
Abstract
Microalgae metabolites exhibit significant biological activity and can act as antibacterial agents (peptide and lipid fraction) against Gram-positive bacteria Bacillus and as a component of nutrient media in the cultivation of yeast Saccharomyces cerevisiae. In this study, a mixture of metabolites (triacylglycerides, fatty acids, o-dialkylmonoglycerides and glycerol trialkyl esters) extracted from disrupted Chlorella sorokiniana microalgae cells by non-polar solvent had inhibitory effect on the growth of Gram-positive bacteria at photosynthetically active radiation (PAR) levels of 100 ± 6 μmol photons/(m2·s). The minimum inhibitory concentration (MIC) of the extract was ≈80 μg/mL. Out of the isolated components of the mixture, triacylglycerides and fatty acids showed antibacterial properties, with MIC = 176 μg/mL and MIC = 445 μg/mL respectively. The water-soluble peptide fraction extracted from microalgae cells had an inhibitory effect on the growth of Gram-positive bacteria Bacillus both under white light illumination and in the dark; MIC of the peptide fraction is ≈125 μg/mL (when illuminated) and 170 μg/mL (in the dark). The water-soluble protein fraction had no antibiotic properties, but is of interest as a component of nutrient medium (0.01 mL/mL) for the cultivation of yeast Saccharomyces cerevisiae. The addition of this fraction allowed a 21% increase in population capacity compared with the control sample, a 1.4-fold increase in the specific growth rate in the exponential phase and a 28% decrease in cell generation time. The study has contributed to establishing the effect of light radiation on antibacterial properties of microalgae metabolites and identifying the minimal inhibitory concentration with regard to Gram-positive bacteria. Also, the potential of the Chlorella aqueous extract to serve as a component of nutrient media for the Saccharomyces cerevisiae cultivation has been researched.
Introduction
Microalgae are microscopic single-celled organisms that can be used to produce a wide range of metabolites such as proteins, fats, carbohydrates, vitamins, and nucleic acids. These substances are used in various human applications and exhibit numerous biological properties such as antibacterial, antioxidant, antihypertensive, immunomodulatory, and anti-inflammatory. 1 -6
Some of the most remarkable metabolites of microalgae are substances that exhibit antibacterial properties—protein hydrolysates and lipid-type substances. 7 -23 Peptide fractions of microalgae Chlorella sorokiniana, Saccharina longicruris, Nannochloropsis sp. had antibacterial activity against Gram-positive bacteria Staphylococcus aureus and Gram-negative bacteria Escherichia coli. 10 -12 The maximum antibacterial effect against these bacteria was elicited with a peptide fraction with a mass <10 kDa obtained by hydrolysis of proteins with pepsin. 10 However, peptides obtained by protein hydrolysis using trypsin and having a molecular weight greater than 10 kDa had greater antibacterial effect on Staphylococcus aureus. 11 In another experiment, the peptide fraction (<3 kDa) obtained by enzymatic hydrolysis of proteins by papain-pancreatin protease complex showed the strongest antibacterial properties against these bacteria. 12 At the same time, the mechanism of action of antibacterial protein hydrolysates of microalgae on bacterial cells has not been fully studied and requires further research. 13 -15
The mechanism of action of cationic peptides is believed to be based on the destruction of the phospholipid bilayer after specific incorporation into the bacterial cell membrane. For other types of peptides, the mechanism of action is different, and it is assumed that they affect intracellular targets. 15
Studies suggest the prospective use of protein hydrolysates of microalgae of different species as antibacterial agents. Yet, the lack of data on the value of the minimum inhibitory concentration of hydrolysate against Gram-positive and Gram-negative bacteria, which is necessary for comparison with already used antibacterial agents and for determining the possible application of the obtained hydrolysate, were the drawbacks of most of the studies.
Lipids of phototrophic microorganisms also exhibit antibacterial properties. 16 -23 R. Pratt was among the first to investigate the antibacterial properties of lipid metabolites of microalgae. 17 According to that research, Chlorella vulgaris extracts inhibited the growth of Gram-positive and Gram-negative microorganisms such as Staphylococcus aureus, Streptococcus pyogenes, Bacillus subtilis, Escherichia coli and Pseudomonas pyocyanea (Ps. aeruginosa). A group of substances consisting of a mixture of fatty acids with antibiotic action was isolated and named “chlorellin”. 17 Modern studies of lipid extracts from microalgae 18 -23 showed that extracts from Phaeodactylum tricornutum, 18 Scenedesmus sp., 21 Chlorella sp. 22,23 had antibacterial effect on both Gram-positive and Gram-negative bacteria.
