
Abstract
A novel thermostable protease, AprX from G. thermoleovorans ARTRW1, was recombinantly produced, purified and characterized. This work shows that the 85 amino acids from the N-terminal was cleaved post-translationally, indicating that the enzyme was synthesized as an inactive precursor “zymogen”. The molecular weight of the mature protein is 38.1 kDA. Prolonged incubation at different temperatures and time intervals showed that the protease caused self-cleavage above 45°C and AprX degraded completely within 6 h. Mass spectrometry analysis has shown that the enzyme has a partial preference to cleave after R, K and L residues similar to trypsin but it also cleaves after the C-terminal end of E, S, V, F, G, H, N and T residues. The enzyme activity reached maximum at 55°C and over a broad pH range between 5 and 11. The protease was found to be highly tolerant of detergents and completely inhibited by PMSF, Zn2+ and Ni2+, similar to trypsin-like serine proteases. It was stable from 30°C to 70°C, retaining 80% activity for 3 h at 55°C. This new protease could be a candidate for use in a variety of industrial processes that require long-term stability at elevated temperatures.
Introduction
Proteolytic enzymes are among the enzymes widely used in industry and constitute more than 60% of the world enzyme market. 1 Alkaline proteases, the most widely used class of all proteases, 2 are used in several areas such as food, textile, leather, waste management, cosmetics and pharmaceutical industries, especially in the detergent industry. 1,3 Alkaline proteases are both stable and active in the presence of surfactants and oxidizing agents, which are among the additives commonly used in detergent formulations. 2
There is great interest in microorganism-derived alkaline proteases for use in biotechnological applications because enzymes produced by microorganisms have high catalytic activity and do not cause undesirable byproducts. In addition, growth conditions of microorganisms can be easily optimized and large quantities of high purity enzymes can be produced in a single production process. 4 The first alkaline protease discovery to be used for the detergent industry was subtilisin Carlsberg from Bacillus licheniformis. 5,6 Subsequently, a number of alkaline proteases from microorganisms, including Aeribacillus pallidus, 7,8 Bacillus circulans DZ100, 9 Bacillus pseudofirmus, 10 Bacillus pumilus NRC21, 11 Geobacillus caldoproteolyticus,12 Geobacillus toebii Strain LBT 77, 13 and Shewanella arcticai, 14 were cloned, characterized and investigated for use in a variety of biotechnological applications such as detergents, leather degradation and waste management.
The fact that Bacilli species can be easily isolated from various sources has made these microorganisms a significant focus of biotechnology. 1 Most of the Bacilli species that have been isolated were thermophiles and have been shown to produce a protease with optimum activity above 60°C. 15 -17 Therefore, Bacillus species are excellent sources of thermostable proteases that can be used for pharmaceutical and industrial applications. There are hundreds of patents covering proteases produced by different species of Bacilli and their uses, including B. subtilis, B. licheniformis, B. pumilus and B. gibsonii. One of these proteases, tradenamed Biotex by Novozymes, produced by B. licheniformis. 18
In this study, a thermoactive alkaline serine protease protein AprX was recombinantly produced from a recently identified thermophilic bacterial strain, Geobacillus thermoleovorans ARTRW-1, 19,20 and characterized for catalytic efficiency, thermal and pH stability, detergent and solvent tolerance. Thermoactive proteases are preferred in many industrial applications as they allow higher processing temperatures to be used. The higher processing temperature provides an acceleration in reaction rates, an increase in the solubility of non-gas reactants, and a decrease in the risk of microbial contamination from mesophilic organisms. 12
To our knowledge, the AprX gene has not been recombinantly produced and characterized before. NCBI BlastP analysis showed that the closest thermostable serine protease gene recombinantly produced belonged to the Bacillus subtilis 168 strain with 56% amino acid sequence identity. In addition, the AprX gene shows 54% amino acid sequence identity 21 with the full protein sequence of the novel antifungal protease identified from Bacillus licheniformis W10 and 68% amino acid sequence identity with the peptidase domain portion. 22 From this point of view, the AprX gene is a candidate to be both a novel protease suitable for industrial use due to its thermostable feature and an antifungal agent that can be used in pesticide treatment due to its similarity in the peptidase domain of an antifungal protease.
Materials and Methods
Recombinant Expression and Purification of AprX Protease
Bacterial strains and plasmids
Geobacillus thermoleovorans ARTRW1, isolated from Armutlu hot springs in Yalova, TURKEY, was used as the source of total DNA for PCR analysis. E. coli BL21 (DE3) was used as host strains for transformation. The vectors used for cloning and expression were pJET1.2/blunt (Thermo Scientific) and pET-28a (+) (Novagen) respectively. Accession number(s): the genome sequences of the Geobacillus Thermoleovorans ARTRW1 has been deposited at DDBJ/EMBL/Genbank under the accession number CP042251. Associated GenBank accession number for AprX protease is QDY71944.1.
Cloning of AprX protease in pET-28a (+)
The synthetic primers AprX R: 5′- agaAAGCTTCTAGCGCTGCAGC -3′ and AprX F: 5′- aggCCATGGgtATGTTCACCTATTCGATC -3′ were used to sequence both serine protease AprX gene strands, using the HindIII and NdeI restriction sites. PCR reactions were carried out using a Gene-Cycler (Bio-Rad). The reaction conditions used were: 1 cycle (98°C for 2 min), 40 cycles (98°C 30 sec, 60°C 30 sec, and 72°C 1.5 min), and a final cycle (72°C for 10 min). The amplified gene was recovered from agarose gel using Macharey-Nagel NucleoSpin Gel and PCR Clean-up kit according to the manufacturer's protocols.
The insert was cloned into the pJET1.2/blunt (Thermo Scientific) cloning vector according to the manufacturer`s protocol. Colony PCR was used to screen the colonies. PCR was carried out using a Gene-Cycler (Bio-Rad). The reaction conditions used were: 1 cycle (95°C for 3 min), 35 cycles (94°C 30 sec, 60°C 30 sec, and 72°C 1 min), and a final cycle of 72°C 5 min. The colony with the insert was subjected to DNA sequencing to confirm the protease gene insertion. The fragment was released by cleavage with restriction enzyme HindIII/NdeI and sub cloned into the intracellular expression vector.
