hardly reproducible. 19 To overcome this problem, the flasks had the same design as Corning Pyrex narrow-mouth Erlenmeyer flasks. The fermentations were carried out by preparing the inoculum in these different aeration conditions, and the fermentation parameters were then evaluated.
Materials and Methods
Preparation of the Corncob: Dilute Acid Pretreatment and Vacuum Concentration
Corncob pellets (3.7 kg dry matter, Paraná, PR, Brazil) containing 9% moisture was pretreated with dilute sulfuric acid (50 mg H2SO4/g dry matter) at a solid:liquid ratio of 1:10, as described by Kleingesinds et al. 10 The reaction was conducted in a 50-L stainless steel reactor at 133 and 121°C for 20 min. After this step, the residual solid material was separated by centrifugation (1,000 × g for 60 min). The liquid phase (hemicellulosic hydrolysate), obtained at a temperature of 133°C, was vacuum concentrated in a 30-L stainless steel evaporator at 70°C to increase the xylose concentration to 75 g·L−1 and stored at 4°C for further use as medium fermentation. 10 The liquid phase from dilute acid pretreatment at 121°C was stored at 4°C for further use as an inoculum medium.
Treatment of the Corncob Hemicellulosic Hydrolysates
Both the hemicellulosic hydrolysate obtained at 121°C and the vacuum evaporated hemicellulosic hydrolysate obtained at 133°C were treated by the overliming detoxification method, as described by Rodrigues et al, 20 where calcium hydroxide was added to raise the initial pH 1.0 to 12.0, followed by the addition of phosphoric acid to reduce the pH to 5.5. At each step, the precipitate was removed by centrifugation (2,000 × g for 20 min). Activated charcoal powder (3.0% m/v) was added to the Erlenmeyer flask (1,000 mL) containing 400 mL of each hemicellulosic hydrolysate (pH 5.5). The mixture was maintained at 200 rpm at a temperature of 30°C for 1 h. The solids were removed by centrifugation (2,000 × g for 20 min). Before inoculum and fermentation, both detoxified hemicellulosic hydrolysate obtained at 121°C and the vacuum-evaporated hemicellulosic hydrolysate obtained at 133°C, were autoclaved at 111°C (0.5 atm) for 15 min to be used as inoculum and fermentation medium, respectively.
Microorganism and Inoculum Preparation in Baffled and Unbaffled Erlenmeyer Shaking Flasks
The S. stipitis CBS 6054 was the selected strain since it is one of the most used in literature. 21 It was maintained on agar malt medium and stored at 4°C, then transferred to a fresh tube for incubation at 30°C and used within 24 h. To prepare inoculums, cells were transferred from the tube to grow to 1,000 mL in baffled and unbaffled Erlenmeyer shaking flasks containing 400 mL of medium supplemented with 6.0 g·L−1 ammonium sulfate, 2.4 g·L−1 potassium phosphate dibasic, 1.2 g·L−1 magnesium sulfate heptahydrate, and 0.72 g·L−1 calcium chloride dehydrated.
Baffles are usually made by hand at a local glass workshop and are therefore not reproducible in geometry and size, leading to significant differences in oxygen transfer between the individual flasks. 19 To avoid standard deviation of the baffled flasks and also between unbaffled Erlenmeyer shaking flasks, both had the same design from Corning Pyrex narrow mouth Erlenmeyer flasks. The baffle design (Corning Pyrex 1234D77, Mfr. No.4450-1L) promotes optimal and consistent solution agitation when used with flask shakers.
The exponential phase of growth was determined by the three-phase linear model. 22 This is based on the fact that during the exponential phase of growth, the specific growth rate is constant, and consequently the natural logarithm of cell density is linear as a function of the time. The time range was selected based on the quality of linear adjustment, quantified by the determination coefficient (R 2 ).