Some studies determined the quantitative and qualitative composition of the extracts, which contained polyunsaturated fatty acids (omega-3) – eicosapentaenoic acid and hexadecatrienoic acid – that showed antibacterial properties even in micromolar concentrations, 18 and palmitic acid methyl esters (C16: 0), which were most active against Gram-negative bacteria (Escherichia coli and Pseudomonas aeruginosa) and had a minimum inhibitory concentration of about 12-24 μg/mL. 21
Lipid and pigment extract from Chlorella vulgaris, with a minimum inhibitory concentration against Gram-positive bacteria of 3.0 μg/disc and for Gram-negative bacteria, 2.0 μg/disc, was composed of: neutral lipids 37.1 ± 0.5 %, triacylglycerides 13.4 ± 0.4 %, fatty acids 24.3 ± 0.2 %, polar lipids 0.9 ± 0.1 %, unsaponifiable substances 14.1 ± 0.2 %, chlorophyllides 16.3 ± 0.1 %. 22 When wastewater bacteria were exposed to light irradiation, the antibacterial properties of Chlorella lipid and pigment extract containing triacylglycerides, O-dialkyl monoglycerides, fatty acids, O-dialkyl esters of glycerol and long-chain alcohols were intensified. 23 Bach-Engler peroxidation theory and the branched chain reactions theory by N.N. Semenov may explain the supposed mechanism of enhanced antibacterial action of lipid exometabolites at light radiation. 23
The presented studies conclude that purified lipid extract of microalgae can become the basis of an alternative preparation for preventing microbial contamination of feed and be used in bioremediation. It seems promising to test the hypothesis that light radiation can enhance the antibacterial effect of lipid fractions. It can be assumed that under the action of light radiation these metabolites of phototrophic microorganisms (potentially present in the intercellular space) can acquire properties that allow them to inhibit bacterial cells around them, while phototrophs themselves in the process of evolution have acquired protection against these compounds, for example, a powerful antioxidant system.
Aqueous extracts of Chlorella microalgae (CVE or CGF) containing proteins, polysaccharides, phenolic compounds associated with carbohydrates in the form of glycosides (antioxidants), and nucleic acids protect cells from negative environmental effects and assist in cell regeneration and growth. 24 -31
Chlorella aqueous extract had a protective effect on yeast cells grown on Saburo medium under anaerobic conditions: the addition of 0.1% (vol.) Chlorella aqueous extract to the suspension significantly increased the viability of yeast cells - ≈95% of live cells for 17 days, while in the control sample (yeast suspension without the addition of the extract) it was ≈29%. 25
Ana-1 macrophage, cultured on a nutrient medium with the addition of the aqueous extract of Chlorella pyrenoidosa (200 μg/mL), proliferated 162.98 % more efficiently compared to the control sample. 26 The aqueous extract of Chlorella vulgaris had a significant effect on signalling pathways and proliferation of rat intestinal cells (IEC-6) through the induction of several genes; 27 -30 it also successfully stimulated the growth of Chinese hamster ovary (CHO) and mesenchymal stem cells (MSC) in both 2D and 3D cell cultures under reduced fetal bovine serum (FBS) conditions for 21 days. Thus, Chlorella aqueous extract could be a viable nutrient media supplement with a wide range of applications in biotechnology and tissue engineering.
Having reviewed the current state of research on the biological activity of extracts of phototrophic microorganisms, it can be concluded that there are almost no studies evaluating the potential of aqueous extracts of phototrophic organisms as a component of nutrient medium for cultivation of the main classical biological agents used in industrial biotechnology, namely eukaryotic organisms like yeasts and mold fungi.
In this connection, the aim of the study was to isolate metabolites (protein hydrolysates and lipid-pigment complex) from the cells of microalgae Chlorella sorokiniana and to investigate their antibacterial properties under light radiation and in the dark, to determine the minimum inhibitory concentration against Gram-positive bacteria. Also, the research aimed to investigate the potential of aqueous protein extract to be used as a component of nutrient medium for cultivating yeast Saccharomyces cerevisiae.
Materials and Methods
PROTOCOLS FOR PREPARATION OF EXTRACTS MICROALGAE
The object of the study was strain Chlorella vulgaris Beijer IPPAS C-1 (Chlorella sorokiniana) obtained at the Timiryazev Institute of Plant Physiology of the Russian Academy of Sciences. The microalgae strain was cultured in a 5-L photobioreactor at 30 ± 0.02°C, photosynthetically active radiation (PAR) = 100 ± 6 μmol photons/m2·s, aeration with 0.15-0.02 carbon dioxide gas mixture (flow rate (1 ± 0.2) L/min) using Tamiya nutrient medium. 32 Extracts were obtained from the biomass at the end of its exponential growth phase (7–8 days of cultivation).
