
Abstract
Polyvinyl alcohol (PVA) is a synthetic polymer widely used for industrial applications and having negative impacts on the environment. In this work, we present a novel, potent PVA degrader, MBB8, isolated from the Gulf of Mannar. MBB8 was identified as Enterobacter cloacae based molecular analysis using 16S rRNA gene sequence as well as biochemical and evolutionary distance analysis. This is the first report of a PVA degrader from the Enterobacter genus. The results showed 83% of the PVA present in the culture medium was degraded by E. cloacae MBB8 after 48 h of incubation with 30°C at 145 rpm agitation. The results obtained from FTIR showed notable differences in PVA-degradation stretches (3,000 to 3,300 cm−1 and 2,500 to 3,000 cm−1) at 24 and 48 h, respectively, compared with control (before degradation of PVA). The effects of carbon and nitrogen sources for PVA degradation were also identified. The results revealed nitrogen sources significantly increase PVA-degrading enzyme production (83 U/mL enzyme activity). Furthermore, culturing E. cloacae MBB8 in M9 minimal medium containing 1% PVA accumulated polyhydroxyl butyrate (PHB) inside the cell. The PHB was purified and characterized by UV-spectroscopy. Further, the PHB was chemically converted into monomer and studied using gas chromatography coupled mass spectrometry (GC-MS). Overall, our results suggest that E. cloacae MBB8 is capable of degrading PVA and accumulate PHB that could be exploited industrially.
Introduction
Biorefining is defined as the process of producing multiple essential products from biomass or raw materials that are organic in nature, as opposed to petroleum hydrocarbons, industrial sludge and similar waste materials. 1 Various bioproducts including fuels, 2 energy, 3 materials, 4 and chemicals 5 from biomass are constantly being developed by biorefining processes. Polyvinyl alcohol (PVA) is a synthetic vinyl polymer that is biocompatible, toxicologically well-characterized, soluble in water and has excellent physical properties. 6 PVA is thermoplastic, and its main chains are connected only by carbon-carbon linkages. 7 –9 PVA is widely used to make water-soluble carriers, emulsions, adhesives and paper coatings. 9 –11 PVA is also a well-established polymer for film coating, and is used in pre-mixed coating formulations in pharmaceutical products.
However, PVA accumulation causes ecological contamination, and given its increased use in various applications, efforts are underway to develop a biological treatment for wastewater containing PVA. 12,13 At present, PVA is degraded using physical-chemical and biological methods. 14 PVA degradation using physical-chemical or wastewater-treatment techniques have several drawbacks, 15,16 such as production of toxic secondary pollutants, low degrading efficiencies, and high costs. 13 However, microbial degradation of PVA has numerous advantages; the process is non-toxic to the environment and economically friendly. 16 Microbial PVA degradation has attracted much attention in recent years and is considered to be a best alternative method to solve this problem. Microorganisms such as Stenotrophomonas rhizophila QL-P417 Bacillus niacin, 16 and Stenotrophomonas sp. SA2118 have been reported to have PVA-degrading capabilities.
In this work, PVA-degrading bacterium Enterobacter cloacae was isolated from the Gulf of Mannar, in Tamil Nadu, India. This is the first report on this genus discussing the PVA-degrading efficiency. The various factors, such as pH, temperature, and incubation time, that could affect the PVA-degradation efficiency of E. cloacae MBB8 were also examined. Furthermore, we show the PHB production ability of E. cloacae MBB8 in the presence of PVA in culture medium. The results of this work may contribute to efforts to produce valuable products while degrading industrial contamination in wastewater treatment.
Materials and Methods
ISOLATION AND CULTURE OF PVA-DEGRADING STRAINS
The marine samples were collected from the Gulf of Mannar (Tamil Nadu, India). Microbes were isolated after a peptone-enrichment culture by suspending 1 mL of a sample into 9 mL of NaCl solution (0.8%). The samples then were incubated at 28°C for 24 h with 135 rpm agitation on a shaking incubator, spread on screening agar plates using PVA as the sole carbon source, and incubated at 30°C for 48 h. 8 mL of I2-KI was added to the plates and reacted for 20 min in the dark. Colonies surrounded by a clear and large halo were selected. The M9 minimal medium was used for screening of PVA-degrading microorganisms. The composition of the M9 minimal medium (per L) was as follows: 1.0 g PVA, 1 g yeast extract, 33.9 g/L Na2HPO4, 0.05 g MgSO4·7H2O, 15 g/L KH2PO4, 5 g/L 0.05 g CaCl2 and 0.02 g FeSO4·7H2O, with the pH adjusted to 7.5. The 1.5% agar was used for the solidification of the screening medium when required. Further, the isolated strains were regularly cultured aerobically in Luria-Bertani medium broth (containing 0.5% yeast extract, 1% tryptone, 1% NaCl) at 30°C with 135 rpm agitation on a shaking incubator. For each strain, 1 mL of the culture was transferred to a 250-mL flask containing 50 mL of M9 minimal medium and then cultured at 30°C and 135 rpm for 48 h in an orbital shaker equipped with incubator (Scigenics, Tamil Nadu). The culture growth was monitored by measuring optical density of the culture at 600 nm via spectrophotometer (Jasco V-730, Japan). The PVA biodegradation ability of each isolate was examined on PVA agar plates containing a solution of iodine and boric acid. 19 The strain showing the maximum PVA degradation activity (MBB8) was selected for further characterization and experiments.
