
Abstract
Myxobacteria have been mainly mining for their drug-like compounds while their essential and versatile protease enzymes is less explored. In this study, in order to obtain myxobacterial strains with the ability to produce extracellular protease, soil samples were collected from different geographical locations of Iran and were cultured on isolation media. The isolates were identified following purification by morphological and molecular methods. All of the identified isolates belonged to Myxococcus, Archangium, Corallococcus and Cystobacter genera. Then, these strains were subjected to the evaluation of protease activity. Archangium sp. UTMC 4504 was identified as a strain with high protease activity (60 Unit). The enzyme was precipitated using up to 60% of the saturation by adding ammonium sulfate in the fermentation supernatant. The optimum temperature of the enzyme was 50–60°C and its optimal pH was 8. The enzyme was inhibited by PMSF, which could indicate its categorization under the serine proteases. Because of the capability of this enzyme in maintaining its activity at high temperatures and alkaline pH, it can be considered a suitable candidate for mass production purposes, including industrial detergents, food industry, or medical applications.
Introduction
Proteases are the most important group of industrial enzymes, accounting for nearly 60% of the total global enzyme sales. 1 –3 These enzymes can be used widely in various industrial purposes, such as detergents, feather processes, food processing, silk gumming, pharmaceuticals, bioremediation, biosynthesis and biotransformation. 1 Proteases are found in almost all organisms but bacterial proteases are the most diverse compared to animal, herbal and fungal proteases. 4 The rate of multiplication, limited cultivation space requirement, easier downstream processing and genetic manipulation are among the characteristics that make bacterial sources of extracellular proteases 5 preferable to plants and animals. 6 New microorganisms that produce novel enzymes suitable for commercial purposes are frequently discovered. 7 Most of the commercialized neutral and alkaline proteases are produced by Bacillus spp. 8 These enzymes are mostly unstable under extreme conditions like high temperature and alkaline pH more commonly found in the industrial process; moreover, with increasing demand from industries and the global market, researchers are looking for valuable natural resources for novel products, including useful enzymes. 9 Myxobacteria are valuable sources for developing novel proteases and for producing alkaline proteases in industrial sectors. 10,11 These bacteria are distinctive owing to their complex life cycle. Upon starvation, bacterial cells aggregate, forming fruiting bodies. 12 Myxobacteria are Gram-negative, mainly aerobic, display gliding motility, and produce myxospores. 13 They thrive in diverse natural environments and are found mainly in soil habitats. Myxobacteria also has a great potential for the production and secretion of extracellular molecules that have significant potential for industrial applications, such as carotenoids, antibiotics, and a variety of hydrolytic enzymes. 14 –16 Myxobacteria are micropredators that secrete enzymes and antibiotics to kill microorganisms and lyse cells. 17 Myxobacteria secrete many bacteriolytic enzymes, including proteases, during their vegetative growth, letting them feed on other bacteria to survive during starvation. 16,18 While most studies have focused on antibiotic production by Myxobacteria, there are only a few reports about their enzymatic potential. This study aims to isolate and screen Myxobacteria and analyze their capability to produce protease enzyme.
Materials and Methods
Chemicals and Reagents
The principle chemicals used in the study include various inorganic salts. Trichloracetic acid, Tris, Coomassie Brilliant Blue R-250 and Coomassie Brilliant Blue G-250 were purchased from Merck Co. (Darmstadt, Germany). Azo-casein was purchased from Sigma-Aldrich (St. Louis, MO, USA). Vitamin B12 was purchased from Merck which was obtained from animal source. Skim milk was purchased from Fluka (Munich, Germany).
Experimental Methods
Sample collection, isolation
Fifty samples were collected from different regions of Iran. Samples contained soils, wood pieces, small dead animals. 19 The samples were air-dried immediately after collection and stored at 4°C. 20 The isolation method, based on predatory activity, was applied with slight modifications. A cross streak of dead cells of Escherichia coli ATCC 8739 (UTMC 1407) was made on WCX agar plates: (g/L) CaCl2 ·2H2O 1, MgSO4. 2H2O 1, KNO3 1, (NH4)2SO4 0.2, K2HPO4 0.2, MnSO4 · 7H2O 0.1, agar 15 (pH 7.5); and chitin agar plates: (g/L) chitin powder 5, CaCl2 ·2H2O 1, MgSO4. 2H2O 1, KNO3 1, (NH4)2SO4 0.2, K2HPO4 0.2, MnSO4·7H2O 0.1, agar 15 (pH 7.5). After autoclaving, 50 μg mL−1 cycloheximide was added to the media. The cultivated plates samples were incubated under humid and dark conditions at 28°C for 2–3 weeks. 21
Purification and maintenance of Myxobacteria
The isolates were recognized by the detection of fruiting bodies or swarm on agar under the stereomicroscope, and vegetative cells and myxospores were colored by Gram stain and observed under an optical microscope (Standard 25; Carl Zeiss AG, Oberkochen, Germany). 22 Isolates were identified according to the criteria previously indicated by Reichenbach and Dworkin. 23
The fruiting bodies or swarm of Myxobacteria were removed from the isolation media and cultured on VY/2 medium (g/L: Baker's yeast 5, CaCl2 .2H2O 1, agar 15, Vitamin B12 0.0005) and WAT medium to purify the strains. The cultures were incubated at 28°C for 3–10 days. After being purified, the purity of Myxobacteria was checked through inoculation in CY liquid medium ((g/L) Casitone 3, yeast extract 1, CaCl2 .2H2O 1) and observed with an optical microscope. Myxobacteria were inoculated into the CY liquid medium and cultured at 28°C, accompanied by shaking for 48 h. 24 The pure cultures of isolates were maintained as 20% (v/v) glycerol suspensions at −70°C and freeze-dried ampoules.