Expression of AprX protease in pET-28a (+)
The vector was transformed into E. coli BL21 (DE3). An 18-h recombinant E. coli BL21 (DE3) inoculation culture was obtained on LB broth containing 55 μg/L kanamycin at 37°C. The inoculation culture was transferred to 1 L of MMI medium (12.5 g tryptone, 25 g yeast extract, 8.5 g NaCl, 0.48 gr Trizma Base (pH 7.4), and 4 mL glycerol in 1 L). The enzyme was overexpressed by the addition of 1.0 mM IPTG (final concentration) and incubated for 24 h at 18°C and 200 rpm. Cells were harvested by centrifugation (10,000 g for 10 min at 4°C) and lysed using BugBusterTM Protein Extraction reagent (Novagen) according to the manufacturer's protocol. 5 mL of BugBusterT reagent and 10 μL of Lysonase Bioprocessing ReagentTM were used per gram of wet cell paste.
Enzyme extracts were analyzed by 12.5% SDS-PAGE using a Bio-Rad Mini-Protean Tetra Cell. Gels were visualized by one step stain using Expedeon InstantBlueTM Coomassie Stain. Western Blot transformation was achieved using a BioRad TransBlot wet transfer system overnight at 100 mA to a nitrocellulose membrane. AprX was detected by incubation with an alkaline phosphatase-conjugated 6x-His-Tag monoclonal antibody (3D5)-AP (Thermo Scientific R932) at 1:5000 dilution on a rocking platform overnight at 4 °C. Detection of the antibody was performed using 1-Step NBT/BCIP SubstrateTM (Thermo Scientific CN:34042) according to the manufacturer`s procedure.
Purification of AprX by Ni-NTA affinity chromatography
Nickel-chelating affinity chromatography (Ni-NTA) purification was performed for purification of recombinant Serine Protease AprX expressed in Pet28 a (+) expression system. The recombinant protein was purified using GE Healthcare HisTrap HPTM column (1 mL) according to the manufacturer's instructions using the AKTA Start Chromatography System. The purity of fractionated samples was evaluated by 12.5% SDS-PAGE.
Verification of AprX expression by mass spectrometry
Cleared supernatant from E. coli expression of AprX was separated on 10% SDS-PAGE. The bands from the SDS gel were cut out, subjected to in-gel trypsin digestion. 23,24 The resulting tryptic peptides were analyzed using a nano LC-MS/MS system (Thermo Scientific™ UltiMate™ 3000 RSLC Nano System Ultra-Performance Liquid Chromatography and Thermo Scientific Q ExactiveTM HF mass spectrometer with a Thermo Scientific™ Nanospray Flex™ ion source). A trap column and an analytical column was used for the separation of tryptic peptides, which are Acclaim PepMap C18, 100 Å, 3 μm 75 mm i.d. × 20 mm Thermo Scientific™ and Acclaim PepMap C18, 100 Å, 3 μm 75 mm i.d. × 150 mm Thermo Scientific™, respectively. The flow rate was 300 nL/min whereas the column temperature was 40°C. A gradient of 10% B to 65% B in 30 min period was used. Buffer A was 2% of acetonitrile, 0.1% of formic acid and Buffer B was 90% of acetonitrile, 0.1% of formic acid.
The MS instrument was run in positive ion mode within the full scan MS range of 375–1,500 m/z. The resolution was 70,000 for the MS mode with the automatic gain control (AGC) of 1x106 ions and a maximum ion transfer (IT) of 20 ms. Source ionization parameters were as follows: capillary temperature 270°C; s-lens level, 80; spray voltage 2 Kv; and MS/MS resolution 17.500 with the AGC value of 1x105 ions. Data analysis was performed using Proteome Discoverer 2.4 (Thermo Scientific) software platform with the Sequest HT search algorithm. The files were searched against the Swissprot Bacteria (reviewed) and Swissprot Geobacillus thermoleovorans Aprx databases with the following parameters: two missed cleavages, precursor ion mass tolerance of 10 ppm, fragment mass tolerance of 0.05 Da and a fixed modification of carbamidomethylation of cysteine. The classification of the proteins into the taxonomy and biological process ontology was performed using the Proteome Discoverer 2.4 software.
Bioinformatics Analysis of the AprX Protein
The physiochemical characteristics of the AprX protein were analyzed with the ExPASy online software ProtParam. 25 Conserved domain analysis of serine protease AprX was performed using NCBI CDD/SPARCLE conserved domain database. 26 SignalP software was used for signal peptide prediction. 27 Functional domains were identified using the UNIPROT server. 28,29
Determination of Protease Cleavage Sites
Bovine serum albumin (BSA) was used as a substrate and was digested in separate vials using trypsin and AprX enzymes. Digestion was carried out by the Filter-aided Sample Preparation (FASP) method using the FASP Protein digestion kit from Abcam, according to the manufacturer`s instructions. The enzyme:protein ratio of 1:25 (w:w) was used for both enzymes. Digestion was carried out for 3 h at 37°C. The resulting peptide mixtures were analyzed using Nano LC-MS/MS as described above.
Determination of Kinetic Parameters
To determine Michaelis-Menten kinetic parameters KM and V max values of recombinantly produced Serine Protease APRX, a universal protease activity assay was used using casein as substrate. 30 The unit was defined as the quantity of enzyme that liberates 1 μg of tyrosine from casein per minute per milliliter. The activities of the casein substrate were measured in the range of 0.0–13.50 mmol/L. The substrate concentrations at which the reactions were performed were 0.11, 0.21, 0.42, 0.84, 1.69, 3.38, 6.75 and 13.5 mM. GraFit 7.0 software was used to calculate kinetic parameters.
Protease Activity Standard Assay
Thermo Scientific Pierce™ Fluorescent Protease Test Kit was used for enzyme activity analysis. The kit contains fluorescently labelled casein as a substrate (FITC). The analysis involves evaluating the protease activity in the sample with a fluorescence resonance energy transfer (FRET) method using a standard. Fluorescent properties of FITC substrate change significantly after digestion by proteases resulting in a measurable indicator of proteolysis. Beckman Coulter DTX 880-Multimode Detector was used for analysis. The enzyme assay was performed according to the manufacturer`s protocol. Briefly, TPCK Trypsin was used as the reference protease calibrator. A calibration curve was created by analyzing 8 calibration points in the range of 0–0.5 μg/mL. The quantity of the AprX enzyme was compared to TPCK Trypsin. All analyses using the standard assay were performed at a concentration of 0.3 μg/μL to ensure a linear response. Blanked Relative Fluorescence Unit (RFU) was used for the calculation of residual proteolytic activity in the assay. Non-homogeneous assay protocol was performed to analyze the pH dependence of the enzyme.
