
Abstract
Brazil has made significant progress in ethanol production from sugarcane juice. However, in terms of water, food, and environmental safety, this production can be optimized by replacing raw food with agricultural waste, giving rise to second-generation ethanol (2G). Among these biomasses, coconut residue stands out in tropical countries like Brazil. This study aimed to fractionate coconut husk through alkaline pretreatment with calcium hydroxide (20% w/w) followed by enzymatic hydrolysis with NovozymesTM Ctec2 and Htec2. The conversion efficiency of polysaccharides into monosaccharides was 19.3% for cellulose and 7.3% for hemicellulose. The inhibitors acetic acid (∼3.0 g/L) and formic acid (∼0.8 g/L) were also observed in the hydrolysate, to which 17.5 g/L NaCl was added to evaluate yeast cells under salt stress. The wild strain Wickerhamomyces sp. UFFS-CE-3.1.2 and the industrial yeast Saccharomyces cerevisiae PE-2 were inoculated into this non-detoxified high-salt hydrolysate and analyzed through a Central Composite Design (CCD). The independent variables were the effects of pH and temperature on yeast metabolism. Assuming cellular growth and sugar consumption as responses to the experimental design, pH of 7.0 and temperature of 40°C were defined as the best cultivation conditions for subsequent fermentation kinetics. As a result, the wild yeast showed the highest biomass production, but it did not produce ethanol. In contrast, the industrial strain fermented glucose, displaying an ethanol yield of 0.24 gethanol/gsugar. Thus, coconut husk can be promising biomass for 2G ethanol production, even though it requires pretreatment and hydrolysis optimization to increase sugar concentration and reduce inhibitors in the hydrolysate.
Introduction
The increase in energy demand and growing concerns associated with fossil fuels have stimulated the development of new technologies to produce renewable fuels. Among biofuels, ethanol stands out for its potential to increase sustainability and change the global economy towards a greener future. 1,2 Currently, the global production of ethanol is led by the United States, followed by Brazil. The two countries account for 84% of world production. 3
Although the production of first-generation (1G) ethanol is well established, the concerns with factors such as food security, water security, and the advance of farmland to areas of native vegetation made room for the development of second-generation (2G) ethanol. 4 However, lignocellulosic biomass used as raw material for 2G ethanol, unlike that used in the production of 1G ethanol, is rich in cellulose, hemicellulose, and lignin. Therefore, sugars are not readily available to be fermented. This means that two prior stages of pretreatment and hydrolysis are required, in which lignin is removed or degraded and the crystallinity of polysaccharides is broken. 5 Pretreatment has a significant influence on the cost of the ethanol production process, accounting for more than 40% of the total cost of production. 6 This step influences the conversion and hydrolysis rates and the necessary enzyme load. 1,7 Moreover, depending on the pretreatment technique employed, the breakdown of hemicellulose and cellulose release sugars such as xylose and cellobiose, which are not naturally fermented by the yeast commonly used in the industry, Saccharomyces cerevisiae. 2
The lack of microorganisms that metabolize these sugars and tolerate the stresses of industrial fermentation economically limits the production of 2G ethanol. 8,9 Faced with this limitation, different researchers have focused on the isolation and bioprospecting of new wild yeast strains capable of metabolizing the sugars present in hydrolysates and tolerating inhibitors of the fermentation process. 2,10,11 For instance, Bazoti and coworkers 12 showed that the yeast Wickerhamomyces sp. UFFS-CE-3.1.2 is able to produce ethanol even in the presence of inhibitors, such as acetic acid and furfural. This same strain showed total xylose consumption in the fermentation of sugarcane bagasse hydrolysate at high pH values. 13 Furthermore, Bonatto and colleagues 14 demonstrated that Wickehamomyces sp. UFFS-CE-3.1.2 showed a maximum ethanol yield (0.42 gethanol/gsugar) close to the theoretical maximum yield when fermenting papaya residues using seawater as solvent.