Molecular identification of selected strains
The strains were cultured in CY liquid medium and incubated at 28°C for 4 days at 220 rpm. Genomic DNA was isolated using DNA extraction kit (Gene All, Seoul, Korea) and 16S rRNA genes were amplified using universal primers (9F, 1541R).
The primer sequences were designed for sense primer, 5'-GAGTTTGATCCTGGCTGAG-3' and antisense primer, 5'-AGAAAGGAGGTGATCCAGCC-3'. 10 PCR reaction was as followed: 1 cycle of denaturation at 94°C for 5 min, 28 cycles of 94°C for 45 s, 58°C for 55 s, 72°C for 75 s, and additional extension at 72°C for 10 min. The homology search of the 16S rRNA sequences was conducted using BLAST program at NCBI. Multiple alignments of sequenced nucleotides were carried out using Clustal W. 25
Screening of protease activity
The protease activity of the strains was evaluated by the skim milk hydrolysis method. 26 For this purpose, isolates, the type strain of Myxobacteria and control strains (Bacillus licheniformis DSM 8785, Bacillus cereus DSMZ 31, Nocardiopsis arvandica UTMC1492) were cultured on skim milk agar (g/L): skim milk powder 10, CaCl2 ·2H2O 1, MgSO4. 2H2O 1, KNO31, (NH4)2SO4 0.2, K2HPO4 0.2, MnSO4 · 7H2O 0.1, agar 15 (pH 7.5–9.0) and incubated at 30°C for 48 h. After incubation, strains exhibiting the largest cleared zone around their colonies were purified and assayed for protease activity.
Production of crude enzyme
To produce crude enzyme, first a fermentation flask of the top strain was cultured in CY-H medium: (g/L) Casitone 1.5, yeast extract 1.5, soymeal 1, glucose1, starch 4, CaCl2.2H2O 1, MgSO4.7H2O 0.5, Fe (III)-EDTA 0.0008 ([pH 7.5]) and shaken (160 rpm) for 8 days at 28°C. 27 Following incubation, the fermented medium was centrifuged at 10,000 rpm for 10 min at 4°C. The cell-free supernatant was applied as a crude enzyme to screen for protease activity.
Measurement of protease activities
Azocaseinase activity was evaluated as follows: 100 μL of azocasein solution (1% w/v in 50 mM Tris–HCl buffer, pH 8.0) was incubated at 50°C for 10 min. After that, 100 μL of enzyme solution was added to azocasein solutions. The mixture was further incubated for 30 min at 50°C. Then, 100 μL of trichloracetic acid (5% v/v) was added to precipitate the proteins. The mixture was centrifuged at 12,000 rpm for 5 min, and the absorbance of the supernatant was determined at 450 nm. 28 One unit of the enzyme activity (U) was set as an increase of 0.01 in absorbance for 1 mL of enzyme solution.
Partial purification of the protease
Cell-free supernatant of the top strain was gathered by centrifugation at 10,000 rpm for 10 min. Ammonium sulfate was slowly added to the supernatant to result in fractions with saturations of 0–40%, 40–60%, and 60–80%. The mixtures were incubated overnight at 4°C. 29 The precipitate was collected by centrifugation at 12,000 rpm for 10 min, dissolved in 50 mM Tris–HCl (pH 8.0), and dialyzed against the same buffer to remove residual ammonium sulfate.
SDS-PAGE and zymography
SDS–PAGE was performed on a 4% stacking and a 12% running gel based on the method of Laemmli (1970), and protein bands were visualized by staining with Coomassie Brilliant Blue G250 (Sigma). 30 Zymography was performed on SDS–PAGE as described by Hadder et al. 1 After electrophoresis, the gel was incubated at 40°C for 1 h in 50 mM Tris–HCl (pH 8.0) containing 1% casein. A clear zone against a blue background on the gel revealed the protease activity.
The method described by Bradford (1976) was applied using bovine serum albumin (BSA) as the standard 31 to determine the protein concentration of the samples. For this purpose, 5 μL of the protein solution was added to 250 μL of the Bradford dye and kept for 10 min at 25°C. The mixture was loaded into microplate wells, and then absorbance was measured at the 595 nm wavelength.