PREPARATION OF A WATER-SOLUBLE (POLAR) PROTEIN CONCENTRATE
Separation of fugate from microalgae biomass was carried out using a centrifuge (Sigma® 2-16 RK/2-16P; SIGMA Laborzentrifugen, Osterode, Germany) at 4,000 rpm for 10 min. The obtained biomass (moisture content of 98–99%) was divided into 25 mL samples. Cell disruption of concentrated biomass (with 98–99% moisture content) was carried out using Scientz IID ultrasonic disruptor at 150 W power for 5 min. 33 A preweighed tube with concentrated microalgae cell biomass was placed in a desiccator at 40°C and dried to a constant mass (Δ = 0.01 g) and then weighed. The extraction of proteins from microalgae biomass was carried out for 20 h at 4°C using 25 mL of phosphate buffer (Na2HPO4, Lenreactive, Saint Petersburg, Russia), KH2PO4, Lenreactive) (pH 7.4) as solvent.
After the extraction, the cell biomass was separated from the extract using a centrifuge (IKA-Werke; IKA mini G centrifuge, Staufen im Breisgau, Germany) for 10 min at 6,000 rpm. Protein content in the extract was determined using spectrophotometric method (Spectrophotometer PE-5400 UV; EKROSCHEM LLC, Saint Petersburg, Russia). 34
PREPARATION OF A NON-POLAR CONCENTRATE
The biomass with disintegrated microalgae cells after protein extraction was dried at 60°C to a constant weight (Δ = 0.01 g). Extraction of total lipids was performed by adding petroleum ether to the microalgae cell biomass in the ratio of 1 g biomass:20 mL petroleum ether, extraction was carried out for 8 h using a Soxhlet apparatus (Behrotest® Extraktionsstück ez100 Dusseldorf, Germany) at 85°C. The mass fraction of extracted lipids was determined using the formula:
where ML is the mass fraction of extracted lipids; %; m1 is the mass of the dried microalgae cell biomass before extraction; g; and m2 is the mass of the dried microalgae cell biomass after extraction, g.
Qualitative and quantitative analysis of lipid mixture extracted from microalgae biomass and the separation of lipid mixture into fractions was carried out using thin layer chromatography method. 35
ANTIBACTERIAL PROPERTIES OF CHLORELLA SOROKINIANA EXTRACTS
Non-polar lipid fraction
A disc method described in 22 was used to search for a minimum concentration of non-polar lipid fraction extracted from Chlorella sorokiniana microalgae cells that would have an inhibitory effect on Gram-positive bacteria (Fig. 1).
Micrography of the Gram-positive bacterial culture Bacillus ( × 1200)
Bacteria isolated using the Koch sedimentation method 36 were used as a test culture: Gram-positive bacilli 0.5–2.5 · 1.2–10 μm in size, aerobes, forming smooth colonies of whitish color on meat-peptone agar.
To identify the bacterial sample isolated from air, DNA was extracted using the alkaline extraction method and Wizard-technology by Promega (USA); PCR fragments encoding copies of 16S rRNA genes were obtained; nucleotide sequences of 16S rRNA genes were obtained by Sanger DNA sequencing; the purity of the provided material was determined; and the most closely related nucleotide sequences of 16Sp rRNA genes were identified with the help of BLAST platform. 37 -40 The obtained bacterial culture was examined for taxonomic affiliation, it was found that the culture was not an axenic culture, the dominant component of the sample belonged to the genus Bacillus.
The minimum inhibitory concentration (MIC) was searched according to the plan in Table 1. A certain dose of non-polar lipid fraction was placed on a 5-mm disc. The discs were placed on Petri dishes with Müller-Hinton (MHA) medium (NICF, Saint Petersburg, Russia). (4 ± 0.5 mm agar layer thickness in the dish), onto which 50 μL of bacteria Bacillus (Gram-positive) with 99.9 · 106 CFU/mL concentration was inoculated. Petri dishes containing the bacterial culture and the added discs were placed in a thermostat and cultured at 37°C. Part of the Petri dishes (Table 1) were illuminated with white light with PAR = 100 ± 6 μmol photons/(m2·s). The positive control in all experiments was azithromycin (Ozone® Pharmaceutical Company, Zhigulevsk, Samara, Russia), 15 μg. Negative control was petroleum ether (EKOS-1®, Moscow, Russia), 80 μl.