MOLECULAR ANALYSIS AND PHYLOGENETIC EVALUATION
The genomic DNA of MBB8 was isolated by the HiPurATM DNA purification kit. The 16S rRNA gene sequence from E. cloacae MBB8 was amplified using genomic DNA as a template for polymerase chain reaction (PCR). Two oligonucleotides (27F,5′-AGAGTTTGATCGTGGCTCAG-3′; 1492R, 5′-TACGGTTACCTTGTTACGACTT-3′ ) were used for PCR in a thermal cycler (Eppendorf, Mastercycler, Germany), with the following program parameters: pre-denaturation 95°C, 4.0 min; 30 cycles at 95°C for 1.0 min, 55°C for 1.0 min, and 72°C for 90 s. After the completion of the cycles, an extension at 72°C for 10 min was added to ensure the complete product amplification. The amplified PCR product was then purified using a gel extraction kit and was confirmed by commercial DNA sequencing. The 16S rRNA gene sequence was aligned with sequences deposited on the National Center for Biotechnology Information (NCBI) website by employing the program BLAST. A phylogenic tree was constructed using the neighbour joining method. 20,21
NUCLEOTIDE SEQUENCE ACCESSION NUMBER
The 16S rRNA gene nucleotide sequence has been deposited in the GenBank databases under accession number MT138639.
EVALUATION OF PVA DEGRADATION ABILITY
E. cloacae MBB8 was cultivated in medium containing PVA with or without yeast extract. The initial concentration of PVA was 1% or 10%. Residual PVA in the culture broth was measured by iodometry based on the green color produced by the reaction of PVA with iodine in the presence of boric acid by a previously described method. 22 The residual PVA in the medium and the growth rate of E. cloacae MBB8 cells were monitored by UV-visible spectrophotometry (optical density at 600 nm). The percentage of PVA degradation of cells at 24 h in medium containing 0.05% yeast extract was determined in cultures with different initial PVA concentrations. 18
ESTIMATION OF PVA BY UV-VISIBLE SPECTROPHOTOMETRY
Culture aliquots were collected at various intervals and were centrifuged at 13,000 rpm for 10 min. After centrifugation, cells were washed twice with deionized water and dried to a constant weight in an oven at 105°C. The biomass is expressed as grams dry cell weight per L (g DCW L−1). The PVA concentration was estimated using a previously described spectrophotometry method. 18 Briefly, 1 g/L of a standard PVA solution was diluted in a solution containing 0.5 mL of distilled water, 0.75 mL of 4% boric acid, and 0.15 mL of I2-KI (12.7 g/L I2 and 25 g/L KI). The final volume of the solution was diluted to 2.5 mL with distilled water and was equilibrated for 30 min at room temperature. Finally, the PVA concentration was analyzed on a UV spectrophotometer at 690 nm, yielding a standard curve that was linear from 0–100 μg/mL. The PVA concentrations in supernatants were measured after being properly diluted.
DETERMINATION OF PVA-DEGRADING ENZYME ACTIVITY
The PVA-degrading enzyme activity was measured according to the procedure outlined by Chen et al. 23 Briefly, 10 mL E. cloacae MBB8 culture (drawn at various time points in growth curve) was centrifuged at 12,000 rpm at 4°C for 10 min using Eppendorf 5804R centrifuge (Eppendorf AG®, USA). The culture supernatant was then filtered through a membrane filter (pore size, 0.45 μm) (MF-Millipore™, Merck KGaA, Germany) and dialyzed overnight in a solution containing 0.1 mol/L phosphate buffer (pH 8.0) to obtain crude enzyme solution. The remaining cells were washed twice and resuspended in 10 mmol/L potassium phosphate buffer (pH 8.0). Cells were disrupted in an ice bath using an ultrasonic processor followed by centrifugation at 10,000 × g for 30 min at 4°C. Subsequently, the sample was filtered through a membrane filter (pore size, 0.22 μm) (MF-Millipore) and was used as crude enzyme solution. The PVA-degrading enzyme activities in the culture broth were examined by the absorbance at 690 nm. 23 The reaction mixture containing 1 mL of crude enzyme solution and 100 μL of 0.1% PVA dissolved in 0.1 mol/L phosphate buffer (pH 8.0). Absorbance decreased at a rate of 0.001 per minute at 690 nm, which was considered 1 unit of PVAase activity. All experiments were performed in triplicate, and the data represent the mean values of results from three independent experiments.
PVA DEGRADATION ASSESSMENT BY FT-IR SPECTROSCOPY
A 50-mL culture was centrifuged at 13,000 rpm for 20 min at 4°C, and the culture supernatant was then filtered through a membrane filter (pore size, 0.22 μm, Millipore) and air dried. 23 PVA in the culture supernatant was measured using IR spectra acquired through FT-IR spectrometer (Jasco, Japan). IR Experiments were performed at room temperature, and the signals in spectral region from 400 to 3,900 cm−1 were recorded. 18 Decrease in the PVA concentration in culture supernatant was calculated by monitoring the decrease in the intensity of peaks (normalized) at 1,435 cm−1 (C-H bending) and 3,350 cm−1 (O-H stretching). 24
EXTRACTION OF POLY HYDROXYBUTYRATE (PHB)
Cells were cultured in the M9 minimal medium (containing 1% PVA) at 30°C with 135 rpm agitation for 48 h. After 48 h, extraction and estimation of the PHB were performed according to the procedure described earlier with slight modification. 25 Briefly, the biomass was pelletized by centrifugation (10,000 × g, 15 min) followed by digestion of biomass by incubating it with 0.5 M sodium hypochlorite at 40°C for 1 h. The lipid granules were deposited by centrifugation, followed by sequential washing using water, acetone, and ethanol. PHB from the residual components was extracted using hot chloroform (40°C). Finally, chloroform was evaporated to dryness at room temperature. The PHB polymer thus obtained were stored at 4°C in parafilm sealed container until further use.