Physicochemical Characteristics of the Partially Purified Enzyme
Effect of temperature on partially purified protease
The optimum temperature for this enzyme against azocasein was established by incubating the reaction mixtures in 50 mM Tris–HCl (pH 8.0) at different temperatures (20–90°C) for 5 min. 32 The relative enzyme activity was determined as a percentage of the maximal activity. Thermal stability assay was performing by incubating 10 μL of enzyme solution in 50 mM Tris–HCl (pH 8.0) at various temperatures (40–60°C) for 120 min. The residual enzyme activity was calculated as a percentage of the starting activity.
Impact of pH on partially purified protease
The impact of pH 5–12 on hydrolytic activity against azocasein was measured at 60°C using 50 mM each of acetic acid-sodium acetate (pH 5.0–5.5), sodium phosphate (pH 6.0–6.5), Tris–HCl (pH 7.0–9.0), glycine–NaOH (pH 9.5– 10.0), and Na2CO3–NaOH (pH 11.0–12.0) as buffers. 33 To evaluate pH stability, enzyme solutions were pre-incubated in buffers of different pH values at 30°C for 6 h before determining the residual activities. The remaining activities were determined as percentages of the activity of the enzyme at pH 8.0 and 4°C in 50 mM Tris–HCl buffer.
Effect of protease inhibitors on the activity of partially purified protease
The effects of phenylmethylsulfonyl fluoride (PMSF) as a serine protease inhibitor and ethylenediaminetetraacetic acid (EDTA) as a metalloprotease inhibitor were evaluated by the enzyme pre-incubation in 10 mM solutions of each inhibitor at 25°C for 30 min. The residual activity was then tested using azocasein as substrate. 34
Results and Discussion
Isolation and Purification of Myxobacterial Strains
A total of five myxobacterial strains were isolated from soil and bark samples collected from different regions of Iran, but only four survived the purification process. According to the morphological traits (shapes of vegetative cells, myxospores, and fruiting bodies), the vegetative cells were slender rods, with tapering or rounded ends (Fig. 1a). All isolates showed gliding motility, and swarms appeared often as shallow sunken craters (Fig. 1b). The swarm area enlarged and reached the rim of the plastic dish in 2–3 weeks. Fruiting body presented with and without sporangiole, which were white and yellow to red color in color (Fig. 1c).

