
Abstract
Cellulomonas flavigena, a Gram-positive and cellulolytic bacterium, is known to produce a curdlan-type exopolysaccharide (i.e., linear β-1,3-glucan). However, curdlan-producing colonies of this organism do not stain on aniline blue agar, a defining characteristic of other curdlan-producing bacteria. In this study, Cellulomonas curdlan, extracted with the inert solvent DMSO, caused a redshift in the absorption spectrum of aniline blue, matching results reported for Agrobacterium curdlan and confirming earlier biochemical analyses. Cellulomonas curdlan has many biotechnological applications.
Introduction
The bacterial polysaccharide known as curdlan was first discovered as a product of the Gram-negative strain Alcaligenes faecalis var. myxogenes 103C, which has since been reclassified as a member of the genus Agrobacterium. 1,2 Agrobacterium curdlan is defined as a linear β-1,3-glucan with a high degree of polymerization (DP ∼500-3,000 glucose residues per polysaccharide chain), is insoluble in aqueous suspension near neutral pH, and possesses characteristic thermo-gelling properties (Table 1). Agrobacterium curdlan, as well as curdlans from other bacterial species, have a wide variety of industrial, medical, and environmental applications including uses as gelling agents in foods, as stimulators of immune responses, and as sequestering agents in bioremediation methods. 2 -8
Physicochemical Properties of Curdlan-Type Polysaccharides
The Gram-positive, bacterial strain Cellulomonas flavigena KU (ATCC 53703) has also been shown to produce high yields of a curdlan-type exopolysaccharide (EPS) when grown in minimal media with a high carbon-to-nitrogen source ratio (C/N). When the chemical structure of this EPS was investigated using standard biochemical techniques, it was found to be a linear, unbranched β-1,3-glucan with an average DP of 500 (Table 1). 9 Thus, the EPS from C. flavigena KU is very similar to Agrobacterium curdlan in terms of primary structure (i.e., monosaccharide composition and glycosidic linkages), although there may be a difference in the DP. Thus, the Cellulomonas EPS meets the criteria for a curdlan-type polysaccharide. Furthermore, Cellulomonas curdlan has the same solubility and thermo-gelling properties as Agrobacterium curdlan (Table 1). And in fact, purified hydrogels of Cellulomonas curdlan, extracted with 1 N NaOH and precipitated with 1 N HCl, stain with the triphenylmethane dye aniline blue, a property specific to curdlan-type polysaccharides (Table 2).
Aniline Blue Staining Reactions of Purified Glucans from Different Sources
However, unlike other curdlan-producing bacteria, colonies of C. flavigena KU do not stain blue on aniline blue agar despite the fact that curdlan-type polysaccharide can be extracted from such colonies. 10,11 This observation raised the intriguing question of whether the “native” form of Cellulomonas curdlan, as it exists in the glycocalyx layer surrounding C. flavigena KU cells, differs from Agrobacterium curdlan and possibly from that of other curdlans from other bacterial species.
Curdlan is soluble in the chemically inert solvent dimethyl sulfoxide (DMSO) (Table 1), and the polymer can be precipitated from DMSO solution as a hydrogel by dilution with an excess of water. The resulting hydrogel can then be washed several times with water to obtain a purified curdlan preparation. This method avoids the potentially damaging effects of treatment with strong alkali and strong acid (e.g., alkaline degradation, acid hydrolysis, etc.) and presumably maintains the native chemical structure of the polysaccharide. DMSO-extracted curdlan from C. flavigena KU cultures was previously shown to contain no detectable non-carbohydrate substituents (i.e., acyl-linked groups), and such curdlan hydrogel preparations exhibit a positive aniline blue staining reaction upon visual inspection. 9,11
The aim of this brief study was to further analyze the interaction between aniline blue and Cellulomonas curdlan. Shortly after its initial discovery, curdlan from strain Alcaligenes faecalis var. myxogenes 103CK (i.e., Agrobacterium curdlan) was shown by Nakanishi et al. to cause a redshift in the visible-range, absorption spectrum of aniline blue as well as a small increase in absorbance. 12 Our preliminary experiments also alluded to a redshift in the absorption spectrum of aniline blue when combined with suspensions of Cellulomonas curdlan. 9,11 Herein we show the effects of DMSO-extracted, Cellulomonas curdlan on the absorption spectrum of aniline blue using the same protocol described by Nakanishi et al. 12
Materials and Methods
Curdlan was prepared from C. flavigena KU cultures following the method of Kenyon and Buller (2002) with minor modifications. 9 Cultures were grown aerobically in M9-yeast extract media with a high C/N (e.g., 1% glucose and 5 mM NH4Cl) at 30°C for 3 days. Culture biomass, consisting of large aggregates of curdlan-producing cells, was collected by centrifugation and washed in physiological saline (0.9% NaCl) following the method of Herbert et al. 13 The high-carbohydrate biomass was then dried by lyophilization, ground to a powder, and stored in a desiccator at room temperature (RT). To extract curdlan, dry biomass was resuspended in concentrated DMSO and stirred for 30 min with a mechanical mixer at room RT. The viscous suspension was centrifuged (25,300 X g for 30 min at RT), and the supernatant was filtered through a glass microfiber filter. The clarified extract was then mixed with three volumes of ddH2O to precipitate curdlan in the form of a hydrogel. Curdlan hydrogel was washed four times by resuspension in water followed by centrifugation under the same conditions. The washed hydrogel pellet was lyophilized, ground to a fine powder, and stored in a desiccator.