Optimum Conditions for Enzyme Activity
The effect of temperature on protease activity
To determine the optimum temperature of the enzyme, the reaction was set up on ice in the presence of 5 μg/mL FITC-casein at pH 7.8 and transferred to the incubator at temperatures ranging from 23–90°C. The enzyme and substrate were incubated at the respective temperature for 30 min and allowed to cool to room temperature before measurement. Fluorescence was measured in a plate reader. Relative Fluorescent Units (RFU) were used to calculate residual proteolytic activity in the assay.
The effect of pH on protease activity
To determine the optimum pH of the enzyme, the enzyme was incubated at various buffers (sodium phosphate buffer (pH 6.0–8.0), Tris-HCl (pH 8.0–9.0), and glycine-NaOH (pH 9.0–13.0)) with different pH values in the presence of 5 μg/mL FITC-casein at 55°C for 30 min. The reaction mixture was allowed to cool to room temperature (RT) before measurement. Fluorescence was measured in a plate reader. Relative Fluorescent Units (RFU) were used to calculate residual proteolytic activity in the assay.
The effect of NaCl on protease activity
To determine the effect of NaCl on enzyme activity the reactions were set in the presence of 5 μg/mL FITC-casein at pH 7.8 containing 0.2%, 0.38%, 0.78%, 1.56%, 3.13%, 6.25%, 12.5%, 25%, 35% and 40% NaCl. The reaction mixture was incubated at 55°C for 30 min and allowed to cool to room temperature before measurement. The activity amount of the enzyme solution without NaCl was accepted as 100%. The activity of NaCl added samples was calculated in terms of residual activity.
Effects of Various Factors on the Protease Activity
The effect of metal ions on protease stability
To determine the stability of the enzyme in the presence of metal ions, the enzyme activity was measured in presence of the metal salts (CaCl2, CoCl2, MnSO4, ZnSO4, KCl, MgCl2, NiCl2 and CuSO4) at 1 mM and 5 mM final concentration. The reaction mixture was incubated for 8 h. The activity amount of the enzyme solution without metal ions was accepted as 100%. The activity of samples with metal ions was calculated in terms of residual activity.
The effect of protease inhibitors on protease stability
To determine the stability of the enzyme in the presence of inhibitors, iodoacetic acid (1 and 5 mM), PMSF (1 and 5 mM), mercaptoethanol (1 and 5 mM), EDTA (1 and 5 mM), DTT (1 and 5 mM), aprotinin (100 μM) and leupeptin (100 μM) were added to the reaction mixture after which the reaction mixture was incubated for 1 h. The activity amount of the enzyme solution without inhibitor was accepted as 100%. The activity of samples in the presence of protease inhibitors was calculated in terms of residual activity.
The effect of organic solvents on protease stability
To determine the stability of the enzyme in the presence of solvents, 100 nM of the enzyme was prepared in Tris-HCl buffer pH 7.8 and was mixed with some organic solvents (butanol, 2-propanol, acetone, acetonitrile, 1-propanol, ethanol, methanol, and chloroform) at 50% (v/v) final concentration after which the reaction mixture was incubated for 1 h. Relative and residual activities were measured according to standard assay. The activity amount of the enzyme solution without organic solvent was accepted as 100%. The activity of samples in the presence of organic solvents was calculated in terms of residual activity.
The effect of detergents and surfactants on protease stability
To determine the stability of the enzyme in the presence of detergents, the enzyme activity was measured in presence of the laundry detergents [Kosla Vanish, Frosch, Bingo Ultra White, Dalin, Perwoll, Perwoll Colored Clothes, Perwoll Blacked Clothes, Mr. Wipes, Woolite, Omo (Powder), Alo Kar Çiçeği (Powder) and Bingo Matik (Powder)] at 10 mg/mL final concentration and also surfactants [sodium dodecyl sulfate (SDS), Triton X-100, Tween80, Tween20, H2O2] at 1% final concentration. The reaction mixture was incubated for 30 and 60 min. The activity amount of the enzyme solution without detergents and surfactants was accepted as 100%. The activity of detergent and surfactant containing samples was calculated in terms of residual activity.
Results and Discussion
Expression of AprX Protease in pET-28a (+)
Our initial efforts in recombinant expression of AprX protease were to clone the AprX gene into a pET-42b (+) vector to produce an IPTG-inducible fusion protein containing N-terminal GST and His tag. As a result of the study, it was confirmed that the soluble AprX enzyme was produced, but AprX lost its N-terminal 6-His-tag during the expression and lysis process (Supplementary Material, Supplementary Figure S1). Therefore, the serine protease structural gene (AprX) was cloned into the pET-28a (+) vector, in which a C Terminal His-Tagged AprX was rendered IPTG-inducible in an E. coli BL21 strain. The overexpressed AprX protein was predominantly in the insoluble fraction after sonication.
To determine optimum conditions for soluble protein expression, 2 different expression temperatures (18°C and 37°C), 5 different induction concentrations of IPTG (0, 0.25, 0.5, 1.0 and 2.0 mM), 3 different expression time points (4, 6, 24 h of culture post-induction) and 2 different Ni-NTA purification methods (native and urea denaturing) were tested. The best yield of protein in soluble form was obtained at conditions of 1 mM IPTG at 18°C and 24 h of incubation. The recombinant protein was purified using the His-Trap HP column using AKTA Start FPLC. The targeted protein was successfully bound to Ni-NTA resin. Supplementary Figure S2 shows the SDS PAGE analysis of the samples obtained during His Tag purification. The protein recovery after purification was 4.2% and the increase in the specific activity of the purified enzyme to the casein as substrate was calculated as 77.9-fold (Supplementary Table S1). The total purification yield for the AprX expression was approximately 9.8 mg for 1 L of culture.