Different types of biomass can be used to produce 2G ethanol, such as green coconut husk. The green coconut industry is present in more than 200 countries, with a worldwide production of 61.8 million tons, of which 2.3 million tons are grown just in Brazil—the fifth-largest producer. 15,16 It is known that the structure of the green coconut consists of epicarp, mesocarp, endocarp, solid albumen, and liquid albumen and that only the solid albumen and the liquid are edible. 17 A coconut typically exceeds 1 kg when fresh, and the unconsumed parts account for half of that mass on a wet basis. 18 Therefore, it is essential to properly allocate the large volume of waste from this production to avoid overloading landfills and environmental pollution. 19
The green coconut husk is a lignocellulosic biomass consisting of 33.2% cellulose, 29.1% hemicellulose, and 25.4% lignin. 17,20 The high proportion of lignin present in coconut husk makes this biomass unfit for reuse in the paper industry. 18 On the other hand, the prominent percentages of cellulose and hemicellulose make coconut husk an attractive raw material for producing 2G ethanol. It is possible to perceive this potential when considering a fermentation efficiency of ∼90%, which is currently practiced in the 1G ethanol industry concerning the yield theoretical maximum (0.51 g of ethanol per g of sugar). 21 Given the production of green coconut in 2018 and imposing the conditions that cellulose is composed of glucose and hemicellulose is composed mostly of xylose (which gives a total of about 19.27 million tons of sugars), it would be possible to obtain 12.39 billion liters of ethanol from the reuse of this biomass. This is a significant amount and is equivalent to ∼24% of the ethanol produced by the United States in 2020 (52.3 billion liters). 3
The concern with freshwater use in the ethanol production process is growing. For each gallon of 1G ethanol produced from corn, 3 to 4 gallons of water are spent. 22,23 For 1G ethanol from sugarcane, it is estimated that 22 m3 of freshwater is spent for each ton of sugarcane, and 10% is spent only on the washing of sugarcane. 24,25 Meanwhile, the greater number of operations required for the 2G process makes water consumption even more expressive. According to Scapini and coworkers, 23 most pretreatment techniques demand large amounts of freshwater. This is because water is used to adjust the solid:liquid ratio of the process and avoid yield losses. In addition, there is a need for washing after pretreatment in order to recover solubilized sugars and remove possible inhibitors, which increases the use of freshwater. 24 Thus, the use of seawater in the stages of ethanol production is an attractive approach to reduce the water footprint of the processes as well as identifying halotolerant enzymes and microorganisms capable of tolerating high osmotic pressures. 23,26,27
This work aimed to analyze the reuse of the green coconut shell for the production of 2G ethanol by producing a hydrolysate from alkaline pretreatment and enzymatic hydrolysis. Moreover, aiming to reduce freshwater use in ethanol distilleries and, consequently, the water footprint of the process, the coconut hydrolyzate was added with a NaCl concentration equivalent to half that measured in seawater. 28 This high-salt concentration hydrolyzate was alternatively inoculated with two yeasts of different origins: the industrial Saccharomyces cerevisiae PE-2, isolated from a 1G ethanol Brazilian distillery, and the wild Wickerhamomyces sp. UFFS-CE-3.1.2, isolated from rotten wood in the Atlantic rain forest. 29,30
Materials and Methods
Raw Material
The biomass used in the study was the coconut husk (epicarp and mesocarp), acquired in the local market of Chapecó, SC, Brazil. It was dried in an oven, between 60 and 80°C for 48 h. The coconut husk was milled in a Willey knife mill (AL-032S). Through the particle size analysis, it was determined that the size of the particles employed in the study would be up to 0.25 mm. 31
Characterization of Lignocellulosic Biomass
The physicochemical characterization of biomass, moisture, 32 ashes, 33 extractives, 34 and lignocellulosic composition 35,36 were determined following the National Renewable Energy Laboratory (NREL) protocols.
Moisture was determined from a 6.0 g sample maintained at 105°C to constant weight (about 24 h) and weighed to calculate the dry weight. 32 Ash percentage was estimated from a 1.5 g sample maintained at 800°C for 2 h and weighed to measure the ash content. 33 In order to evaluate the extractive content, filter paper cartridges with 4.0 g of biomass were prepared. These cartridges were subjected to Soxhlet extraction with water and later with ethyl alcohol. 34 After the end of the process, the cartridges were dried to constant mass and weighed to calculate the percentage of extractives.
To evaluate lignocellulosic composition, flasks with 3.0 mL of sulfuric acid (72% w/w) for each 0.3 g were arranged in a thermal bath for 1 h at 30°C, being homogenized periodically. 35 84 mL distilled water was then added. To provide complete hydrolysis of the oligomers, the flasks were autoclaved (1 h, 1 bar, at 121°C).