The cells were cultivated until the exponential phase of growth and were recovered by centrifugation (2,000 × g for 20 min), washed with sterile distilled water, and used to prepare a cell suspension for inoculum. An aliquot of the inoculum was transferred to fermentation medium to reach an initial cell concentration of 0.8 g·L−1, which was based on a standard curve relating 600 nm absorbance to gDCW·L−1 (Dry Cell Weight).
Fermentation
The fermentation medium was prepared using corncob hemicellulosic hydrolysate (CHH) obtained at 133°C (50 mL) after vacuum concentration and treatment, as described above, in 125-mL Erlenmeyer flasks supplemented with the same inoculum nutrients (6.0 g·L−1 ammonium sulfate, 2.4 g·L−1 potassium phosphate dibasic, 1.2 g·L−1 magnesium sulfate heptahydrate, and 0.72 g·L−1 calcium chloride dehydrated). Cultivation was conducted in an orbital shaker (New Brunswick Scientific Co., Enfield, CT), at 30°C and 150 rpm, with an initial cell density of 0.8 g·L−1. The fermentations carried using inoculum produced in baffled flasks and unbaffled flasks are designated FBF and FUF, respectively. Samples were taken periodically for monosaccharides and ethanol quantification, as well as measure the cell concentration. All the experiments were done in duplicates.
Analytical Methods
Determination of cells concentration, viability, purity, and morphology
Cell concentrations were determined using a Beckman DU 640B spectrophotometer (Beckman Coulter, Pasadena, CA) by reading the optical density at 600 nm wavelength and from the previously determined calibration curve (dry weight × Abs600). The cell viability was evaluated by microscopic analysis, using a solution 0.01 % (w/v) of methylene blue dissolved in sodium citrate 2 % (w/v). The purity and the cellular morphology were observed by fixed slides dyed with fuchsin.
Determination of sugars, ethanol, glycerol and acetic acid
The concentrations of glucose, xylose, arabinose, xylitol, ethanol, glycerol, and acetic acid of hemicellulosic hydrolysates and the fermentation time points were determined by high-performance liquid chromatography, employing Bio-Rad Aminex HPX-87H (300 x 7.8 mm) at 45°C with 5 mM of sulfuric acid as eluent and flow rate of 0.6 mL·min−1. The injection volume was 20 μL. The Waters 410 refractive index detector at 35°C was used to quantify the products. The diluted samples were filtered on Sep Pak C18 filter (Millipore, Burlington, MA) before the injection. Before use, the eluent was vacuum filtered on cellulose ester membrane (0.45 μm pore, 47 mm diameter) (Millipore) and was degassed in an ultrasound bath (Thornton) for 25 min. 20
Determination of specific rates
Specific rates of growth (μx), sugar consumption (μs), and ethanol production (μp) during the experiments were calculated using a geometric method. 23 All time points of the growth curves were obtained by spline fit, using polynomial regression. 24 The natural logarithm was calculated for each point and was fitted using the Origin 8.5 (Microcal Software). The fidelity of the regression was validated by the determination coefficient (R 2 ).
Fermentation Parameters
Ethanol yield (Yp/s, g·g−1) was calculated as the ratio between ethanol concentration and sugar consumption. Ethanol productivity (QP, g·L−1·h−1) was calculated as the ratio between ethanol concentration and time of maximum ethanol production. The efficiency (η, %) of sugar-to-ethanol bioconversion was calculated as the ratio between the experimental Yp/s value (g·g−1) and the maximum theoretical yield value (0.51 g·g−1).
Statistical Analysis
The statistical analysis was performed by the analysis of variance (One-way ANOVA) with a probability level (p) less than 5 % (p ≤ 0.05), using the program STATISTICA 8.0 from StatSoft Inc. (Tulsa, OK).