Conditions for Testing the Antibacterial Efficacy of the Non-Polar Lipid Fraction 1
Values represent the mean value ± absolute error
The values of the bacterial growth inhibition zones around the substance discs presented in Table 2 are the arithmetic average of the three parallel experiments. The minimum inhibitory concentration of the lipid extract was determined using the method described in Bonev et al. 41
Antibacterial Activity of the Non-Polar Lipid Fraction 1
values represent the mean value ± standard deviation for triplicate experiments (n = 3); Note: Values with different lettering (a, b, c, d) are significantly different at p < 0.05
The study of antibiotic properties of individual fractions of non-polar extract of microalgae was carried out using the method of wells, as described in. 42 The positive control was azithromycin in the amount of 15 μg. Pure silica gel powder with 120 μL of petroleum ether was used as negative control. Each experiment was repeated three times.
Water-soluble protein fraction
The study of antibacterial properties of water-soluble protein fraction of microalgae was carried out using the well method, according to the method described in. 42 The experiment was carried out according to the scheme shown in Table 3.
Conditions for Testing the Antibacterial Efficacy of the Water-Soluble Protein Fraction 1
values represent the mean value ± absolute error
Protein hydrolysate
The study of antibacterial properties of the water-soluble peptide fraction of microalgae was carried out using the well method according to the plan described in Table 4. The peptide fraction was obtained preliminarily from the protein fraction by hydrolysis of proteins with the enzyme pepsin taken at a ratio of 1(mass unit of enzyme):100(mass units of protein). The treatment was carried out at pH 2 for 4 h. The enzyme was inactivated by heating the mixture to 85°C for 15 min. The pH was adjusted with 1M sodium hydroxide to 7.
Conditions for Testing the Antibacterial Efficacy of the Protein Hydrolysate 1
values represent the mean value ± absolute error
The values of the bacterial growth inhibition zones around the substance discs shown in Tables 5,7 are the arithmetic average of the three parallel experiments. The minimum inhibitory concentration of the peptide extract was determined using the method described in Bonev et al. 41
Antibacterial Activity of the Triacylglycerides and Fatty Acids (at PAR = 100 μmol photons/(m2 × s)) 1
values represent the mean value ± standard deviation for triplicate experiments (n = 3).
Note: Values with different lettering (a, b) are significantly different at p < 0.05
STIMULATORY PROPERTIES OF CHLORELLA SOROKINIANA EXTRACTS
In the experiment the effect of water-soluble protein extract as a component of nutrient medium on the cultivation process of Saccharomyces cerevisiae yeast was studied. 43 Dry food yeast (emulsifier 1%, instantaneous dry yeast 99%) is used for baking and beverage fermentation. A liquid nutrient medium (400 mL) containing: sucrose – 9 wt%; diammonium phosphate – 0.3 wt%; ammonium sulphate – 0.16 wt%; potassium chloride – 0.06 wt%; magnesium sulphate – 0.02 wt%. (pH 4.5–5.0), was inoculated with a suspension of Saccharomyces cerevisiae yeast, in the amount of 10% of the nutrient medium volume, as well as a water-soluble protein extract.
The flasks were then placed in a thermostat with a stirrer at 30–31°C, the suspension was barbotaged with an air-gas mixture at a flow rate of 1 L/min. Sampling was done to measure pH, cell concentration and concentration of reducing sugars every hour. Table 6 shows the conditions of the experiment. The experiment was repeated three times.
Composition of Nutrient Medium for the Cultivation of Yeast Saccharomyces cerevisiae
The number of microalgae cells in the suspension was carried out by direct counting method in a Neubauer chamber. 44 Determination of reducing sugars (glucose) concentration in culture liquid was performed using iodometric method. 45 The pH level was measured with a portable pH meter (Testo® 206 pH1, Testo SE&Co. KGaA, Titisee-Neustadt, Germany).
The calculation of specific growth rate of yeast cells was carried out using Equation 1:
where xl is the concentration of microalgae cells at 0 and n days, mln cells/mL; and t is the day of cultivation, 24 h.
Generation time was calculated by Equation 2:
where μ is the specific growth rate of yeast.