At the end of the process, the solid and liquid phases were separated by vacuum filtration. The liquid phase was used to quantify soluble lignin, carbohydrates, acetyl groups, furfural, and HMF. These chemicals were quantified through HPLC analysis (see below).
The lignocellulosic composition was determined through acid hydrolysis (sulfuric acid 72% w/w) in a thermostatic bath at 30°C for 1 h, followed by autoclave (1 h, 1 bar, at 121°C). The percentages of lignin (soluble and insoluble) and the proportions of cellulose (Equation 1) and hemicellulose (Equation 2) were estimated according to the concentrations of sugars (glucose, xylose, and cellobiose) and decomposition products (HMF, furfural acetic acid). The factors of conversion scans used were described by Rabelo. 37

where: Ce = cellobiose; G = glucose; HMF = 5-hydroxymethylfurfural; FA = formic acid; V = final filtration volume; E = extractives; B = biomass without extractives; X = xylose; F = furfural; AA = acetic acid.
Pretreatment and Enzymatic Hydrolysis
Alkali pretreatment was performed as described by Tadioto et al. 38 The experiments took place in flasks with 10 g of coconut husk and 100 mL of calcium hydroxide solution (20% w/w). The flasks were taken to the shaker incubator (SL-223) at 70°C and agitated at 200 rpm for 24 h. After pretreatment, the flasks were removed from the shaker incubator to reduce the temperature and pH adjustment. The pH was adjusted with citric acid (1M) at 5.0–5.5 (pH optimal for enzymes) to start enzymatic hydrolysis. Enzymes CELLIC® CTec2 and CELLIC® HTec2 were used. They were diluted in acetate buffer at concentrations (concerning dry biomass mass) of 2% for HTec2 (28 FPU) and 0.5% for CTec2 (113 FPU) and added to the flasks.
The enzymatic hydrolysis process occurred afterward. The flasks were returned to the shaker incubator for 24 h, at 50°C and 200 rpm. After the enzymatic hydrolysis, the hydrolysate was neutralized with calcium carbonate to reach a pH of 5–6. Subsequently, aliquots of neutralized hydrolysate were filtered using 0.22 μm membrane filters to determine sugars.
Analytical Methodology
Glucose, xylose, cellobiose, acetic acid, formic acid, and ethanol concentrations were quantified by high-performance liquid chromatography (HPLC; LC-MS 2020, Shimadzu) equipped with a refractive index detector RID-10A and an Aminex column HPX-87H (Bio-Rad). Samples were chromatographed at 50°C, with 5.0 mM sulfuric acid, as mobile phase, and flow rate of 0.6 mL/min. 39 Furfural and 5-hydroxymethylfurfural concentrations were determined using an SPD-M20A detector operated with an NST-18 column, eluted with 85:15 acetonitrile/water and 1% acetic acid, at 40°C and flow rate of 0.8 mL/min. 40 Before HPLC analyses, the samples were filtered using nylon filters with 0.45-μm pores.
Adjustment of Sugars in the Hydrolysate
Since the hydrolysis step rendered a low concentration of fermentable sugar, the green coconut hydrolysate had its glucose and xylose concentration adjusted to 25.0 g/L each. 3.0 g/L of monobasic potassium phosphate was also added for micronutrient adjustment, and 17.5 g/L of sodium chloride was added to evaluate the effect of osmotic stress imposed by the presence of this salt. 14,28 The hydrolysate had the pH adjusted according to the values of the experimental planning and was sterilized by vacuum filtration, using nylon filters with 0.22 μm pores.
Microorganisms
Two yeast strains were used in the study: UFFS-CE-3.1.2 and PE-2. The strain UFFS-CE-3.1.2 was previously isolated by this group from rotten wood samples (collected in Chapecó, SC, Brazil) and taxonomically identified as a new species, not yet described, of the genus Wicherhamomyces. 30 The strain PE-2, used for comparative purposes, is one of the leading industrial yeasts of the species Saccharomyces cerevisiae used in Brazil's 1G ethanol production. 29 This strain was kindly provided to us by Fermentec (Piracicaba/SP).