DERIVATIZATION AND CHEMICAL CHARACTERIZATION OF PHB
The PHB obtained was converted to crotonic acid by acid hydrolysis. Briefly, 1 mg of the PHB obtained was added to 2 mL of concentrated H2SO4 in a capped test tube and heated for 20 min at 100°C in a water bath. At the end of incubation time, the mixture was cooled to room temperature. The absorbance of the sample was measured by Jasco V730 UV spectrophotometer at 235 nm against a sulfuric acid blank. The calibration plot was prepared with 3-hydroxybutyric acid (Sigma-Aldrich) as standard. The concentration of PHB in the test solution were calculated from the absorbance value using the standard graph. PHB was quantified as the percentage composition of the DCW (dry cell weight). The residual mass was calculated as the difference between DCW and PHB. The PHB thus purified was further analyzed using gas chromatography coupled with mass spectrometry (GC-MS). 25
MICROSCOPIC EXAMINATION OF PHB BY SUDAN BLACK B STAIN
The MBB8 culture was inoculated in M9 media and maintained at 37°C with 120 rpm agitation for 18 h. The culture media was centrifuged at 8,000 rpm for 30 min to separate the cells from supernatant. The cell supernatant was washed thrice with PBS buffer (pH 7.2). Several drops of bacterial broth were fixed on a glass slide by applying heat, further stained with a 3% Sudan Black B (w/v in 70% ethanol, Himedia) solution, and incubated for 10 min. The stained slide was then immersed in xylene until completely decolorized. The sample was then counterstained with safranin (5% w/v in Milli-Q water, Merck) for 20 seconds, washed again with distilled water, and dried. Subsequently, the immersion oil was added to the slides, the cells were screened by 1,000 X magnification using phase contrast microscopy (Leica, Tokyo, Japan). 26
GC-MS ANALYSIS FOR PHB
To evaluate the presence and abundance of PHB, the polymer sample was methanolyzed using sulfuric acid and methanol, as described by Bhuwal et al. 27 Briefly, 5 mg of the polymer sample was dissolved in 2 mL of chloroform, and then 2 mL of methanol was added and acidified with 3% (v/v) H2SO4 and heated at 100°C for 3.5 h for depolymerization and methanolysis of polyesters. The methanolysis derivative product was used for determination of PHB by gas chromatography. An Agilent 7890B gas chromatograph with 5977B MSD detector was used. 1 μL of PHB sample was injected in the ratio of 50:1 using Agilent auto-sampler into the back inlet with source temperature at 120°C. The separation was carried out in HP-88 capillary column (100 m length x 0.250 mm width) and 0.20 μm film with helium as carrier gas at a constant flow rate of 1 mL/min. Initial temperature was 120°C and hold time was 1 min, followed by increasing to 175°C at 10°C ramp rate with 10 min hold time, again increasing to 210°C at 5°C ramp rate with 5 min hold time, and finally increased to 230°C at 5°C ramp rate with 5 min hold time, completing the cycle of 37 min 50 sec. The separated peaks were analyzed using Agilent 5977B mass spectrometer.
Results and Discussion
PVA-degrading bacteria were qualitatively screened from Gulf of Mannar region seawater samples. Eight colonies were obtained after incubation of the sea water sample on M9 minimal medium containing PVA as the sole carbon source for 2 d (28°C). The attained individual colonies were transferred onto a M9 minimal medium agar plate and grown at 28°C for 2 d. Formation of clear halo surrounded by colonies indicated the presence of PVA-degrading bacteria. Among them, one colony (MBB8) was chosen for further study. MBB8 showed 83% PVA degradation in the culture medium after 2 d. Further, the strain taxonomic position was identified by biochemical and 16S rRNA gene sequencing analysis. The strain MBB8 formed pale white color colonies on MM agar and was a Gram-negative, motile, and rod-shaped bacterium. The strain was positive for catalase, glucose utilization, starch hydrolysis, and Voges-Proskauer and negative for indole, methyl red, lactose, and gelatin hydrolysis. The strain evolutionary distance was identified by the 16S rRNA gene sequencing and phylogenetic analysis. According to the BLAST analysis results, the 16S rRNA gene revealed that a high degree (99%) of identity with different strains of E. cloacae. Figure 1 shows the phylogenetic position of E. cloacae MBB8. Further, we have performed the carbon and nitrogen source optimization for PVA degradation. Our results revealed that the nitrogen sources have a significant increase in PVA-degrading enzyme production (83 U/mL enzyme activities, data not shown).