Results and Discussion
Corncob Hydrolysis
The results of acid hydrolysis are shown in Table 1. At 133°C, release of xylose increased 30.3%, glucose 11.2%, and arabinose 51.2%; however, acetic acid also increased by 71.6%. Because hydrolysis at 133°C showed better sugar release, all downstream processes were performed using this hydrolysis condition. Conductivity was also higher at 133°C, which is in line with the fact that the more severe the treatment, the bigger the ionic release will be, because of the formation of furfural, HMF, and organic acids and the exposure of phenolic and hydroxyl groups of lignin. 25 Regarding sugar release, Cheng et al. used twice the solids charge at similar conditions and obtained 28.7 g·L−1 of xylose, 26 while Rodrigues et al., using another approach for hydrolysis, achieved 21.5 g·L−1 of this pentose. 20 Although the xylose releases obtained in this work were in agreement with previous reports, the glucose and arabinose contents were 2–6 fold lower. 20,26 To proceed to the fermentation step, the corncob hemicellulosic hydrolysate was vacuum concentrated, where the sugars concentration increased about 3-fold, and acetic acid, a volatile compound, only showed a 1.5-fold increase in concentration.
Corncob Hydrolysate Characterization After Hydrolysis at 121°C and 133°C, Using 50 mg Acid Per g of Dry Corncob, After Concentration Step (C) and Treatment Step (T)
Once the robust industrial yeast S. cerevisiae cannot consume xylose, the main product of this hydrolysis, the yeast S. stipitis was usually chosen as an alternative for C5 assimilation. 27 Although the S. stipitis naturally produces xylose reductase and xylitol dehydrogenase, the oxidative path to convert xylose to xylulose, this yeast still suffers from inhibitors present in hydrolysate. 28 The removal of these compounds is a crucial step for fermentation. The overliming step contributed mainly to remove inhibitory phenolic compounds and dehydrated carbohydrates like furfural and HMF, produced due to acid hydrolysis, 16,29 allowing better fermentation conditions. The ionic removal was demonstrated by conductivity decrease, and different from already reported, the overliming step could maintain the sugar concentration. 30
Kinetic Similarities and Disimilarities of Baffled and Unbaffled Flasks in Inoculum Cultivation
The inoculums were prepared in baffled and unbaffled flasks to achieve different oxygen transfer rates. It is known that fermentations carried under microaerophilic conditions result in a higher ethanol yield than in aerobic conditions. 31 However, as far as we can tell from the literature, the effect of aeration during inoculum preparation of S. stipitis before the fermentation process has not been evaluated. Figure 1 shows that, for baffled flasks (BF), the cell growth was shown to be faster, in line with previous works. 