STATISTICAL ANALYSIS
Statistical analyses were performed using SPSS version 20.0. Normality of distribution of the experimental data was checked using the Shapiro-Wilk test. The significant differences between the mean values for the different experiments were determined by one-way ANOVA and post hoc test by Tukey's test at the level of significance of p < 0.05. In addition, correlation analysis was performed to examine the relationship between extract concentration (independent variable) and the diameter of the zone of inhibition (dependent variable). Pearson correlation was found between them. Experimental data presented in the tables are the mean value ± standard deviation for triplicate experiments (n = 3).
Results
ANTIBACTERIAL PROPERTIES OF CHLORELLA SOROKINIANA EXTRACTS
Non-polar lipid fraction
In the first stage of evaluating the minimum inhibitory concentration of the non-polar lipid fraction extracted from microalgae cells, it was determined that the total concentration of substances of lipid nature identified by thin-layer chromatography in the mixture was approximately (138 mg/mL): fatty acids – 35 mg/mL, triacylglycerides – 47.5 mg/mL, O-dialkylmonoglycerides – 25 mg/mL, Sterol esters or esters of waxes or trialkyl esters of glycerol – 30 mg/mL.
In the second stage of the experiment, a non-polar lipid fraction extracted from the disintegrated biomass cells of the microalgae Chlorella sorokiniana was found to have an inhibitory effect on the growth of Gram-positive bacteria when illuminated with white light (PAR) 100 ± 6 μmol photons/(m2·s), the minimal inhibitory concentration of the non-polar lipid fraction was ≈80 μg/mL (R 2 = 0.85). In darkness (PAR = 0 μmol photons/(m2·s)), there was no inhibitory effect even when the maximum amount of non-polar substances 7,590 μg/mL was applied to the disk (Table 2).
The experiment to determine the minimum concentration of individual fractions of substances of non-polar extract from Chlorella sorokiniana biomass which inhibits Gram-positive bacteria of the Bacillus genus showed that only triacylglycerides and fatty acids of Chlorella sorokiniana non-polar extract have antibiotic properties (Table 5). The mixture of substances of lipid nature of Chlorella sorokiniana non-polar extract has a more pronounced antibiotic effect than individual fractions of substances at PAR level 100 ± 6 μmol photons/(m2·s).
WATER-SOLUBLE PROTEIN FRACTION
Evaluation of the antibacterial properties of a water-soluble protein fraction of the microalgae revealed that the water-soluble protein fraction extracted from the microalgae biomass has no antibacterial properties either in the dark or in the light.
PROTEIN HYDROLYSATE
The results of the experiment to evaluate the antibacterial properties of a water-soluble peptide fraction of the microalgae, presented in Table 7, suggest that the water-soluble peptide fraction extracted from the cells of the microalgae Chlorella sorokiniana has an inhibitory effect on the growth of Gram-positive bacteria Bacillus under both white light PAR = 100 ± 6 μmol photons/(m2·s) and darkness (0 μmol photons/(m2·s)), MIC of the peptide fraction is ≈125 μg/mL when illuminated and 170 μg/mL in the dark.
Antibacterial activity of the protein hydrolysate 1
values represent the mean value ± standard deviation for triplicate experiments (n = 3); Note: Values with different lettering (a, b, c, d, e) are significantly different at p < 0.05
STIMULATORY PROPERTIES OF CHLORELLA SOROKINIANA EXTRACTS
Having analyzed the results of the experiment on the evaluation of stimulating properties of the water-soluble protein fraction of the microalgae on the growth of eukaryotic cells ( Figs. 2,3,5 ), we can see that cultivation of Saccharomyces cerevisiae yeast on nutrient medium with addition of water-soluble protein extract of Chlorella microalgae in an amount of 0.005 and 0.01 mL per 1 mL of the nutrient medium, samples 3 and 4, differed significantly from the control sample (without adding the extract) and sample 2 (0.0025 mL of the water-soluble extract per 1 mL of the nutrient medium).