Yeast Cultivation Following Central Composite Design (CCD)
The yeasts were pre-grown in YPD medium (10 g/L of yeast extract, 20 g/L of peptone, and 20 g/L of glucose) for 48 h, at 25°C and 145 rpm. A CCD varying pH and temperature (Table 1) was assembled with Protimiza Experimental Design software. 41
The experiments took place in Erlenmeyer flasks filled with hydrolysate to one-fifth of their volumes. The yeasts were inoculated, and the flasks were kept under constant agitation (150 rpm). The temperature was adjusted according to the CCD for 48 h. At the end of the experiment, samples were collected to analyze cellular growth and sugar consumption. Cellular growth is presented in terms of biomass production (Equation 3). A spectrophotometer measured optical density at 570 nm. Culture supernatants were collected to evaluate the reducing sugar (RS) concentration through a microplate DNS method we previously described. 42
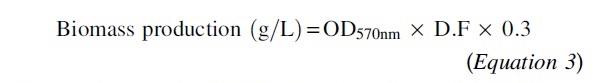
where: OD570nm = Optical density at 570 nm; D.F = Dilution factor; 0.3 = absorbance-to-yeast biomass conversion factor.
Fermentation Kinetics
After determining through CCD the best condition in terms of pH and temperature, fermentation kinetics were performed under constant agitation at 150 rpm. For this, the yeasts were pre-grown for 48 h (as described in the previous section) and inoculated in Erlenmeyer flasks filled with hydrolysate to one-fifth of their volumes. The experiments were carried out in triplicate (three independent experiments), and the standard deviations were always less than 10%.
Aliquots were taken at regular intervals. These samples were centrifuged for 5 min at 3,500 rpm and stored for further HPLC analysis (see above) to determine sugar consumption and ethanol production. In parallel, aliquots were harvested to evaluate cellular growth (described above).
Results
Biomass Characterization and Coconut Hydrolysate
The raw material showed a moisture content of 5.23%. The chemical composition of the biomass was evaluated on a dry basis and showed 26.26 ± 0.98% of extractives, 26.73 ± 0.99% of total lignin, 25.11 ± 0.74% of cellulose, 19.39 ± 0.94% of hemicellulose, and 2.51 ± 0.09% of ashes. The hydrolysate obtained from this residue showed 4.86 ± 1.10 g/L of glucose and 1.42 ± 0.36 g/L of xylose, and the presence of disaccharide cellobiose was not detected. As inhibitors, formic acid (0.76 ± 0.02 g/L) and acetic acid (3.03 ± 0.12 g/L) were observed. The concentrations of sugars found are equivalent to a conversion rate of 19.33% (w/w) of cellulose in glucose and 7.30% (w/w) of hemicellulose in xylose.
Experimental Design
The data for cellular biomass production and RS consumption are shown in Table 1. The initial RS concentration of the hydrolysate was 76.85 g/L, and this value was used to calculate the percentage of RS consumed in each assay. The CCD analysis showed that pH exerted a significant positive effect on both responses (cellular biomass and RS consumption) for both strains, indicating that the higher the pH value (considering the pH range analyzed), the higher the yeast performance. In contrast, the temperature did not significantly affect a significance level of α = 10%, despite calculated t being positive (Fig. 1). Therefore, the condition adopted for the subsequent fermentation kinetics for the two strains was the same as in experiment 4 (pH of 7 and temperature of 40°C, Table 1), which showed the higher cellular growth and sugar consumption.

Pareto charts for
Fermentation Kinetics
The wild yeast UFFS-CE-3.1.2 produced a negligible amount of ethanol from 6.72 g/L of glucose that the cells could consume during the incubation period (Fig. 2A). By contrast, the industrial S. cerevisiae PE-2 consumed 80.5% of glucose. It displayed maximum ethanol productivity of 0.17 g/L.h and a fermentation yield of 0.24 g of ethanol per gram of glucose consumed (Fig. 2B), representing 47% of the maximum theoretical ethanol yield (0.51 g/g). None of the strains were able to consume xylose. Although UFFS-CE-3.1.2 had a lower fermentation performance, it produced almost twice the cellular biomass as PE-2.

() Cellular growth, () ethanol production, and () consumption of glucose and (•) xylose during batch fermentation by
Discussion
Biomass Characterization
The characterization of biomass is indispensable for it to be considered an economically efficient energy resource because it assists in defining its application perspectives. 43 Biomass characteristics vary according to type, locality, climatic conditions, and cultivation soil. In the present study, the results were presented on a dry basis for better comparing to the literature.