Verification of AprX Expression by Mass Spectrometry
Gel-based proteomics approach was used to verify the AprX expression. Cleared supernatant from E. coli expression of AprX was separated on 10% SDS-PAGE. Based on SDS-PAGE analysis, the molecular weight of the enzyme was estimated to be approximately as ∼38 kDa (Fig. 1a). The 38 kDa gel band from the SDS-PAGE was cut out, subjected to in-gel trypsin digestion. The resulting tryptic peptides were analyzed by Nano LC-MS/MS analysis. Peptide spectra of the trypsin-digested protein were matched to the AprX protein using the SEQUEST search engine and 29 unique AprX peptides were identified. These spectra matched the predicted 442 amino acid sequences of the AprX sequence with >99% confidence, with a score of 1555 according to the SEQUEST scoring algorithm (Fig. 1b).
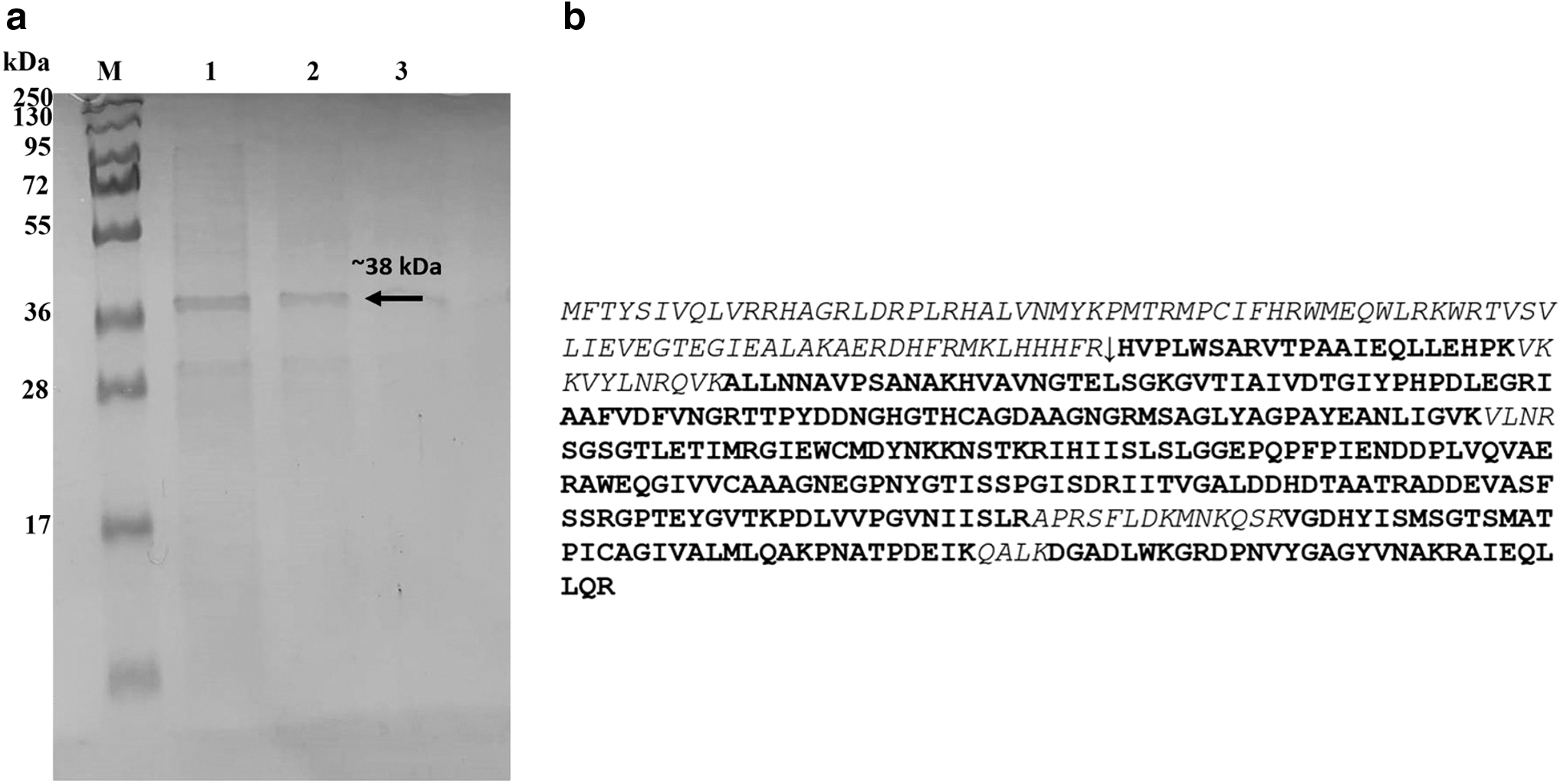
The 38 kDa band was shown to include the whole protease except the N terminal 85 amino acid sequence. The AprX gene from other species was shown to be expressed as pro-enzyme and subjected to N terminal cleavage after biosynthesis. 31,32 Since the 85 aa residue in the N terminal end of the protease is missing, it is highly probable that the enzyme goes through a post translational modification after the synthesis to be in the active form. It was also determined by proteomic analysis that small protein bands seen below the 38 kDa band were fragments of different sizes of the protease itself (data not shown).
Bioinformatics Analysis of AprX Protease
Expasy ProtParam analysis revealed that the mature protein consists of 357 amino acids and has a molecular weight of 38114 Da and a theoretical pI of 6.55. Using the structure-based alignment of more than 40 subtilisin-like protease members, Siezen et al. demonstrated that there is a core of structurally conserved regions in subtilisin-like proteases. 33 This core is approximately 190 residues containing almost all common α-helix and &-strand elements, including catalytic residues Asp 32, His 64 and Ser 221, which are conserved at the active site for most proteases. According to the conserved domain analysis of serine protease AprX performed by NCBI CDD/SPARCLE conserved domain database 26 analysis, the novel AprX produced in this study has similarity with other subtilases starting at residue 142 and extending to the C-terminus of the protein.