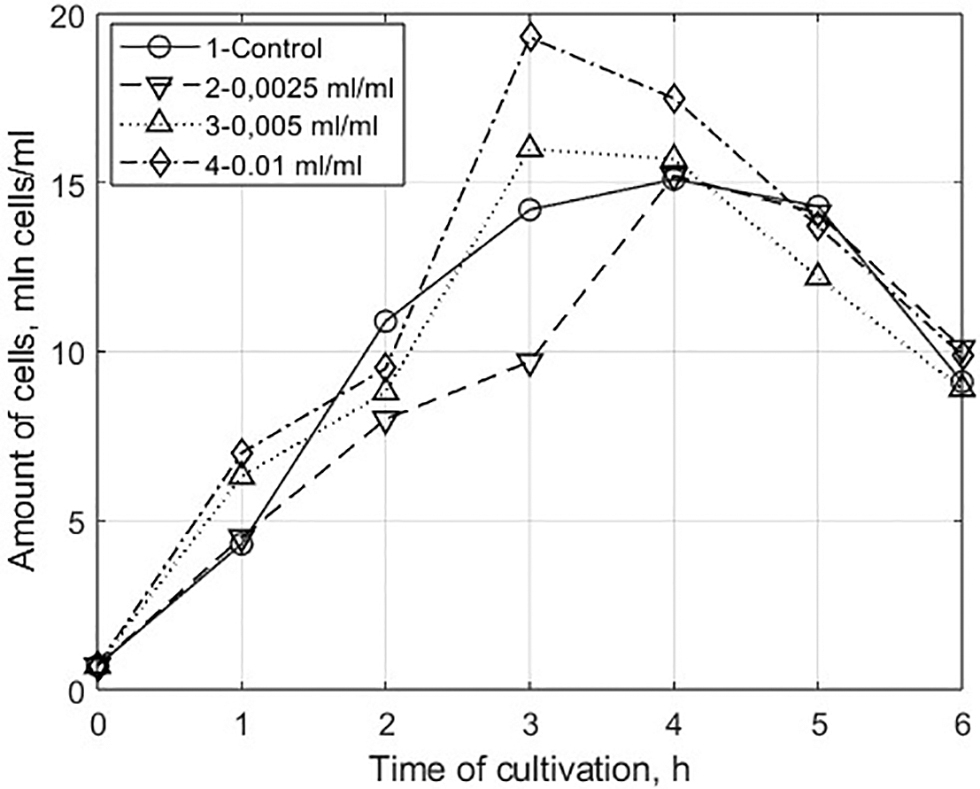
Changes in the concentration of cells
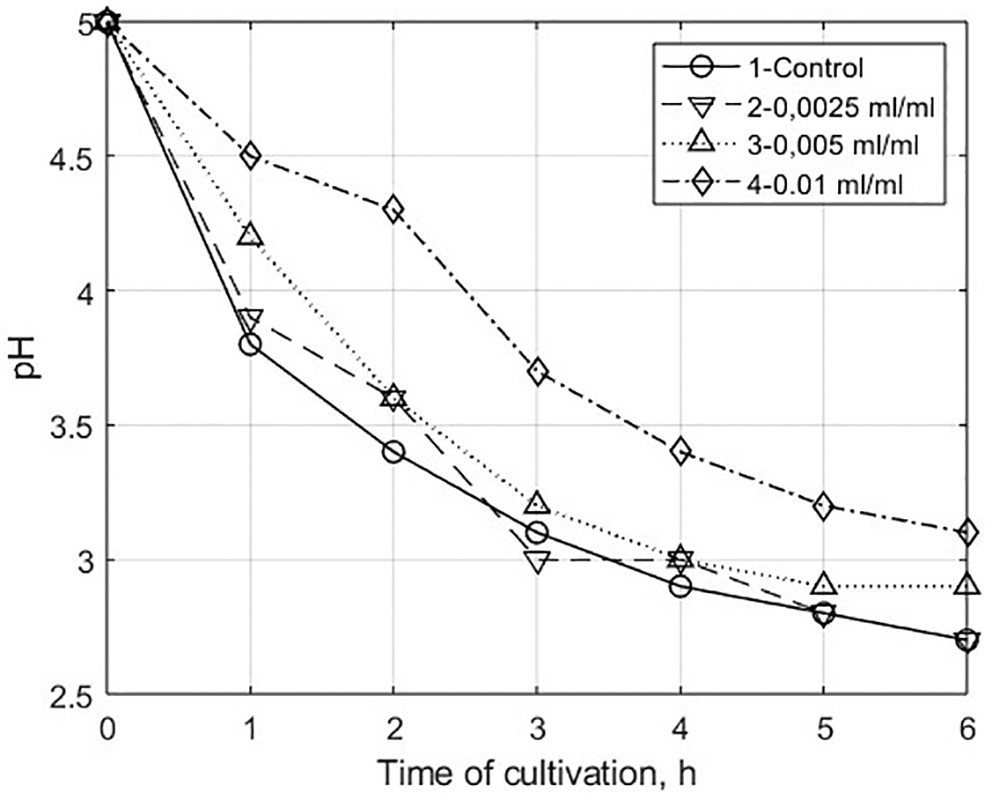
Changes in the acidity of the nutrient medium
The population capacity of samples 3 and 4 increased by 5% and 21%, respectively, compared with the control sample (Fig. 2). At the same time, the value of specific growth rate in the exponential phase increased by 1.3–1.4 times, and the time of cell generation decreased by 23–28%, respectively (Table 8).