31,32 The lag phase remains equal for these conditions, starting growth after 5 h. In non-detoxified hydrolysates, this lag phase can be sustained until 12 h. 33 In both conditions, the stationary phase remains similar, reaching around 10 g·L−1 of cells after 40 h of incubation (Fig. 1A). The pH followed cell growth (Fig. 1B), probably due to acetic acid consumption (Fig. 1C). The acetic acid total depletion occurred after 48 h in baffled flasks, while in the unbaffled flasks this compound remains at 12% of initial concentration after 48 h. These data corroborate with S. stipitis's ability to consume this organic acid at high oxygen availability, decreasing the capability to consume the acetic acid as the oxygen availability decreases. 34 Because of this increase in pH and the aerated conditions, the carbon flux deviated to biomass production instead of ethanol. The optimum pH for ethanol production was around 6.0–6.5, which could explain the absence of ethanol at the inoculum preparation. 35 After 48 h, 23.1 and 19.5% of xylose was consumed, in unbaffled and baffled conditions, respectively (Fig. 1D). Xylose consumption started after glucose depletion (Fig. 1E). Although consumption was demonstrated to be similar, the rate of consumption in baffled conditions was slightly higher until 30 h of cultivation, reaching similar xylose concentration at the end of cultivation. Regarding arabinose, the assimilation of this pentose is strictly aerobic; however, even at higher oxygen availability, the yeast could not consume this sugar (Fig. 1F). 36
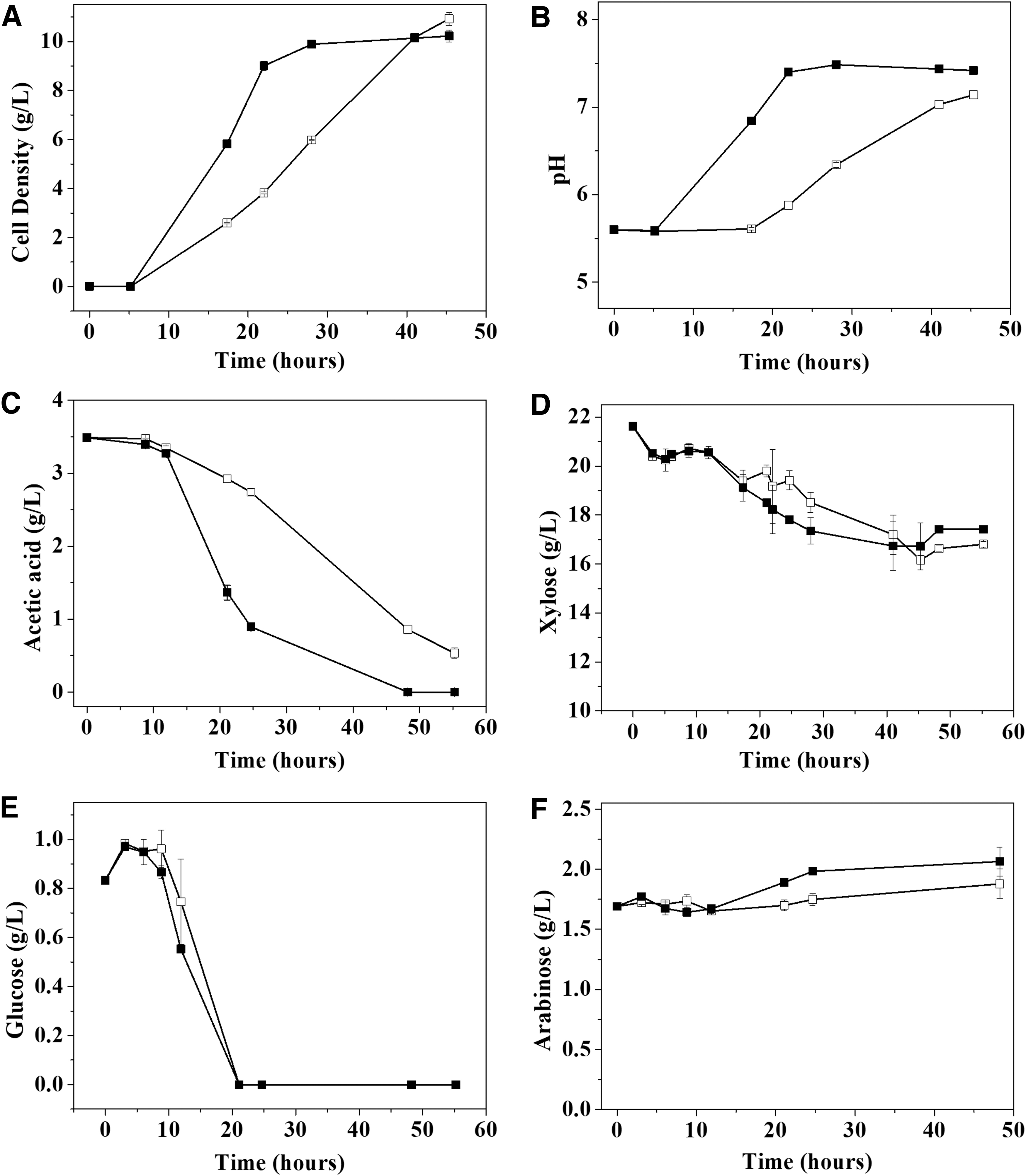
Effect of aeration in Scheffersomyces stipitis CBS 6054 inoculum preparation in hydrolyzed corncob in Baffled (closed squares) and Unbaffled flasks (Open squares). (
Inoculum Growth in Baffled Flasks Presented Higher Ethanol Production after Fermentation
Inoculum preparation could impact fermentation yields. 37 Studies were already done regarding initial biomass concentration 38 and agitation during the fermentation process. 36,39 However, the influence of oxygen transfer rates (OTR) caused by baffled flasks during the S. stipitis CBS 6054 inoculum preparation was never reported. One of the main problems to working with unbaffled shaking flasks is the oxygen supply. 19 Oxygen depletion can slow the entire metabolism of the culture, and the product formation can suffer consequences in productivity and yield. 19 Ideally, the combination of agitation, aeration, and geometry of the flask—with or without baffles—should provide an oxygen transfer rate (OTR) sufficient for cell growth without limitation on the inoculum preparation. Based on the fact that higher OTR led to faster cell growth, as observed by Silva et al. 34 and Su et al., 39 we expected that the exponential phase of growth is also reached faster.