Macromorphology of myxobacterial strains isolated in this study (name of genus taxon + Genebank accession number). (
Molecular Identification of the Potent Selected Strains
Based on the molecular identification and sequencing of 16S rRNA, these isolates were identified as Myxococcus sp. (100% sequence similarity to the type strain Myxococcus virescens), Corallococcus sp. (99.8% sequence similarity to the type strain Corallococcus exiguous), Cystobacter sp. (100% to Cystobacter badius) and Archangium sp. (99.7% sequence homology to the type strain of Archangium violaceum), respectively.
Screening of Protease Activity of Isolates
The strain Archangium sp. UTMC 4504 was selected for evaluation of its protease enzyme and deposited in NCBI database under the accession number of MF455220. The isolate was able to hydrolyze the skim milk agar with a dimension 26 mm (Fig. 2). The rate of hydrolysis in Archanium UTMC 4504 was higher than that of other isolates, and this rate was close to the strains of Bacillus that are potent in the production of proteases.
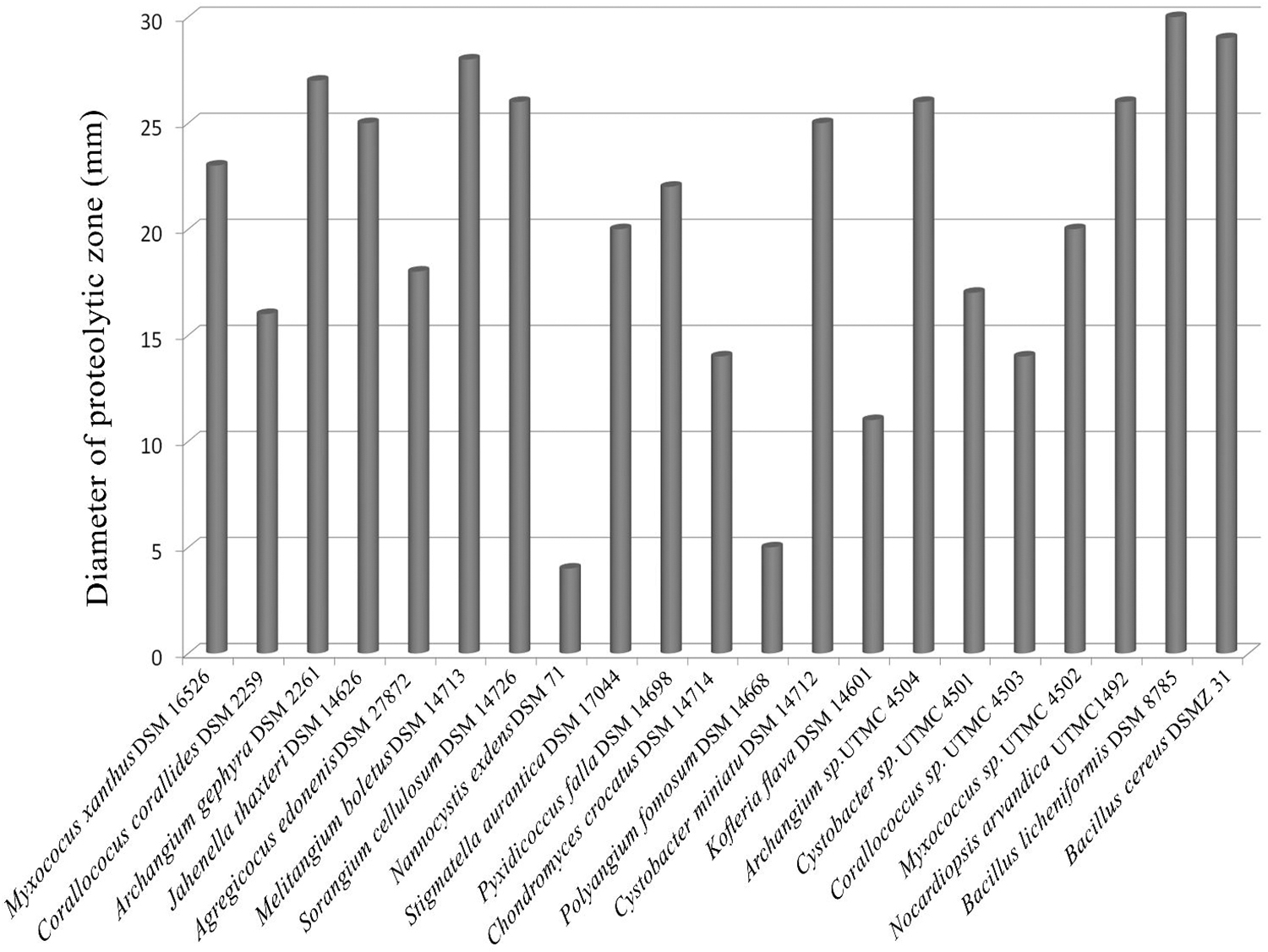
Comparative protease activity of myxobacterial strains of this study in contrast to other potent protease producers of myxobacterial type strains and representatives of Bacillus and Nocardiopsis on Skim milk agar.
Assay of Enzyme Activity
In qualitative assessment, azocaseinolytic activity by Archangium UTMC 4504, Nocardiopsis arvandica UTMC 1492 and Bacillus licheniformis UTMC 2390 were 0.8 U/mL, 0.6 U/mL and 1.2 U/mL, respectively (Fig. 3 ).

Quantitative assay of protease activity of the protease positive strains in comparison to other potent protease producers of Bacillus, Nocardiopsis and myxobacterial members.
Kinetics of Growth and Protease Activity
The production of proteases by strain Archangium sp. UTMC 4504 was investigated during 8 days of cultivation in the production medium. After a day lag phase, exponential growth was observed on the third day, and the stationary phase was reached on the fifth day. Interestingly, strain Archangium sp. UTMC 4504 secreted extracellular proteases at the beginning of the exponential phase (third day) and achieved maximal production of up to 2,445 U/mL at the end of this phase (fifth day) (Fig. 4).