Cultivation Parameters of the Yeast Saccharomyces cerevisiae (Exponential Phase) 1
Values represent the mean value ± standard deviation for triplicate experiments (n = 3); Values with different lettering (a, b) are significantly different at p < 0.05
It is important to note that the length of the exponential phase of samples 3 and 4 decreased to 3 h (Fig. 2) compared with the control. This is explained by the fact that the stimulating agents of Chlorella extract accelerate metabolic processes in cells and lead to an earlier accumulation of inhibitory metabolites (ethanol, acetate), high concentrations of which inhibit cell growth. 46,47
Discussion
The experimental results show that the antibacterial effect of the non-polar lipid fraction extracted from the cells of the microalgae Chlorella sorokiniana on Gram-positive bacteria was evident in the presence of light. Analysis of the chemical structure of the components of the extracted fraction (fatty acids; triacylglycerides; O-dialkylmonoglycerides; sterol esters or wax esters or glycerol trialkyl esters) suggested the presence of a large number of double bonds in these compounds, where the oxidation of these substances under the influence of light radiation was observed. This assumption agrees with the Bach-Engler theory of lipid peroxidation and with the theory of branched chain reactions by N.N. Semenov. 23 The lipid radicals formed may attack protein and nucleic acid molecules and oxidize cytoplasmic membrane lipids, which causes metabolic disruption and death in bacteria. 23
The peptide fraction was produced by treating the water-soluble fraction of proteins with pepsin, and 24–30% of this fraction of autotrophic microorganisms, in particular Chlorella microalgae, consisted of the RUBISCO enzyme.
48
By analyzing the amino acid sequence of the RUBISCO enzyme (isoform A), which is characteristic of Chlorella sorokiniana microalgae cells, and considering that the enzyme pepsin breaks peptide bonds that contain the amino groups of leucine, phenylalanine, tryptophan and tyrosine, we could predict which peptides would predominate in that mixture.
49,50
Using the PeptideCutter (
These properties allow the peptides to interact with the cytoplasmic membranes of bacteria, which are composed of phospholipids (Fig. 4). 52,53 It is assumed that antibacterial properties of the peptides are based on the following mechanism: peptides penetrate into the lipid matrix of cell membranes and form pores there through which intracellular contents leak into the intercellular space, although some peptides have intracellular targets. 15,54 Even in the latter case, the peptide-membrane interaction is important for peptide penetration into the cell. Peptides containing large amounts of positively charged amino acids (arginine, histidine, lysine) can integrate into the negatively charged cytoplasmic membrane of a bacterial cell (bacterial cell membranes have a larger negative charge). 55