The percentage of ashes we found is close to 2.45 ± 0.01%, which was seen by Araújo 19 for the same biomass, while for the husk of a mature coconut, this value is usually higher (3.31 ± 0.32%). 44 Soares and collaborators 17 found the percentage of 30.1 ± 0.02% of extractives in water, while Nogueira and collaborators 15 found 19.62 ± 0.39% using the same methodology. Therefore, the percentage of extractives found is among the literature already exposed.
The amount of lignin can be compared with the percentage of 25.4% presented by Soares and collaborators, 17 while holocellulose (hemicellulose + cellulose) was higher than that reported by Araújo et al., 19 of 38.03 ± 0.47%. This considerable percentage of carbohydrates (∼44.5%) demonstrates that the green coconut husk can be considered a potential source of fermentable sugars for 2G ethanol production.
Coconut Hydrolysate
The choice of pretreatment should be based on the characteristics of biomass. 45 Studies in the literature demonstrate the efficiency of using alkaline pretreatment for subsequent coconut hydrolysis. 18,19,46 The percentage of cellulose conversion found is close to that found by Araújo et al. 19 to the coconut husk with an alkaline pretreatment with hydrogen peroxide, followed by 72 h of enzymatic hydrolysis. The low conversion of hemicellulose in xylose presented in the present study may have occurred because xylan, a polymer found in more significant quantities in the plant cell wall, requires several enzymes, with different specificities against its heterogeneous complex chemical composition. 9 The saccharification of the main hemicellulose chain by endo-β-xylanases releases xylooligosaccharides of varying degrees of polymerization, which then may be hydrolyzed into xylose by β-xylosidase. 47 However, β-xylosidase activity is deficient in most commercial enzyme cocktails. This leads to the inefficient hydrolysis of hemicellulose and causes the accumulation of xylooligosaccharides, which can further reduce cellulase efficiency. 48 To optimize enzymatic saccharification, studies in the literature seek to optimize this step by microorganisms screening processes, 9,44,49 and by adding surfactants, proteins, and other polymers. 19,50,51
Experimental Design
Ethanol production from microbial fermentation represents 80% of the production of global renewable fuels. 2,52 It is estimated that around 1.388 to 9.812 L freshwater is spent for each liter of ethanol produced. 53 Aiming to reduce the water footprint of ethanol production, an equivalent to 50% of seawater NaCl concentration was added to the coconut hydrolysate to evaluate the osmotic tolerance of the yeasts studied.
The wild yeast showed the highest cellular growth under all tested conditions. This strain also had the highest RS consumption and stayed just behind the PE-2 in experiment 5 (Table 1). Data analysis also shows the significant positive effect of pH for both strains and both responses (cellular biomass and RS consumption). There was higher cellular growth when this variable was at its higher assayed level (Fig. 1). Indeed, Bonatto et al. 13 observed that fermentations with pH 7 favored glucose and xylose consumption by UFFS-CE-3.1.2, increasing ethanol production. In addition, a more alkaline pH helps in reducing the inhibiting effects of acetic acid. According to the authors, this inhibitor has its undissociated form easily diffused into the cell at acidic pH levels.
As a result of the difference in pH, acetic acid dissociates inside the cells for its ionic form, resulting in proton release and decreased cytosolic pH. Cytoplasm acidification damages the cell, directly reflecting lower sugar consumption rates and lower ethanol yield. By contrast, the acetic acid is already in its dissociated form at a higher pH, having more difficulty passing the plasma membrane. 54 –56
The temperature variable exerted no significant effect (Fig. 1). However, the best conditions for the biomass production of both strains occurred at higher temperatures. Indeed, especially for the wild yeast, the increase in this variable clearly caused an increase in RS consumption. Thus, the optimization of this variable is crucial due to the direct influence on the conversion of sugars, 57 directly affecting the cell metabolism. 2 Usually, the optimum temperature for yeasts is below 35°C. 49 However, species of the genus Saccharomyces and Kluyveromyces are known to display cell growth above 40°C. 58 The strain Kluyveromyces marxianus CICC 1727-5, for example, was able to produce higher concentrations of ethanol, from glucose, at 40°C. 59
Nonetheless, the highest ethanol production of this strain using xylose as substrate was at lower temperatures. High-temperature fermentation is desirable for ethanol production because it avoids bacterial contamination and reduces cooling costs. 2 Moreover, the search for thermotolerant yeasts may also enable increased efficiency in simultaneous saccharification and fermentation processes (SSF), excluding the need for a cooling system and, consequently, saving energy resources. 58 Therefore, the choice of pH for the subsequent fermentation kinetics was based on the preference of yeast cells indicated in Fig. 1. At the same time, the option for a higher temperature considered its relevance for lignocellulosic ethanol production.