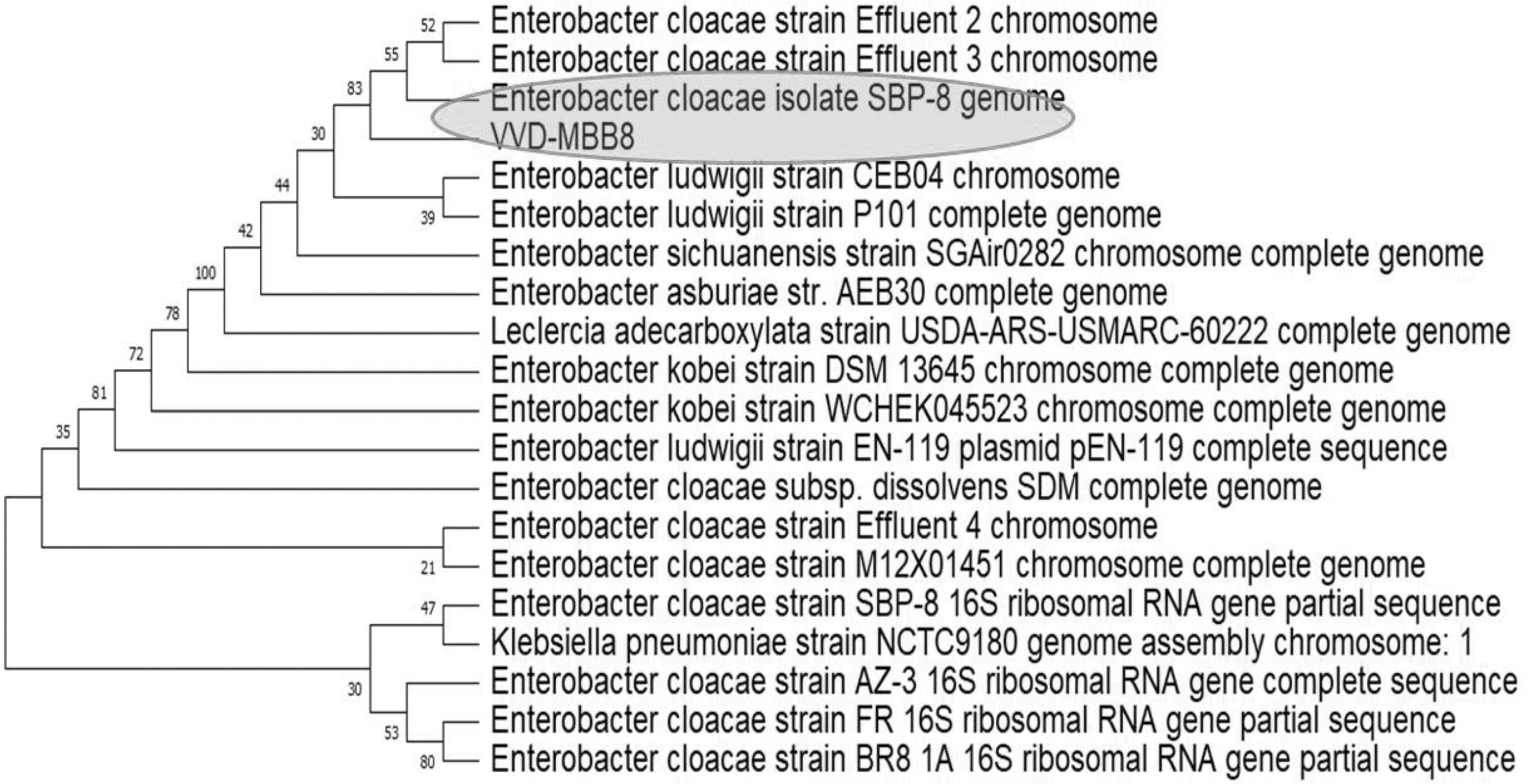
Phylogenetic analysis of E. cloacae MBB8 (highlighted in green) and related taxa using 16S rRNA gene sequences.
The UV chromatograms of the samples were taken before (0 h) and after degradation (24 h) process for 1 and 10% PVA sample in the broth culture medium (Fig. 2). LB broth was used as reference baseline chromatogram to avoid interference in the signal from the components of culture media. Figure 3a depicts the UV absorption spectra of 1% PVA before and after degradation process. It can be noted that in the UV chromatogram of 1% PVA, there was peak broadening at 375 nm that continues up to 410 nm and tails until 650 nm. A similar effect was observed in the chromatogram of 10% PVA before degradation. It is to be noted that this broadened peak is absent in the LB broth, therefore it can be assumed that the peak broadening is due to the reactants. However, after degradation the peak broadening at 375 nm is greatly reduced in 1% PVA and completely absent in the 10% PVA (Fig. 3b). This indicates that the organism effectively degrades the PVA in the medium within 24 h.