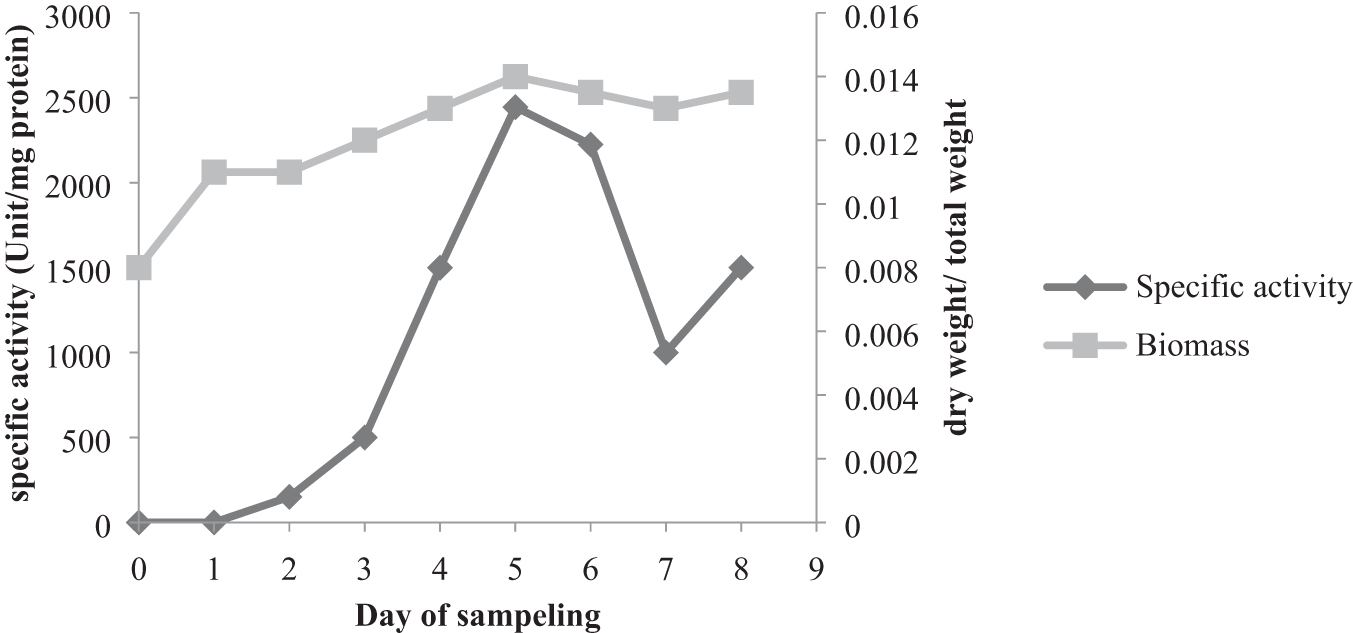
Growth and extracellular proteases production of Archangium sp. UTMC 4504 during 8 days of growth in CY-H medium and incubation at 28°C.
Partial Purification of Extracellular Proteases Produced by Archangium SP. UTMC 4504
The fraction of 40–60% saturation from Archangium sp. UTMC 4504, with a superior activity of 12,123/L U/mg protein, had the highest protease activity (Fig. 5); however, the fraction 0–40% also had a high rate of activity, unlike the 60–80%
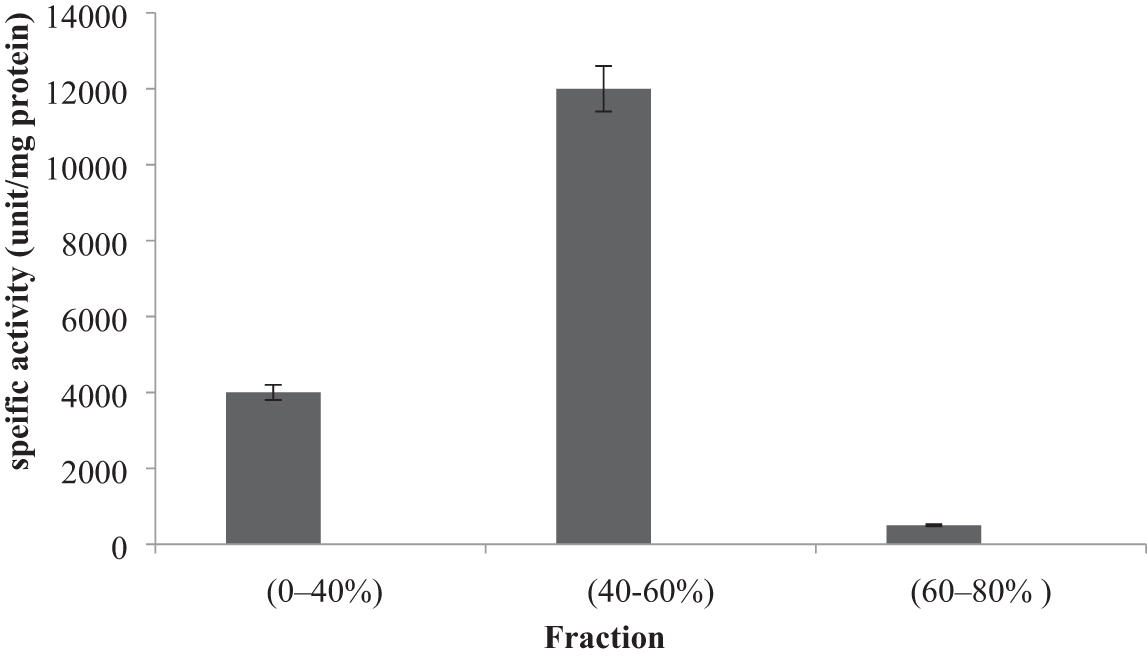
Specific enzyme activity of extracellular proteases produced by Archangium sp. UTMC 4504 for each fraction after precipitation with ammonium sulfate. Fraction of 40–60% with activity of 12,123/L U/mg protein had the highest protease activity compared to other fractions (results are reported as mean ± standard deviation).