The active site residues Asp-151, His-183 and Ser-381 appear to be clearly conserved (Supplementary Figure S3) and include the “structurally conserved regions” defined by Siezen & Leunissen. 27 SignalP 5.0 is a server that predicts the presence and, if any, location of signal peptides in proteins from gram-positive and gram-negative bacteria, archaea and eukarya. 35 Analysis of the primary sequence of the AprX with the SignalP 5.0 server has shown that the likelihood of the AprX protease to include a signal peptide is very low. Therefore, the deduced amino acid sequence after comprehensive research revealed an N-terminal pro-peptide of 85 residues followed by the mature protein comprised of 357 residues.
Characterization of the Autoproteolysis of AprX
It has been observed that the pure protein obtained hydrolyzes itself over time introducing various autoproteolysis products. To examine this autoproteolysis process in detail, the protease was incubated for 6 h at 4°C, 25°C and 45°C in the presence or absence of PMSF, a protease inhibitor. The samples were analyzed with SDS-PAGE (Fig. 2a). It was observed that the band corresponding to 38 kDa nearly disappeared at 45°C. The fact that the same disappearance was not apparent at 4°C and 25°C suggests that the activity of the protease band is increased at high temperatures, which is expected for a protease derived from a thermophilic organism. To observe the kinetics of the change of protease band, samples were incubated at 45°C for 0, 0.5, 1, 2, 3, 4, 5 and 6 h and analyzed by SDS PAGE (Fig. 2b). It was observed that while a protein band corresponding to 25 kDa increased, the band corresponding to 38 kDa nearly disappeared in 6 h at 45°C. As the protease band decreases over time, the formation of small protein fragments has proved that the enzyme cleaves itself with time.

The samples were also analyzed using Western Blot analysis and it was observed that only the protease band at 38 kDa constitutes the C-terminal His Tag (Fig. 2c). Each protein band on the SDS-PAGE gel in Fig. 3C was subjected to proteomic analysis and it was found that the 38 kDa band is the only fragment containing all three active site residues and conserved regions. Therefore, we speculate that the 38 kDa band is the only fragment with the activity.
Determination of Protease Cleavage Sites
To better understand substrate recognition by AprX, we identified the protease cleavage sites using proteomic analysis. Bovine serum albumin (BSA) was used as a substrate and was digested in separate vials using trypsin and AprX enzymes. The resulting peptide mixtures were analyzed using Nano LC-MS/MS. Table 1 summarizes the results of the proteomic analysis results. When the results were evaluated, we saw that both enzymes effectively digested BSA after 3 h of incubation. The sequence coverage obtained for BSA using Trypsin and AprX enzymes was 85% and 71%, respectively. The number of peptides obtained after digestion was found to be 79 for trypsin and 382 for AprX.