Fermentation Kinetics
The strain PE-2 has been widely studied for its adaptability to industrial conditions with greater tolerance to stress and higher fermentative performance. 2,29,60 However, this strain cannot metabolize xylose, 61 as shown in Fig. 2B. However, despite the presence of inhibitors and NaCl, the industrial strain was able to produce ethanol from glucose. A study that employed the same PE-2 into a mature coconut fiber hydrolysate obtained maximum ethanol productivity of 0.30 ± 0.15 g/L.h and a fermentative efficiency of 85.17 ± 1.17% concerning the theoretical maximum ethanol yield. 62 This difference between the results may demonstrate the negative effect of NaCl on industrial yeast during our fermentation kinetics. It is worth noting the relevance of prospecting wild yeasts with osmotic tolerance to ensure the reduction of the water footprint of the process. In fact, Zaky and coworkers 53 observed that marine yeasts might perform equal to or higher than terrestrial, industrial strains under high osmotic stress conditions.
Bazoti et al. 12 demonstrated that the strain UFFS-CE-3.1.2 can metabolize xylose and produce ethanol, even in the presence of inhibitors. However, this strain did not show significant consumption of the analyzed sugars and did not produce ethanol satisfactorily. It is also observed that even minimally consuming glucose and xylose, UFFS-CE-3.1.2 showed higher biomass production than the PE-2 strain. This may indicate the presence of other non-analyzed carbon sources present in the coconut hydrolysate, as evidenced by the initial RS concentration (76.85 g/L).
Soares and coworkers 17 observed the presence of mannose and arabinose in coconut hydrolysates. Also, according to the raw material and the enzymatic cocktail used, some enzymatic hydrolysis products of xylans may remain as oligosaccharides. 63,64 According to Qing and Wyman, 65 the complete saccharification of xylooligosaccharides, in a typical period of 72 h, using only moderate doses of commercial enzymes, without supplementation with β-xylosidase, is an arduous process. Considering the residence time used in our study (24 h) and the use of a commercial hemicellulases cocktail, we may infer the possibility of xylooligosaccharides accumulation, which the wild strain could have consumed. These compounds may explain the initial RS concentration observed in the hydrolysate. However, the detection and separation of these compounds require other chromatographic analysis means, such as high-performance anion-exchange chromatography coupled with pulsed amperometric detection (HPAEC-PAD). 66
The presence of inhibitors was also a limiting factor for yeast performance. Studies show that acetic acid concentrations between 2.0–5.0 g/L can seriously inhibit most xylose-fermenting microorganisms. 67,68 Formic acid, typically present in lower concentrations in lignocellulosic hydrolysates, has an even higher toxicity than acetic acid. 69 The presence of these weak acids negatively affects the growth and metabolism of microorganisms, particularly xylose metabolism, thus reducing ethanol production. 70 The short-term acclimatization of yeasts to these acids during pre-cultivation can significantly improve xylose fermentation. 69 This process allows increasing tolerance to inhibitors through metabolic engineering, based on a response to a short period of stress.
Conclusion
The coconut husk is a potential raw material for ethanol production due to its high percentage of holocellulose that can be converted into fermentable sugars. However, optimizing the pretreatment and enzymatic hydrolysis processes is necessary to use this biomass better, achieving higher concentrations of fermentable sugars and lower concentrations of inhibitors. The wild yeast UFFS-CE-3.1.2, despite not producing ethanol under the imposed conditions, showed higher RS consumption and biomass production.
Footnotes
Acknowledgments
The authors thank Dr. H. V. Amorim (from Fermentec, Piracicaba/SP, Brazil) for kindly providing the strain S. cerevisiae PE-2.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported in part by grants and fellowships from the Brazilian National Council for Scientific and Technological Development – CNPq (Grant Numbers 454215/2014-2 and 305258/2018-4), the Santa Catarina State Research and Innovation Support Foundation – FAPESC (Grant Number 749/2016; T.O. 2016TR2188), and the Research Promotion Program from Federal University of Fronteira Sul – UFFS (Grant Numbers PES-2019-0638 and PES-2020-0213). AD received a scholarship from the Support Program for Scientific and Technological Initiation from the Federal University of Fronteira Sul (PRO-ICT/UFFS).