Molecular Weight Estimation
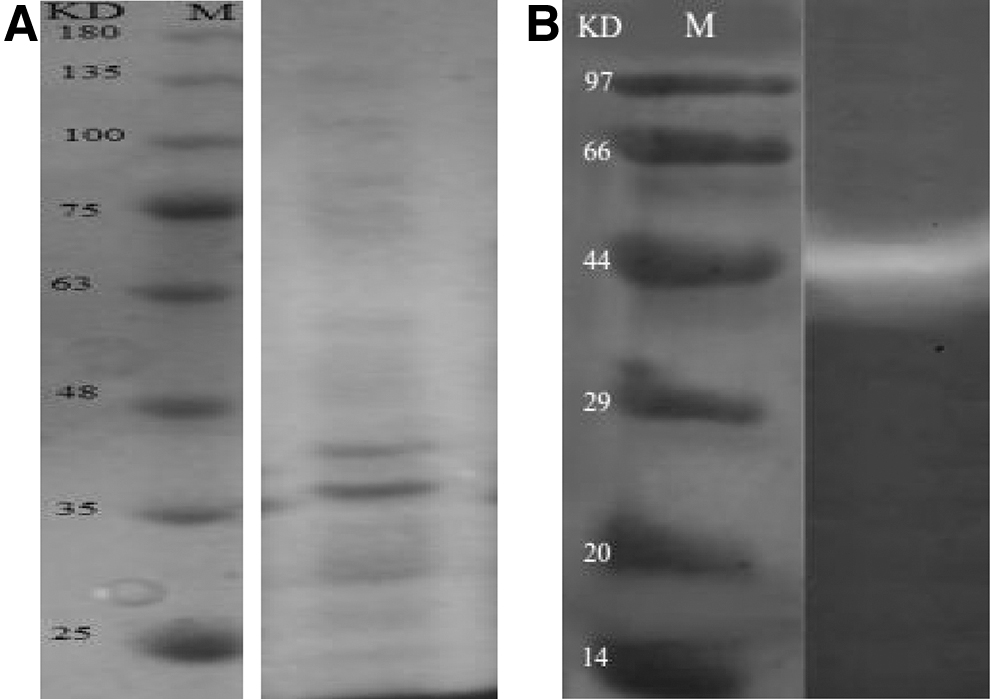
The partially purified enzyme was further analyzed by SDS–PAGE, which showed different bands; two bands corresponded to 39 and 45 kDa, suggesting that the purified enzyme was not a homogeneous material (Fig. 6a). The partially purified enzyme from culture supernatant appeared as a single band corresponding to a molecular weight of around 45 kDa which was designated protease (Fig. 6b).
Effects of Temperature and PH on Enzyme Activity
The enzyme was active at temperatures between 20 and 90°C. It showed maximum enzyme activity at 50°C and 60°C. It retained 27% of its maximum activity at 20°C, and retained 25% of its maximum activity at 90°C (Fig. 7a).
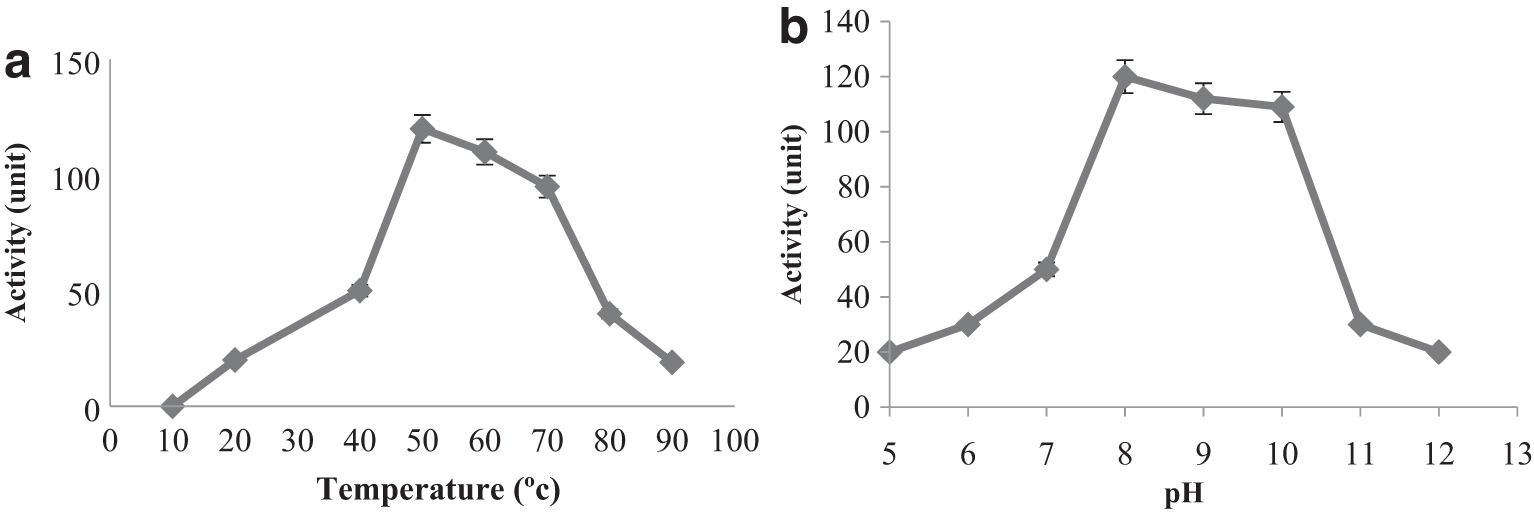
Effect of temperature on protease activity of Archangium sp. UTMC 4504. The enzymatic activity was determined in 50 mM Tris–HCl buffer (pH 8.0) (
The enzyme showed no activity at pH between 3.0–4.5. The enzyme was active at pH between 5.0–12.0. The enzyme showed the maximum activity at pH 8.0, and was highly active (about 87% of the maximum activity) between pH 8 and 10.0 (Fig. 7b).