To prepare an inoculum, it is important to achieve high cell density, and the culture needs to be in the exponential phase of growth. 40 Also is important to prepare the inoculum in the presence of xylose, as was done in this work, since xylose induces the expression of its metabolism in C5-fermenting strains. 41 The inoculum preparation in higher OTR was then evaluated. The lag phase for both conditions, BF and UF, remained the same, until 5–7 h. At the initial pH 5.5, acetic acid is mostly dissociated, allowing easy diffusion into the cell cytoplasm, decreasing the intracellular pH and leading to a long lag phase. 35 The exponential phase was reached faster in baffled flasks. What was 12–18 h for the exponential phase in unbaffled flasks, decreased to 7–14 h of agitation (Fig. 2). Besides the lower time required to achieve the exponential phase, the cell density obtained in baffled flasks during this period (7 h; 0.20 g·L−1 to 14 hours; 2.78 g·L−1) was higher than obtained that in unbaffled flasks (12 h; 0.01 g·L−1 to 18 hours; 0.70 g·L−1).
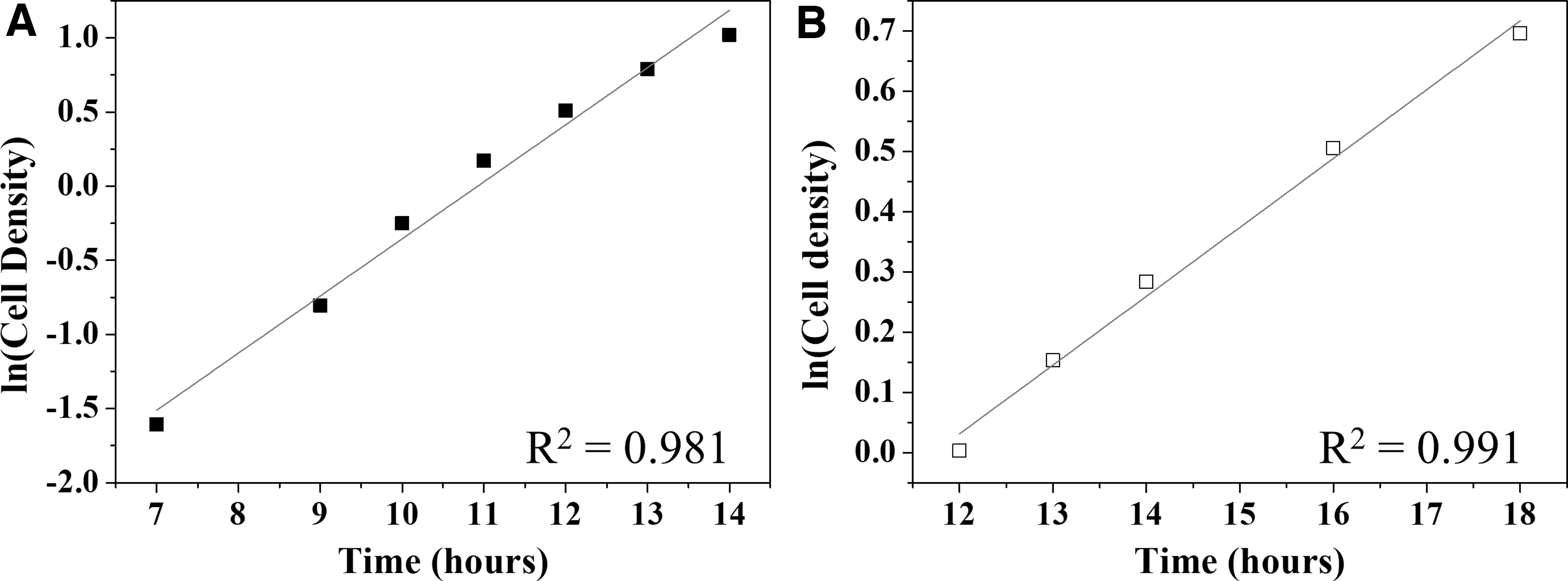
Analysis of the exponential phase of growth on inoculum preparation of Scheffersomyces stipitis CBS 6054 using baffled flasks (closed squares) and unbaffled flasks (open squares).
Despite the less time required to reach the exponential phase and higher density cells, the different inoculums were evaluated against the fermentation capacity. The fermentations were inoculated in the exponential phase in both cases, and starting with 0.8 g·L−1 of initial cell density.
In both fermentations, the pattern of growth was extremely similar during the 96 h tested, reaching 30.20 ± 0.03 and 30.35 ± 0.24 g·L−1, for FUF and FBF, respectively (Fig. 3A). S. stipitis is known to have a great respiratory capacity, even higher than industrial yeast S. cerevisiae. 42 This capacity can lead to achieving high cell growth. An increase in pH during the fermentation also contributed to shifting the carbon flux to biomass production. 