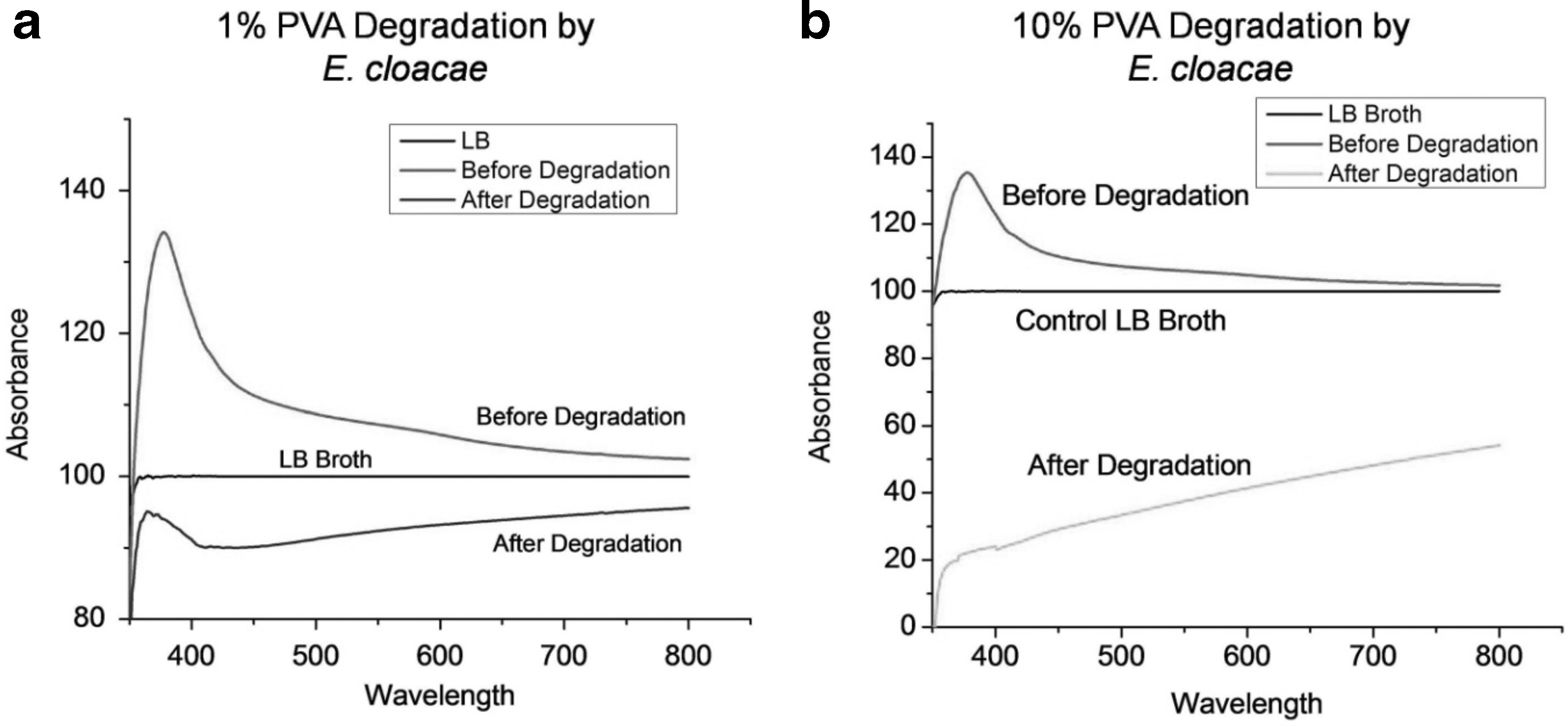
UV-Visible spectrogram of medium supernatant recorded before and after degradation of PVA by E. cloacae MBB8. LB broth (without PVA in the medium) was used as control during the UV-Visible spectroscopic measurements;
An FTIR spectral analysis was performed to investigate the structural changes of PVA in E. cloacae MBB8 culture supernatant during PVA degradation process at various time points viz 0, 24 and 48h. IR spectra were obtained in a wave number range from 650 to 4,000 cm−1 (Fig. 4). The characteristic absorption peaks of PVA were observed in the IR spectrum of 0 h (control). The broad peak at 3,300 cm−1 is due to the OH stretching in PVA (Fig. 4a). The major peaks that were observed during the process were at 710, 960, 1,027, 1,120, 1,200, 1,310, 1,420, 1,500, 1,650, 2,100, 2,900, 3,000 and 3,300 cm−1. The absorption at 710 and 960 are due to the out of plane C-H stretching and vinyl C-H out of plane bending, respectively. The peak at 1,027 can be attributed to in-plane C-H bending. The vinylene and vinyl C-H in-plane bending were observed at 1,310 and 1,420 cm−1, respectively. Further, the peak at 1,500 can be attributed to C-C stretching. The CH2 bending in PVA is observed at 1,425 cm−1 and the peak at 1,324 cm−1 might be due to C-H deformation (Fig. 4b). A relatively narrow, weak-to-moderate absorption, around 1,650 cm−1, is indicative of olefinic unsaturation. Either the high thermal treatment of PVA (up to 121°C for 30 mins) could have yielded in the formation of olefinic bonds in PVA or fatty acids such as maleic acid, lauric acid, or unsaturated fatty acids in the bacterial culture could have cross linked with PVA, resulting in the formation of olefins (C = C). The spectrum in the region of 2,700–3,500 cm−1 shows medium appearance of vinyl C=CH2 and C=CH in PVA at 2,975 and 3,020cm−1, respectively. Single bonds usually exhibit free rotation due to non-constraint over the spatial orientations for the molecules. However, constraint in the spatial orientation in PVA will have an impact on the IR spectrum. Furthermore, the preferred molecular orientations will have an impact on the physical and spectral properties of the material. The FT-IR spectrum of the PVA extracted from the supernatant after 24 and 48 h showed the similar characteristic peak but at reduced intensity. Thus it can be interpreted that the concentration of the PVA that was present in the culture supernatant had decreased. The PVA was degraded into other products, such as 4-hydroxy 2-pentanone or 3-hydroxybutyrate. Notably, these smaller products in the culture supernatant were either removed during the methanol precipitation process while preparing the sample for FT-IR or taken up by the bacteria inside the cell for metabolic processes, therefore the spectral properties relating to these compounds were less apparent in the IR spectrum other than the 1,650 cm−1 peak for C = O.