This type of polysaccharide preparation was then analyzed for total carbohydrate and protein content and typically found to be >90% carbohydrate by dry weight as determined by the method of Dubois et al. and <0.01% protein by dry weight as determined by the method of Markwell et al. 14,15 Cellulomonas curdlan preparations extracted with NaOH and precipitated with HCl routinely had a higher protein content (∼ 0.2% dry wt.), presumably due to contamination with cellular protein, and did not give reproducible results (data not shown). Therefore, only DMSO-extracted preparations were used in this study.
The method described by Nakanishi et al. 12 was then used to analyze the effects of Cellulomonas curdlan on the absorption spectrum of aniline blue and to directly compare our results with those reported previously for Agrobacterium curdlan (i.e., curdlan from Alcaligenes faecalis var. myxogenes IFO 13140). Differing quantities of dry Cellulomonas curdlan (0 μg, 50 μg, 100 μg, 150 μg, and 200 μg) were suspended in 10 mL of 5 mM NaOH. Each polysaccharide suspension was then added to 10 mL of 0.025% aniline blue in 0.5 M phosphate buffer. After thorough mixing, polysaccharide suspensions were allowed to stand at RT for 2 h. After resuspension, 1 mL of each mixture was then transferred to a quartz cuvette with a path length of 1 cm and absorbance values were read at RT from 520-680 nm using a Beckman DU-40 spectrophotometer. Absorption spectra were then generated by plotting absorbance values vs. wavelengths using SigmaPlot® v15. All chemicals used were from Sigma-Aldrich®.
Results and Discussion
Figure 1 shows that DMSO-extracted, Cellulomonas curdlan caused a redshift in the absorption spectrum of aniline blue, and this effect was proportional to the amount of Cellulomonas curdlan added to the mixture. In Fig. 1, data points are shown at every 40 nm interval between 520–680 nm, and the absorption maxima (λmax) of the aniline blue-curdlan mixtures are marked by vertical dotted lines. A λmax redshift of approximately 12 nm was observed for mixtures containing 200 μg of Cellulomonas curdlan compared to aniline blue alone (i.e., a redshift in λmax from approximately 580-592 nm). By contrast, amylose did not cause a redshift in the absorption spectrum of aniline blue (data not shown). Likewise, Nakanishi et al. reported that the glucans dextran and cellulose also do not significantly affect the absorption spectrum of aniline blue. 12 These observations demonstrate the binding specificity of aniline blue for linear β-1,3-glucans (i.e., curdlan-type polysaccharides).
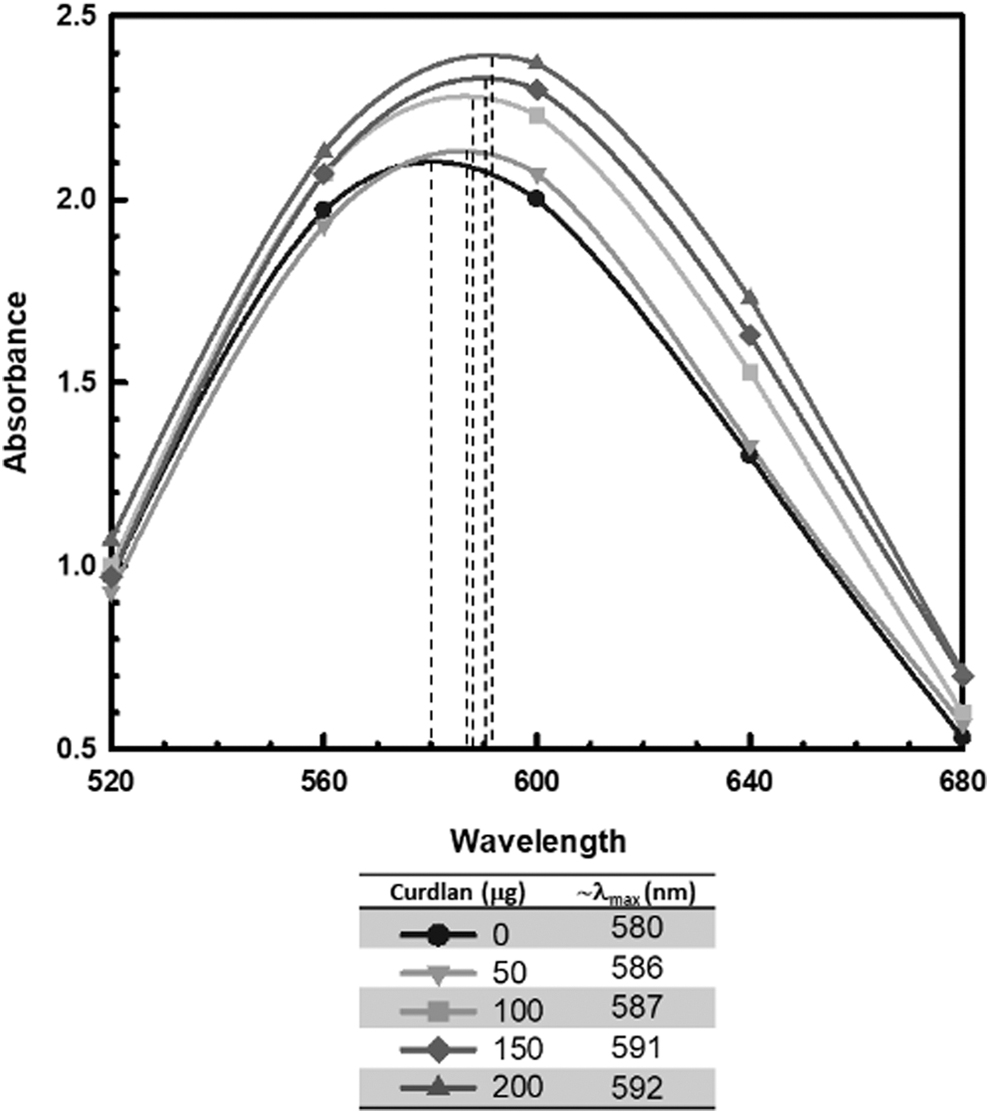
Absorption spectra of aniline blue mixed with increasing amounts of Cellulomonas curdlan. Dashed lines indicate the approximate absorption maxima for each aniline blue-polysaccharide mixture, as summarized in the key. Results are from a representative set of experiments. Cellulomonas curdlan was prepared using the DMSO extraction and purification protocol initially described by Kenyon and Buller (2002) 9 as modified herein.