Trypsin has a very specific function; it only cleaves peptide chains on the carboxyl side of the amino acids lysine (K) or arginine (R). Therefore, the number of peptides is lower compared to a non-selective enzyme. Examination of the peptides obtained by trypsin cleavage has shown that these peptides selectively contained C-terminal R/K residues. Due to its high selectivity, trypsin is exclusively used in mass spectrometry facilities for protein identification purposes. The high peptide count obtained after cleavage with the AprX enzyme is simply an indication that AprX is not very specific.
Investigation of the peptides obtained by AprX cleavage in detail has shown that the majority of the peptides ended with C-terminal R/K and L residues indicating that the AprX protease has trypsin-like activity. However, several peptides ending with other amino acids E, S, V, F, G, H, N and T have also been observed confirming that it cleaves the substrate randomly. This showed that the enzyme non-specifically cleaved its substrate.
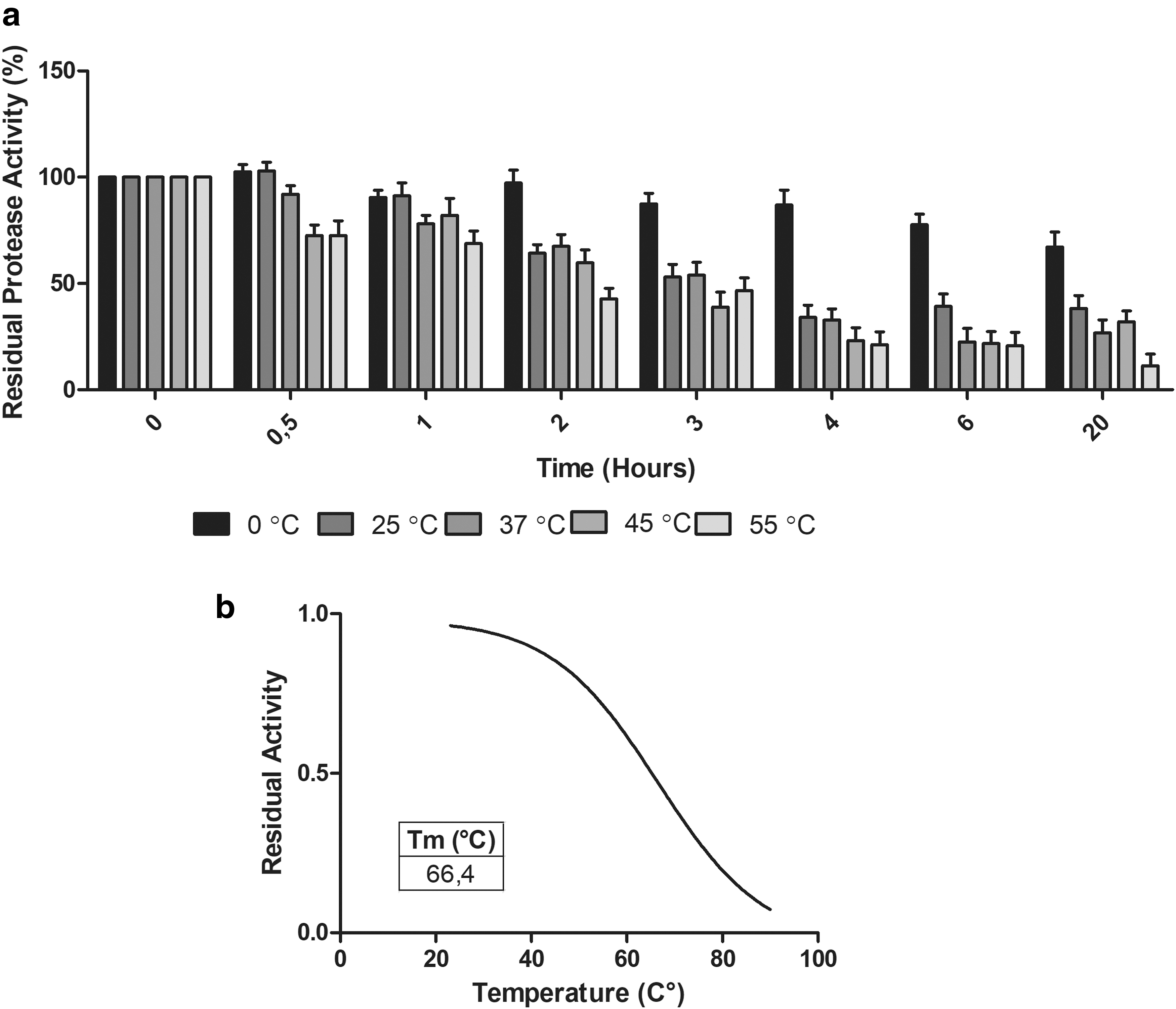
Thermal Stability of AprX
The thermal stability of AprX was examined by the incubation of the enzyme at pH 7.8 for different time intervals (0, 0.5, 1, 2, 3, 4, 5, 6 and 20 h) at temperatures of 0°C, 25°C, 37°C, 45°C and 55°C (Fig. 3a). The activity of the enzyme was retained at more than 65% after incubation at 55°C for 1 h. The enzyme also retained 45% of its activity following incubation at 55°C for 3 h. To calculate the melting temperature (Tm) of the AprX, the enzyme was incubated at various temperatures between 20°C to 90°C for 30 min and the activity was measured. The residual activity was plotted against temperature using a non-linear regression curve (Fig. 3b). The Tm of the enzyme was determined as the temperature at which the 50% residual activity was retained. Tm of the enzyme was determined as 66°C.
Determination of Kinetic Parameters
The kinetic parameters Km and Vmax values of recombinantly produced serine protease AprX were determined from the Michaelis-Menten plot. Casein was used as a substrate at the range of 0.0–13.5 mM. Kinetic parameters of the purified protease for hydrolysis of casein are presented in Table 2. The results indicated that the enzyme has a significant activity towards casein with a Km value of 0.78 mg/mL. The deduced catalytical efficiency (kcat/Km) of the AprX enzyme was 4.35 (min−1 mM−1).
Optimum Conditions for Enzyme Activity
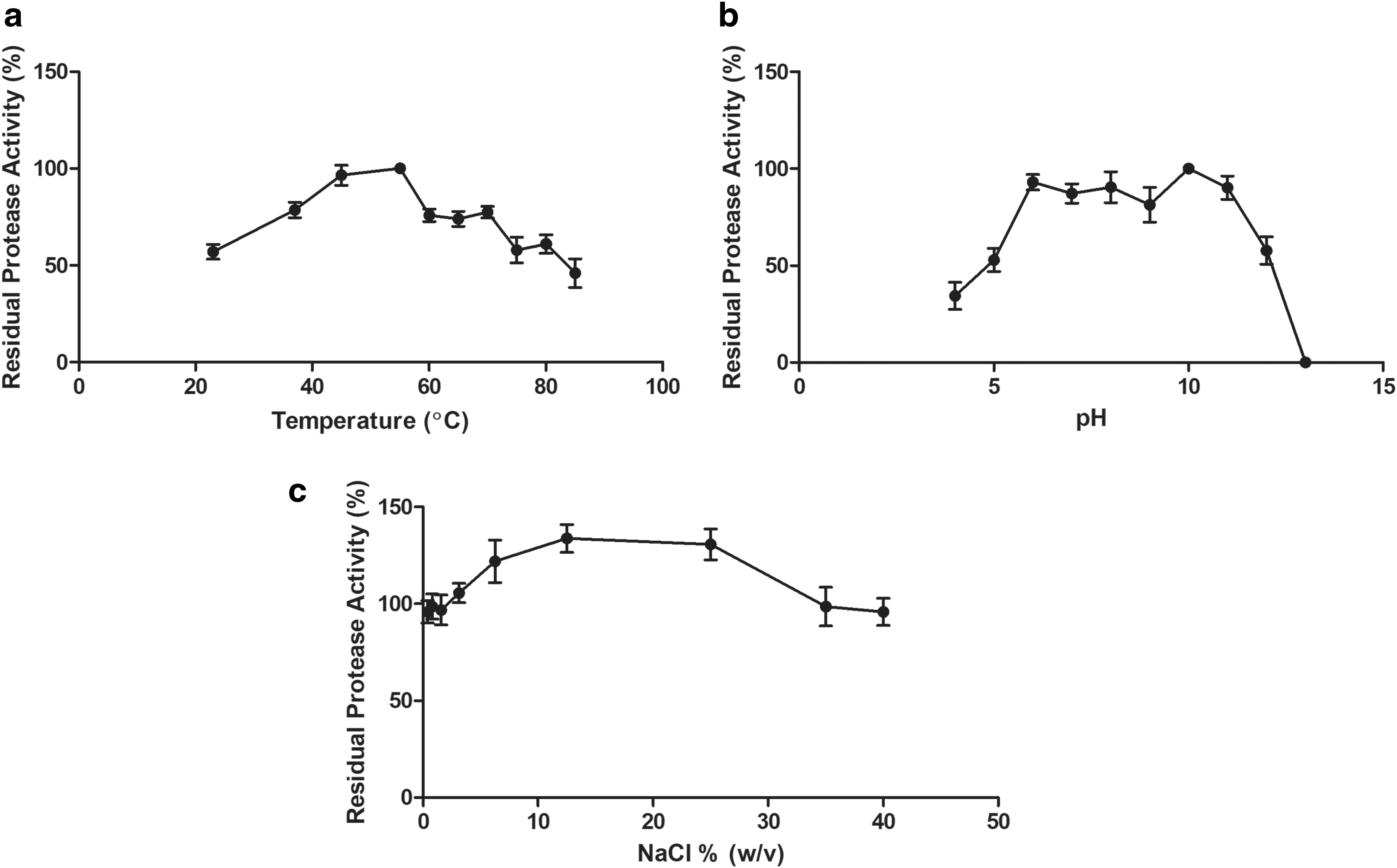
Concerning proteolytic activity, the optimum temperature was determined by conducting protease assay at pH 7.6 at different temperatures from 23°C to 90°C. The optimum temperature for the activity was determined to be 55°C (Fig. 4a). It was observed that the enzyme was active at a broad temperature range from 25°C to 85°C. The enzyme retains approximately 80% of its activity in the range of 50°C to 70°C. To determine the optimum pH of the enzyme, the protease activity was measured over a pH range of 5.0 to 13.0. The residual proteolytic activity was determined by standard assay. The protease was highly active in the pH range of 5.0 to 12.0 with an optimum pH of 6.0 to 11.0 (Fig. 4b).