Effect of Enzyme Inhibitors on Protease Activity
The effect of enzyme inhibitors on protease activity was investigated (Table 1). Enzyme activity was not or only slightly inhibited by EDTA at a concentration of 10 mM. The inhibitory effect of EDTA was concentration-dependent but only reached 33% at a concentration of 10 mM. These results suggest that partially purified extract may not be a metalloprotease. The strong inhibition by PMSF suggests that partially purified extract could be a serine protease (Table 1).
Effects of Enzyme Inhibitors on Activity of Extracellular Protease Produced by Archangium sp. UTMC 4504
The control contained Tris–HCl buffer and enzyme solution.
Discussion
Myxobacteria are believed to be a great source of novel compounds, including enzymes and other industrially valuable bioactive compounds. 35 Approximately 40% of secondary metabolites produced by Myxobacteria represent novel structures. 36 Many of these metabolites possess unusual or even novel structures and mechanisms of action. 37 Among major myxobacterial metabolites are hydrolytic extracellular enzymes that indicate their ecologic function for the predatory behavior of these bacteria. 21,38 They are predators via production of the extracellular enzymes that catalyze the degradation of biological macromolecules. 39 The potential of Myxobacteria to synthesize a particular metabolite is strain-specific, making an attractive proposition for bioprospection of potent enzymes as well. 17
With recent advancements in biotechnology, it has become possible to modify microbial enzymes for distinctive characteristics to catalyze under various extreme conditions, such as high pH, temperature, oxidative, organic solvents, etc. 40 Since microbial proteases have several applications in various industries, efforts to indentify, isolate and purify these enzymes are accelerating. In this study, we investigate Myxobacteria as a better source of protease production. As studies on Myxobacteria from Iran habitats are scarce, in this study myxobacterial strains with the ability to produce extracellular protease were isolated from soil samples of Iran.
Protease-producing strains are often recognized by growth on skim milk or casitone-containing media. 41 Strains showing maximum zones of clearance around their colonies on the PDCY medium 42 were screened in this study.
The protease is synthesized and secreted initially during the exponential growth phase, with a substantial increase close to the end of the growth phase and maximum protease production in the stationary growth phase. 43 The obtained results in the present study showed that the production of proteases by Archangium sp. UTMC 4504 is initiated after the first day of the lag phase and reaching a maximum at the beginning of the stationary phase on the fifth day of fermentation. Furthermore, the production of this enzyme was growth-dependent. Since the bacteria are metabolically more active in the exponential phase, more enzyme (protease) production can be observed in this phase. This trend is similar to the previous report by Poza et al. revealing that the production of rennet for cheese production by M. xanthus strain 422 increased during the exponential phase of growth, reaching a maximum at the beginning of the stationary phase. 16 The production of alkaline proteases by Bacillus sp. B001 started at the exponential phase (6 h) and achieved maximal production of up to 34,277 U/mL at the end of this phase (30 h). 44
The molecular masses of alkaline proteases range from 30 to 50 kDa. 45 The molecular weight of an extracellular protease from Myxococcus virescens was estimated to be 26 kDa. 11 The molecular weight of the milk-clotting protease from Myxococcus xanthus was estimated to be 40 kDa. 16 An extracellular metalloprotease with the molecular weight of 31 kDa is also reported from Myxococcus xanthus. 46 In this study, the molecular weight of Archangium sp. UTMC 4504 protease was 45 kDa, which makes it suitable for both industrial and purification processes.
The optimal conditions for protease activity from Archangium sp. UTMC 4504 were pH 8.0 and 50–60°C. The optimal pH for the extracellular protease of Myxobacter strain AL-1 was 8.0 and the optimal temperature was 60°C. 47 The protease reported from Myxococcus xanthus strain 422 has an optimal pH of 6.0 and an optimum temperature of 37°C. 16 Gnosspelius et al. reported a protease with an optimum pH of 8.5 and a temperature of 45°C. 11 The optimal temperature for the protease of isolates of Myxococcus sp. KYC 1134 and KYC 1139 was 60°C with optimal pH 7.0. 42 In addition, according to the study of Olajuyigbe et al., three Bacillus species (B. macerans IKBM-11, B. licheniformis IKBL-17 and B. subtilis IKBS-10) had maximum protease production at 60°C and pH 8.0. 48 Protease production was about 65% even at 65°C and pH 10.0. 48 The results of the present study show that this enzyme is also properly active in a wide temperature range of 20–90°C and pH 5.0–12.0, and it could maintain 30% of its activity at 90°C and pH 12, making it suitable for industrial applications.