42 The glucose was consumed first, as expected, and after 12 h was depleted from the medium in FBF. After this same amount of time, FUF presented at 0.3 g·L−1, and no residual glucose was observed at 24 h (Fig. 3B). Xylose was only consumed after glucose depletion, even at low glucose concentration (< 3.0 g·L−1), which is expected from the majority of xylose-fermenting organisms that suffer from catabolite repression, also named diauxic shift (Fig. 3C). 43 Besides this repression, the mixed sugars glucose and xylose compete for the transporters binding sites, 44 explaining the lag in xylose consumption. Xylose concentration in this assay seems not to cause osmotic stress in the yeasts since the cell viability analyzed on methylene blue showed above 90% of viability during the fermentation. In addition, the medium presented only traces of glycerol, a possible indicator of cell membrane damage for osmotic stress. 45

Fermentation of Scheffersomyces stipitis CBS 6054 using inoculum prepared in unbaffled flasks (FUF - open squares) or baffled flasks (FBF - closed squares). (
Starting the fermentation at 3.5 g·L−1 of acetic acid, 72% was consumed under both conditions. Although this organic acid presents severe inhibitory effects on S. stipitis CBS 6054 fermentation, at this concentration the yeast could consume a great part of the inhibitor. 46 Acetic acid consumption led to an increase in pH of the fermentation broth (5.5 to 7.0). The lower OTR condition (FUF) presented higher xylose consumption, reaching 16% of residual xylose against 31% in FBF after 96 h. This higher consumption was also observed in inoculum preparation using BF. This observation does not corroborate with previous works when at higher oxygen availability, xylose consumption was higher. 39 This higher xylose consumption observed in FUF (54.12 versus 44.01 g·L−1) did not bring advantages in glycerol, xylitol, or ethanol production; quite the opposite, it showed lower ethanol production (10.34 versus 9.20 g·L−1) (Fig. 3D). Under both conditions, less than 0.3 g·L−1 of glycerol and no xylitol were detected. Arabinose consumption was less than 1.0 g·L−1.