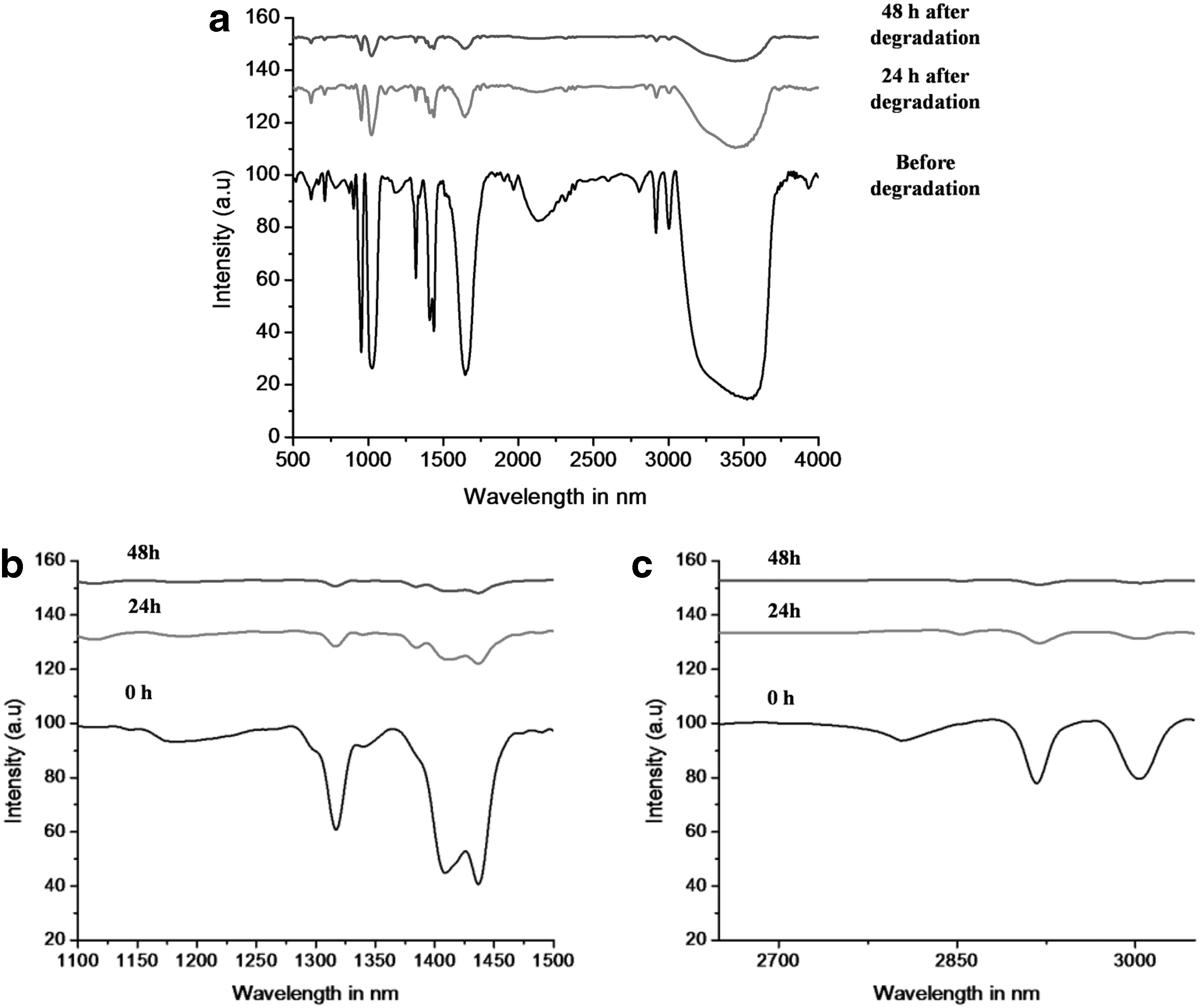
IR spectra of E. cloacae MBB8 culture supernatant obtained during PVA degradation process at various time points viz 0, 24 and 48 h;
There are two possible reaction schemes. The first is by the action of oxidase, and the second is through the action of dehydrogenase. The E. cloacae MBB8 could take either of the two routes (Fig. 5). Earlier reports have suggested the cooperation of pyrroloquinoline quinine (PQQ)-dependent PVA dehydrogenase (PVADH) and a hydrolase during the course of PVA degradation. 28 If the dehydrogenase acts on PVA, then the two hydroxyl groups on the 1,3-diol of PVA were converted to a β-diketone directly. On the other hand, if oxidase comes into play then either one of the two diols at 1,3 positions were converted into ketone, followed by the other. Thus, oxidase converts the 1,3-diol on PVA in a two-step process to form a β-diketone by the sequential oxidation of hydroxyl groups. The product thus formed is subjected to hydrolysis by hydrolase and results in the formation of terminal methyl and carboxylic acid group in the polymer chain. However, a more extensive analysis of the enzymatic chain-cleavage reaction will be needed to ascertain the scheme of the reaction in E. cloacae MBB8. It is not clear whether the β-hydroxy ketone moiety of the PVA form product other than β -diketone by the action of some other enzymes in the course of the biodegradation. If the PVA can be converted to 3-hydroxy butyrate, then this strain could potentially produce PHB upon proper experimental conditions.