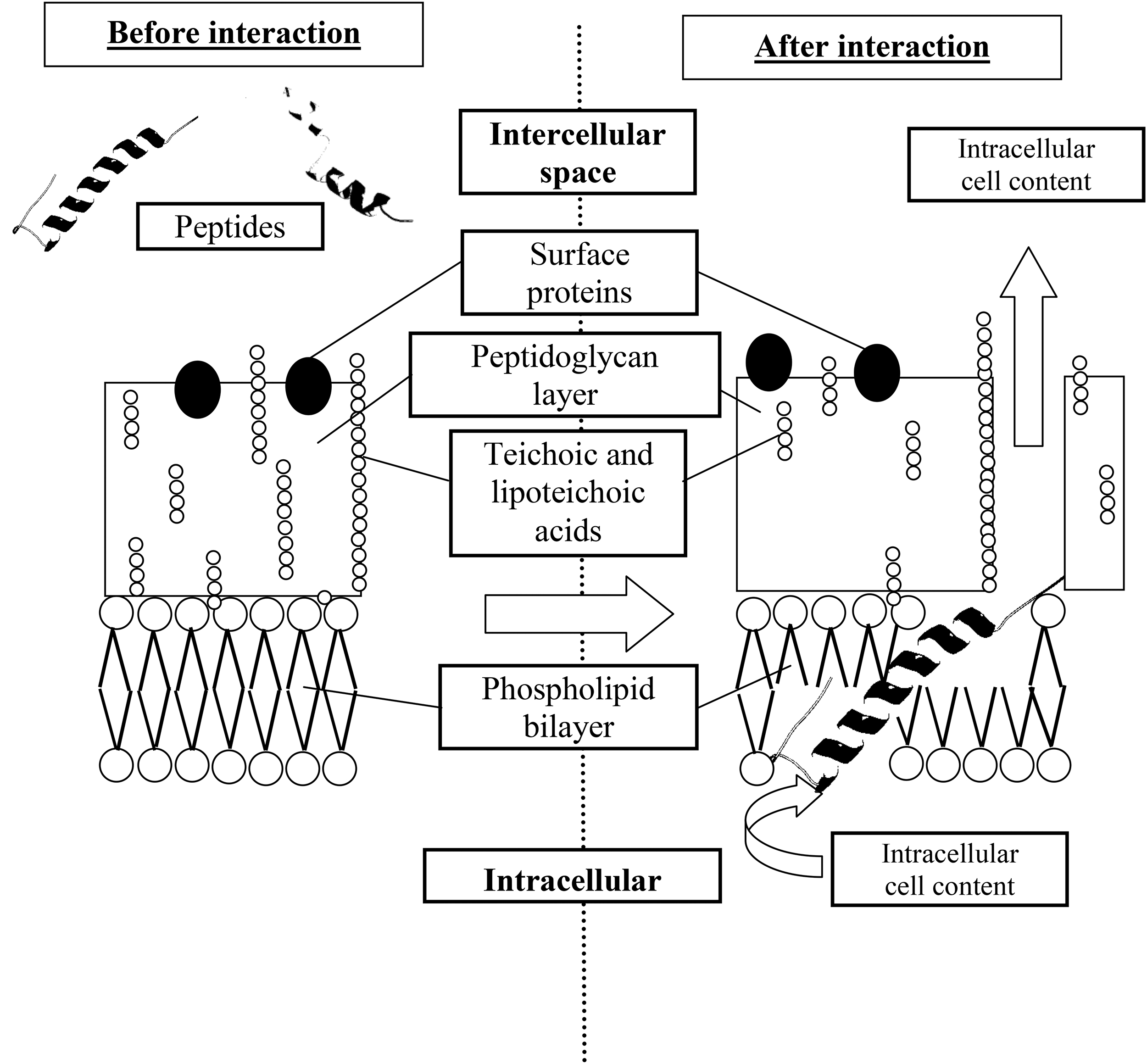
Putative mechanism of the antibiotic action of the microalgae peptide fraction.
The use of Chlorella aqueous extract as a growth stimulant, in addition to protecting against negative effects from toxic extracellular metabolites, stimulated various metabolic processes in Saccharomyces cerevisiae yeast cells. 25 This was confirmed by more effective conversion of sucrose into reducing sugars (glucose and fructose) in samples 3 and 4 (Fig. 5).

Changes in the concentration of reducing sugars.
Conclusion
A mixture of substances extracted from the disrupted Chlorella sorokiniana microalgae cells with a non-polar solvent had an inhibitory effect on bacterial growth at PAR levels of 100 ± 6 μmol photons/(m2·s). The minimum inhibitory concentration of the extract was ≈80 μg/mL (R 2 = 0.85). When analyzing the chemical structure of the components of the non-polar fraction extracted from the cells of the microalgae Chlorella sorokiniana, it was found that the non-polar extract includes triacylglycerides, fatty acids, O-dialkyl monoglycerides, sterol esters or wax esters or trialkyl glycerol esters. As far as the isolated components of the mixture were concerned, triacylglycerides with MIC = 176 μg/mL (R 2 = 0.96) and fatty acids with MIC = 445 μg/mL (R 2 = 0.98) exhibited antibacterial properties.
A water-soluble peptide fraction extracted from cells of the microalgae Chlorella sorokiniana had an inhibitory effect on the growth of Gram-positive bacteria both in white light PAR = 100 ± 6 μmol photons/(m2·s), and in darkness (0 μmol photons/(m2·s)), )), MIC of the peptide fraction was ≈125 μg/mL (when illuminated) and 170 μg/mL (in the dark).
The water-soluble protein fraction had no antibacterial properties but is of interest as a component of nutrient medium for the cultivation of the yeast Saccharomyces cerevisiae. Adding this fraction in an amount of 0.01 mL/mL of nutrient medium increased the population capacity by 21% compared with the control sample, boosting the specific growth rate in the exponential phase by 1.4 times and reducing the time of cell generation by 28%.
Footnotes
Author Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by [Mikhail S. Temnov], [Yana V. Ustinskaya] and [Kirill I. Meronyuk]. The first draft of the manuscript was written by [Mikhail S. Temnov] and [Dmitry S. Dvoretsky] and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.