In FBF, ethanol production started earlier, at 12 h, wherein with FUF, traces of ethanol were only detected after 48 h. The higher ethanol productivity (0.108 versus 0.096 g·L−1·h−1) and final ethanol concentration (10.3 versus 9.2 g·L−1) suggests higher efficiency using the inoculum produced in baffled flasks. Table 2 summarizes the fermentative parameters obtained from both conditions.
Fermentative Parameters Obtained Using Inoculum Produced in Unbaffled Flasks (FUF) and Baffled Flasks (FBF)
Sugar is the sum of xylose and glucose, and the yields were calculated using this value; 2 Conversion efficiency was calculated based on ethanol and biomass production.
With all fermentation parameters, the assay using inoculum prepared with more aerated conditions were higher than inoculum prepared on unbaffled flasks. The maximum specific rates of ethanol production (μPmax) and cellular growth (μXmax) were 8-fold and 2-fold higher, respectively, when baffled flasks were used for inoculum cultivation. The maximum specific rate of xylose consumption (μSmax) was slower than FUF, even though the ethanol productivity in FBF was still higher. Although the YP/X still higher, the initial idea that aeration variation during the inoculum preparation impact on the fermentation was proved to exist. The YP/S increased and total sugar conversion was higher, demonstrating the beneficial effect of inoculum aeration on the fermentation. Comparing the maximum specific rate of xylose consumption with those obtained by Kleingesinds et al. 10 in corncob cellulose hydrolysis and fermentation, using the same yeast strain (S. stipitis CBS 6054), the values were similar (0.13 vs 0.12 h–1), although less inhibitory compounds were present in the cellulosic hydrolysate. 10 Looking at ethanol productivity, similar results are found in rice straw hydrolysate fermentation. 47 It can be noted that non-optimized conditions were used in the fermentation carried out in this study. The results obtained here can be enhanced by several methodologies, such as biotreatment of hydrolysate, as done by Fonseca et al. 47 The concept evaluated here was limited to the baffled and unbaffled flasks in inoculum preparation, and further studies can be directed to the optimization of fermentation conditions.
To validate this difference obtained with different inoculum preparations, an analysis of variance was performed. Ethanol production response was evaluated by the independent variables, FBF and FUF. The statistical significance is illustrated in Table 3.
Analysis of Variance for Ethanol Production as a Function of Fermentation Using Baffled Flasks and Unbaffled Flasks
SS, sum of squares; DF, degree of freedom; MS, mean square.
The ANOVA showed that FBF was more efficient in ethanol production than FUF at 95% confidence level (p ≤ 0.05). This way, ethanol production during fermentation was improved by the use of inoculum prepared in baffled flasks compared to unbaffled flasks.
Conclusion
The conditions of inoculum are extremely important in fermentation yields. This work evaluated the influence of fermentation yields using inoculum prepared at different oxygen transfer rates, using baffled and unbaffled flasks conditions and corncob hemicellulosic hydrolysate. The results demonstrated that lower time was required to achieve the exponential phase of growth using a more aerated regimen (7–14 h in baffled flasks versus 12–18 h h in unbaffled flasks), and also baffled flasks showed higher biomass production in this phase (0.20–2.78 versus 0.01–0.70 g·L−1 of cells). Despite these advantages, the fermentation FBF presented an increase in yield (YP/S of 0.17–0.23) and ethanol production (9.20–10.34 g·L−1), demonstrating the importance of oxygen providing on the S. stipitis CBS 6054 inoculum preparation in fermentative assays.
Footnotes
Acknowledgments
The authors thank Coordenação de Aperfeiçoamento de Pessoal de NÚvel Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grant #2018/02865-2 for financial support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank CAPES, CNPq, and FAPESP for financial support.