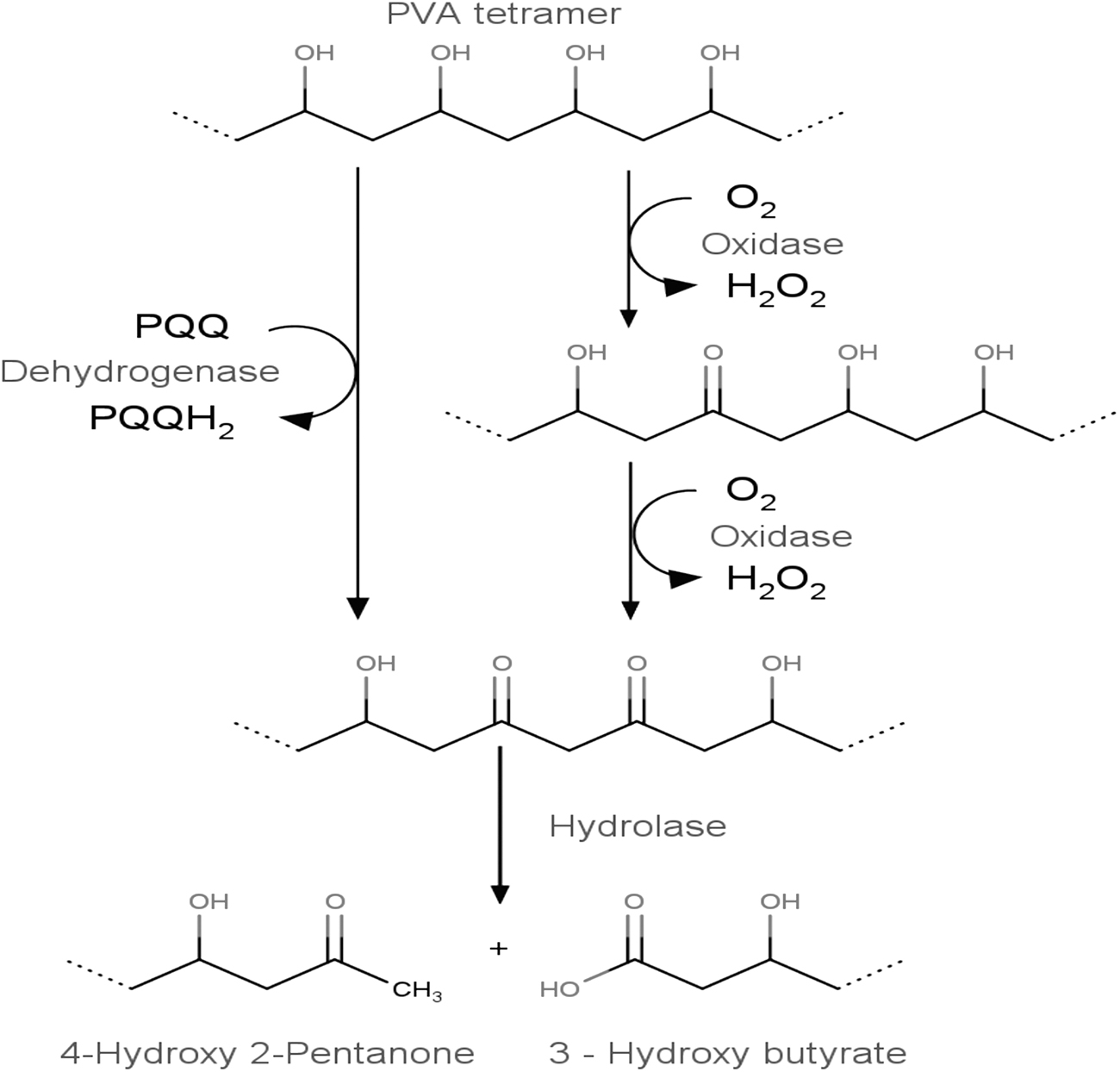
Schematic representation of enzymatic degradation mechanism of PVA by E. cloacae MBB8
At this juncture, we were unclear about the PHB-production capability of E. cloacae MBB8. However, the presence of 3-hydroxy butyrate in culture supernatant warrants investigation into PHB production and accumulation. If E. cloacae MBB8 was able to produce PHB by degrading PVA, then the inference may provide greater insights into the biochemical relationship of polymer degradation and PHB accumulation in microbes. It was our hypothesis that under proper experimental conditions E. cloacae MBB8 in M9 minimal medium containing 1% PVA can produce 3-hydroxy butyrate, that would be further converted to PHB and accumulated inside the cell. To test this hypothesis, we cultured E. cloacae MBB8 in M9 minimal medium containing 1% PVA for 48 h. The PHB accumulation inside the cell was initially verified by Sudan black staining method (Fig. 6a). The presence of blue inclusion bodies inside the cell may indicate possible accumulation of PHB. However, Sudan black also stains lipids and lipoproteins, therefore the qualitative method can be an indicator of possible PHB accumulation. The cells were centrifuged and the pellets were disrupted to purify PHB. The PHB thus obtained was characterized using UV-spectroscopy (Fig. 6b). To chemically characterize the PHB, it was converted to crotonic acid by acid hydrolysis using concentrated H2SO4 and analyzed using GC-MS (Fig. 6c). The reaction scheme for PHB conversion is provided in Fig. 6d. The mass fingerprint analysis of the GC-separated PHB indicated the presence of crotonic acid fragments at m/z 86.1, 71.1, 57.1, and 53.1, indicating the fragmentation pattern derived from m/z 102.07 (Fig. 6e).