The enzyme underwent incubation with several inhibitors, and the protease activity was analyzed to determine the type of protease. PMSF, a common serine protease inhibitor, inactivated the purified protease completely at a concentration of 5 mM. The purified serine alkaline protease from Bacillus stearothermophilus AEAL2 and Bacillus sp. B001 has also been entirely inhibited by 10 mM and 15 mM of phenylmethylsulfonyl fluoride (PMSF), respectively. 44 Thus, it can be concluded that this enzyme is an alkaline serine protease. It is stable at alkaline pH and high temperature. Thus, strain UTMC 4504 might be an economic source with potential applications in the enzyme industry, and proteolytic enzymes produced by Myxobacteria represent hold potential for application in various industries. The proteolytic enzymes produced by Myxobacteria can have clinical and agricultural applications, in addition to industrial applications. 42 For instance, Myxococcus virescens and Corallococcus exiguous produced extracellular proteinases that improved plant health by protecting against plant pathogens. 13
Keane et al. (2016) 49 showed that Myxococcus xanthus, during its predatory life cycle, secretes several secondary metabolites along with several classes of hydrolytic enzymes, including MepA protease, chitinase, endonuclease and trypsin used to digest prey macromolecules. There is an increase in the production of hydrolytic enzymes during sporulation of M. xanthus. 50 From the mixture of the enzymes produced by M. xanthus, prolyl oligopeptidase (POP enzyme) was found as an important enzyme in the treatment of diseases such as Alzheimer's, amnesia, depression, cancer and celiac disease 51 and the physicochemical characteristics of the enzyme produced by recombinant E. coli has been evaluated. 33 Angiotensin-converting enzyme (ACE) from Stigmatella aurantiaca cleaves Aβ peptides as a metalloproteinase similar to human ACE. Therefore it has the potential to be used in designing new therapeutic strategies against Alzheimer's disease. 52
The main enzyme and secondary metabolite reported from Archangium spp. include Tubulysins, a potent cell multiplication inhibitor compared with taxol, epothilone and vincristine, influencing multidrug-resistant cell lines. Argyrin A produced by Archangium gephyra is another highly cytotoxic compound against various human tumor cell lines.
53
The extracted enzymes from Archangium spp. include serine-type keratinolytic proteases;
Conclusion
The significance of the protease enzyme purification from Myxobacteria is that, under nutrient deficient or stressful conditions, these bacteria produce a large number of proteolytic enzymes to digest their vegetative cells and transform into resistant spores. 49 Therefore, mass production of enzymes does not require expensive nutrients to grow and ferment these bacteria, which is critical given the need for industries to optimize input costs. 7 Therefore, the purification of the protease enzyme from Myxobacteria can be among future alternative protease resources. In addition, due to the resistance of their protease to alkaline conditions and activity at high temperatures (60°C), 57 these bacteria have the potential to be applied across the food, fermentation, detergents, leather and tannery, oil, alcohol and pharmaceutical industries. For instance, the protease-containing cocktail of Pancreatin® is an alternative for digestive enzymes commercialized for chronic pancreatitis. Furthermore, protease has the potential to be used in the proteolytic destruction of protein aggregates formed in diseases such as cancer and Alzheimer's.
Some of the proteases used as commercial enzymes are structurally and functionally active in a particular temperature range and are used exclusively in a particular industry. For instance, the commercial protease trypsin is active in specific temperature and alkaline conditions in the leather industry 58 and cannot function in the food industry at low temperatures.
The protease extracted in this study is active in a wide range of temperatures. This protease has not only a maximum activity at 50–60°C, but also retains 25% of its activity at 20°C. With regard to this wide range of catalytic activity, it can be used as a multi-functional protease enzyme in various industries, especially in the production of hydrolyzates and in the detergent industry for use at low temperatures, where it can be cost-effective and safe to use. For example, Novo Nordisk Bio-industry Company in Japan has produced a protease called Kannase from Bacillus sp., which can be active at low temperatures. Consequently, future studies are recommended to explore other structural and functional properties of these enzymes. Moreover, they can be genetically manipulated to generate new enzymes with altered properties that are compatible with multiple usages with numerous industrial applications in detergents, leather, and chemical industries.
Footnotes
Acknowledgments
The authors would like to thank Hoda Sabati and Maryam Noordadi for their friendly assistance.
Author Disclosure Statement
Authors declare no conflict of interest.
Funding Information
The authors wish to thank the vice-chancellor of research of the University of Tehran for the partial financial support of the project.