These results strengthen the conclusions of earlier biochemical analyses, and help to confirm that the EPS produced by C. flavigena KU (ATTC 53703) is indeed a curdlan-type polysaccharide. 9,11 In light of these results with DMSO-extracted curdlan, it is likely that the lack of staining of C. flavigena KU colonies on aniline blue agar is due to the nature of the Cellulomonas glycocalyx, as previously theorized. 9,11 In support of this hypothesis, “native” curdlan (i.e., in situ curdlan as part of the glycocalyx, capsular layer surrounding bacterial cells) is highly resistant to both acid and enzymatic hydrolysis, perhaps suggesting that it has a higher, intrinsic degree of crystallinity compared to “regenerated” curdlan (i.e., solubilized and reprecipitated curdlan). 11,16
Although the DMSO-extracted curdlan used here should be chemically unaltered, it is likely that the physical conformations of polysaccharide chains undergo significant changes due to solubilization, and this physical transition may explain the difference in staining properties observed between the native and regenerated forms of Cellulomonas curdlan. In fact, different conformational forms of Agrobacterium curdlan have been known for some time (Table 1), and the degree of crystallinity of β-1,3-glucans is known to affect their staining properties. For example, paramylon granules from Euglena spp. are highly crystalline and recalcitrant to staining with aniline blue. 17 Interestingly, both scanning and transmission electron micrographs of the C. flavigena KU glycocalyx previously suggested a compact curdlan capsule as well as an outer layer of glycocalyx-associated, proteinaceous material, and the biochemical nature of these surface layers awaits analysis. 9,11,18
Conclusion
In conclusion, Cellulomonas curdlan, extracted with DMSO and precipitated with water, interacts with aniline blue to alter the absorption spectrum of the dye and shift the λmax to longer wavelengths (i.e., redshift). This interaction with aniline blue is nearly identical to that previously reported for Agrobacterium curdlan. 12 Therefore, the recalcitrance of living C. flavigena KU colonies to staining on aniline blue agar appears not to be due to the chemical structure of Cellulomonas curdlan, but instead, may reflect a difference in the conformational structure of the polymer and may also involve interference from glycocalyx proteins as previously suggested. 9,11
Interestingly, the physical nature of the curdlan glycocalyx of C. flavigena KU is likely to play an important role in the biofilm-mediated degradation of cellulose by this organism. 19,20 Subsequently, the fact that not all curdlan-producing bacteria stain blue on media containing aniline blue, is an important consideration in future screening and isolation efforts for such organisms. And of special note, curdlan-producing Cellulomonas spp. are an important source of this versatile polymer, offering many advantages over other curdlan-producing bacteria including the absence of endotoxin and production of curdlan from cellulose and hemicelluloses. 9,11,19 -21
Footnotes
Acknowledgments
The authors would like to thank Dr. Sara J. Molesworth-Kenyon, biology professor at the University of West Georgia, for helpful suggestions and critical review of the manuscript. The bacterial strain used in this study (Cellulomonas flavigena KU) was a gift from Dr. Clarence S. Buller, former professor and chair of the Department of Microbiology at The University of Kansas.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by University of West Georgia professional development funds awarded to WJK.