Effect of
Considering that the pH range of laundry detergents is generally 9.0 to 12.0, this property is very important for the possible use of AprX in detergent formulations. In addition, the enzyme retains more than 85% of its activity at the range of 6.0 to 11.0, suggesting that the AprX enzyme is an alkaline serine protease. It has been known that salt has a protective role in stabilizing most of the proteases. 34 –36 Therefore, the effect of NaCl on the activity of protease was also evaluated by adding NaCl into the reaction mixture at a final concentration of 0.2 to 40% (Fig. 4c). Figure 5C shows that 12.5% of NaCl increases the activity of the enzyme significantly.
Effect of commercial detergents on the activity of AprX when FITC-labelled casein was used as a substrate.
Effects of Various Factors on the Protease Stability
The effect of metal ions on protease stability
To determine the effect of metal ions on the activity of the enzyme, the activity was measured in presence of the metal salts [CaCl2, CoCl2, MnSO4, ZnSO4, KCl, MgCl2, NiCl2 and CuSO4] at 1 mM and 5 mM final concentration. The results are given in Table 3. It was seen that K+ ion at 5 mM concentration increased the activity of the enzyme to 220%. The activity was decreased to 42% and 32% in the presence of Mn2+ and Cu+ ions in 5 mM concentration, respectively. Ni2+ and Zn2+ had a great inhibitory effect on the enzyme, 1 mM of these ions completely inhibited the activity. It has been reported by several authors that trypsin-like serine proteases are deactivated in the presence of Ni2+ and Zn2+ ions. 37 Zn2+ mediated inhibition of trypsin-like proteases has been structurally investigated and reported. 38 The finding of inactivation in the presence of Zn2+, combined with the fact that the protease has a partial preference to cleavage after the R and K residues indicates that AprX is a trypsin-like serine protease.
Not significant: n.s.; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.000.
The effect of organic solvents on protease stability
The activity of the enzyme, in the presence of solvents butanol, 2-propanol, acetone, acetonitrile, 1-propanol, methanol, ethanol, and chloroform, were also tested. The effects of organic solvents at 50% v/v concentrations are summarized in Table 3. It was determined that the activity was lower than 50% of the maximum activity in the presence of chloroform, acetone, acetonitrile, propanol, and isopropanol. The presence of butanol reduced the activity to 54%. The enzyme retained above 80% of its activity in the presence of ethanol. It can be concluded that apolar solvents strongly reduce the activity of the enzyme while more polar ones have a smaller effect.
The effect of inhibitors on protease stability
To determine the stability of the enzyme in the presence of inhibitors, iodoacetic acid (1 mM and 5 mM), PMSF (1 mM and 5 mM), β-mercaptoethanol (1 mM and 5 mM), EDTA (1 mM and 5 mM), DTT (1 mM and 5 mM), aprotinin (100 μM) and leupeptin (100 μM) were added to the reaction mixture after which the reaction mixture was incubated for 1 h. The activity amount of the enzyme solution without inhibitor was accepted as 100%. The activity of the samples containing protease inhibitors was calculated as a residual activity.
Table 3 summarizes the effect of inhibitors on the AprX activity. It was observed that the protease lost its activity in the presence of 1 mM and 5 mM PMSF and 100 μM Aprotinin. PMSF and Aproptin are serine protease inhibitors. Since AprX belongs to the serine protease family, it is an expected finding that AprX loses its activity in the presence of these inhibitors. On the other hand, it was observed that the enzyme retained its activity in the presence of iodoacetic acid and DTT. The activity was increased in the presence of β-mercaptoethanol suggesting that the enzyme is thiol dependent. 7 EDTA, a metalloenzyme inhibitor, inhibited the enzyme activity by 56% in 1 mM concentration suggesting that the AprX is metalloserine protease.
The effect of surfactants on protease stability
To determine compatibility with surfactants, the enzyme activity was measured after pre-incubation in the presence of the surfactants sodium dodecyl sulfate (SDS), Tween80, Triton X-100 and H2O2 at 1% (w/v) final concentration. Enzyme retained more than 90% of its activity in the presence of SDS and Tween-80 (Table 3). The activity of an AprX was completely inhibited by Triton X-100, which is the case for some of the serine proteases reported. The activity of the enzyme was decreased to 54% in the presence of H2O2. Literature search on serine proteases has shown that SDS stable proteases are very rare. Therefore, high stability in the presence of SDS is important.