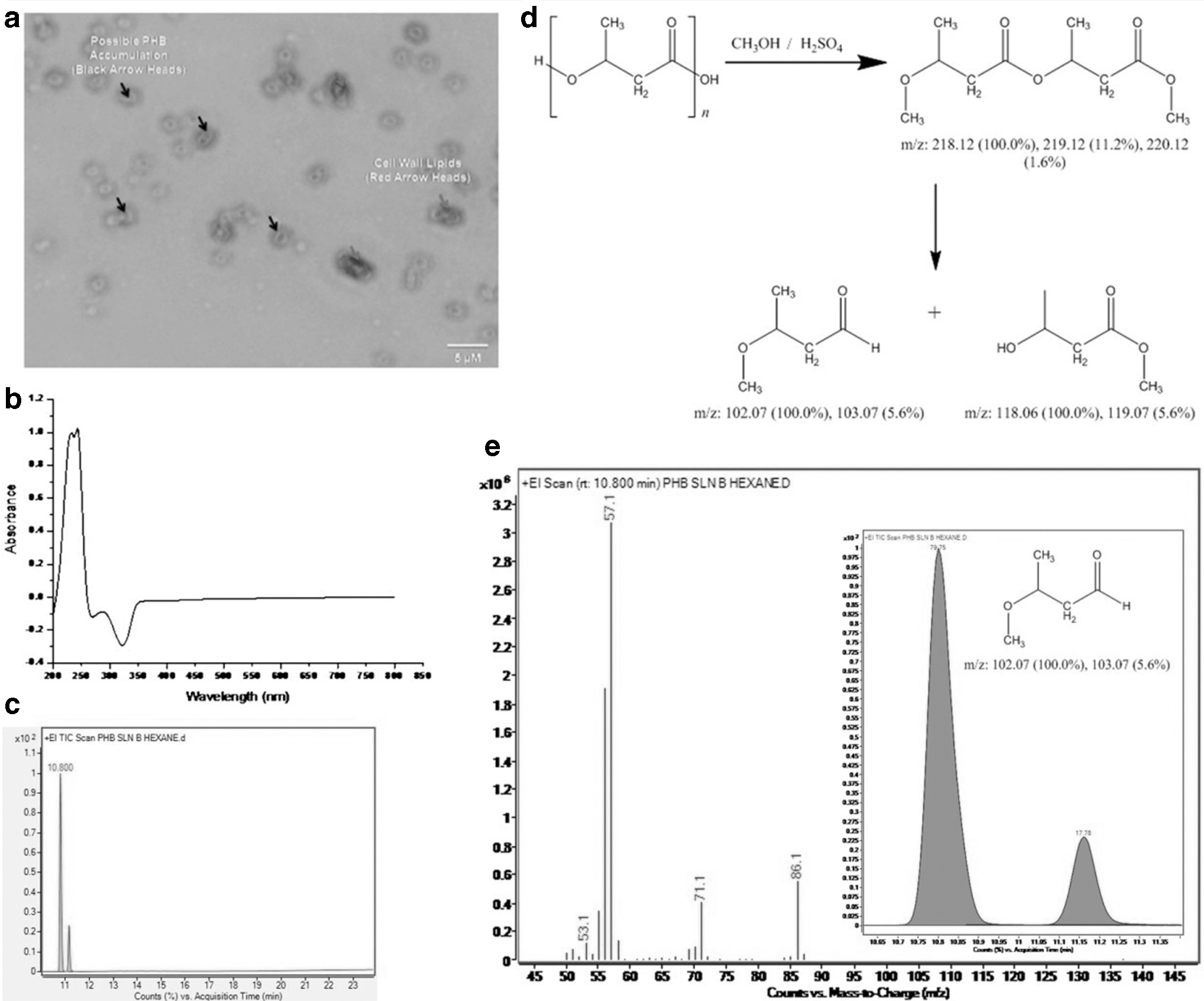
Characterization of PHB produced by E. colacae MBB8;
The optimum growth and the maximum PHB accumulation by E. cloacae MBB8 cultured in M9 medium containing 1% PVA occurred at 48 h. The results indicate PHB production and growth were concomitant with growth conditions. As biomass of E. cloacae MBB8 increased, PHB accumulation also increased, attaining maximum at 48 h (at peak biomass production). At 72 h, the cellular PHB level begins to decrease. This might be due to nutrient depletion, which forces the bacteria to use the accumulated PHB as an energy source.
Large-scale production of PHBs is largely hindered by the lack of a reliable, low-cost, large-volume feedstock. Employing a biorefining approach is interesting, as it utilizes/recycles the environmentally abundant substrate for PHB synthesis using environmental stressor tolerant microorganisms, thereby reducing the operational cost in PHB synthesis. Listewnik et al 29 pre-calculated the biosynthesis of PHB in a plant producing 500 tons PHB/year and reported the economic feasibility of PHB production from methane at the rate of $7.64/€6.35/ Rs. 560 per kg. However, accounting for the downstream processing costs, the projection extrapolated to $15.1–18.3 (Rs.1, 107/kg). Herein, we have not performed the full-scale cost analysis due to our own limitation at laboratory-scale. However, our estimated economic feasibility of PHB production using Enterobacter at pilot scale would be under $10 per kg PHB (Rs. 729.95/kg). Our experimental results suggest the feasibility PHB production by E. cloacae MBB8 in M9 minimal media, even in the presence of 1% PVA.
Conclusion
A novel, PVA-degrading bacterium (E. cloacae strain MBB8) was isolated from Gulf of Mannar using PVA as the sole source of carbon. This strain could degrade 70% of the PVA present in the culture medium in 24–48h. The organism has the potential to produce PHB biopolymer from PVA. The use of PVA as the sole carbon source acts as the limiting factor in PHB production. The results of the present study suggest the need for a new paradigm in biopolymer production, degradation, and transformation. Comprehensively, E. cloacae MBB8 are potential PVA-degrading microorganisms with interesting industrial applications.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Ramya Venkatesan is a Prime Minister Research Fellow (PMRF). R.V. greatly acknowledges the Department of Science and Technology–Science and Engineering Research Board (DST–SERB) India, Confederation of Indian Industry (CII) and VVD and Sons Private Limited, Thoothukudi, Tamil Nadu for the DST (SERB)-CII Prime Minister Research Fellowship award (April 2018 Batch). Prakash Shyam Karuppiah, Rajkumar Prabhakaran, and R.V. immensely thank Department of Scientific and Industrial Research (DSIR) for recognizing V.V.D and Sons Private Limited as an Industrial Research and Development Unit. M.M thanks Udaya School of Engineering for the support to conduct the research and development activities.