The effect of detergents on the activity of the protease
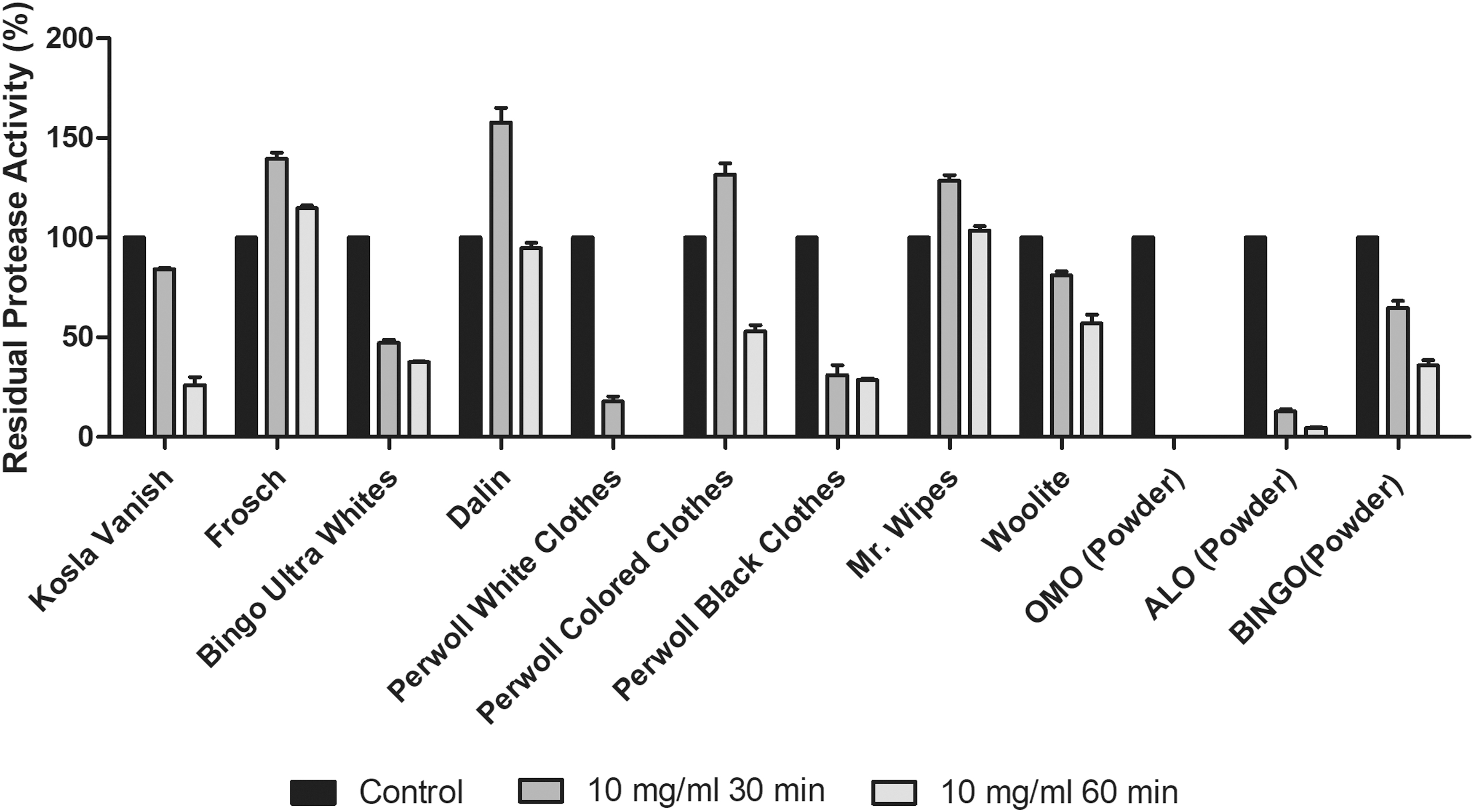
The detergent industry is one of the major application areas of proteases. Therefore, it is crucial to study the enzyme's activity in the presence of laundry detergents and surfactants. To determine the compatibility with detergents, the enzyme activity was measured after pre-incubation in the presence of the laundry detergents [Kosla Vanish, Frosch, Bingo Ultra Whites, Dalin, Perwoll White Clothes, Perwoll Colored Clothes, Perwoll Blacked Clothes, Mr. Wipes, Woolite, Omo (Powder), Alo (Kar Çiçeği-Powder) and Bingo (Powder)] at 10 mg/ml for 30 min and 60 min. The effect of detergents was evaluated and results are given in Fig. 5.
The activity of the enzyme was decreased by 50% in the presence of powder detergents such as OMO, ALO, BINGO and the ones with bleaching activity such as KOSLA vanish and Perwoll white-colored clothes in 30 min. This finding suggests that AprX's activity is reduced in the presence of relatively strong detergents, possibly due to the whitening chemicals in the formula. It has been observed that AprX maintains its activity for 60 min and its activity increases in the first 30 min in the presence of detergents Frosch, Dalin, Perwoll Colored Clothes, Mr. and Wipes. Some of these detergents are organic and chemical-free, and some of them are specific to delicate laundry. This raises the possibility that a similar protease has been added to these detergents, and may even be formulated to increase enzyme activity.
Conclusion
Our study confirmed that the AprX gene from Geobacillus thermoleovorans ARTRW1 produced a serine protease with a molecular weight of 38.1 kDa and a theoretical pI of 6.55. The enzyme was shown to be highly active at high temperatures and high pH values. The optimum activity was observed at 55°C and pH from 6.0 to 11.0. The enzyme retained its activity over a broad temperature range of 25°C to 85°C consistent with the thermophilic nature of the protein. The enzyme showed a maximum activity towards casein with a kcat and Kcat/Km values of 3.39 min−1 and 4.35 min−1.mM−1, maximum activity under optimum conditions. It was found stable from 25°C to 55°C and retained about 50% of activity for 3 h at 55°C.
Thermostable proteases, whose thermostability studies have been reported in the literature, have been shown to have a half-life of 1 h at 50°C, 39 25 min at 60°C. The half-life of subtilisin Carlsberg, was reported as 2.5 min at 60°C. 21 In contrast, our results show higher thermal stability at 55°C. Since AprX Geobacillus thermoleovorans ARTRW1 is a highly thermostable alkaline serine protease which cleaves the substrates non-specifically, it can be a candidate to be used in industrial processes requiring long term stability at elevated temperatures. The enzyme almost retained its activity in the presence of methanol and ethanol and retained 50% of its activity in the presence of butanol. Other solvents had an inhibitory effect on the enzyme. The effects of various metal ions, detergents and inhibitors were also investigated. The protease is highly tolerant to detergents compared to similar enzymes in the literature.
The detergent tolerance of the enzyme also makes it a suitable candidate to be used in detergent formulations since in industrial laundry conditions alkaline proteases are commonly used for 10 min at 60°C. As a result, a highly active, detergent tolerant and thermostable protease was produced, purified and characterized. This protease obtained can be used in many industrial applications. Further study should be carried out on the characterization of the enzyme including the characterization of the auto cleavage process of the AprX and prevention of the self-cleavage of protease using site-directed mutagenesis approaches. Prevention of the self-cleavage of the protease would eventually increase the enzyme stability.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.