
Abstract
Strong mineral acids such as sulfuric acid are still mainly used to break down carbohydrates in processes that prepare plant-based waste for further use. However, researchers are increasingly studying the use of organic acids as alternatives. This study aimed to obtain organic compounds by fermenting a hemicellulosic extract rich in simple sugars from the organic acid pretreatment of brewer’ spent grain. Hydrolysis tests using 4.5% oxalic acid for 24 minutes at 125.6°C dissolved over 99% of the hemicellulose, producing 14.9 g/L of xylose and 4.9 g/L of glucose. The production of inhibitory compounds was relatively low (0.05 g/L and 0.12 g/L of 5-hydroxymethylfurfural and furfural, respectively), which is beneficial for the yeast’s performance during the next step. The extract was then fermented by three strains of Saccharomyces cerevisiae: one industrial strain (PE-2) and two genetically modified strains (MP-P5 and JDY-01). Despite the hydrolysate containing around 45 g/L of oxalic acid, which significantly inhibited yeast activity in producing ethanol, for example, the three strains still produced several important volatile organic compounds, including 2-phenylethanol, known for its antimicrobial properties, and 4-hydroxy-2-methylacetophenone, a phenolic compound with antioxidant and antiseptic properties.
Introduction
Over the past decades, lignocellulosic biomass from various plants has been studied as a renewable resource for producing environmentally sustainable energy and value-added products. Despite its large-scale production across many economic sectors, much of this waste is currently burned, used as animal feed, or disposed of in landfills. 1 The global production of lignocellulosic biomass is estimated at around 181.5 billion tons annually. 2 Lignocellulosic material mainly consists of cellulose, hemicellulose, and lignin, arranged in a highly stable crystalline structure that must be broken down through pretreatments to isolate the desired fractions for further processing. 3 –5
Brewer's spent grain, a byproduct of the brewing process, is composed mainly of malt husk and pulp, remaining after the filtration of beer wort before boiling. This residue has significant potential for reuse, with approximately 20 kg produced for every 100 L of beer. In 2022, global beer production reached about 189 billion liters, generating an estimated 37 million tons of brewer’s spent grain. 6 Although primarily used as low-value animal feed, this biomass has promising characteristics, being rich in carbohydrates such as cellulose and hemicellulose, which can be transformed into value-added products through biotechnological processes. 7 –10
A myriad of products can be derived from the fermentation of sugars present in the lignocellulosic matrix of biomass. The hemicellulose of malt bagasse is rich in pentoses, such as xylose and arabinose, which can serve as raw materials for the production of xylooligosaccharides, 11,12 xylitol, 13,14 biofilms/bioplastics, 15,16 enzymes, 12,17 and furan derivatives. 12,18 Additionally, through genetic modification of traditional yeasts such as Saccharomyces cerevisiae, ethanol can be produced from pentoses as efficiently as it is from glucose. 19
However, to obtain these value-added products, biomass pretreatment is a critical step in the biorefinery process, as it modifies the lignocellulosic structure to access its main components. 20,21 Several methods can be employed, including acid hydrolysis (primarily using mineral acids such as H2SO4 and HCl), 22 alkaline hydrolysis (usually with NaOH), enzymatic hydrolysis, and autohydrolysis (also known as hydrothermal treatment). To optimize these processes, key variables such as temperature, pressure, reagent concentration, and time must be carefully controlled. 7,23
A key challenge in the pretreatment stage is to effectively disrupt the lignocellulosic matrix to increase contact area and solubilize its components while minimizing the formation of fermentation-inhibiting compounds such as furfural, 5-hydroxymethylfurfural (HMF), and acetic acid. Strong mineral acids can damage biorefinery equipment, causing corrosion and disposal issues. Therefore, research has focused on using less aggressive organic acids, which can reduce the formation of inhibitory compounds and minimize damage to equipment. 24 –27 However, the effect of the use of organic acids, such as oxalic acid, on the solubility and fermentability of carbohydrates present in biomass is still inconclusive.
Oxalic acid is a dicarboxylic organic acid (pKa1 = 1.25, pKa2 = 4.27) with a low molecular weight (≤ 300 g/mol) and is found in nature as either the acid or its conjugate base, oxalate. 28 It offers several advantages for pretreating lignocellulosic waste: it produces fewer inhibitory compounds that could affect fermentation, such as 5-HMF and furfural, can be optimized for various types of biomasses, and selectively hydrolyzes the lignocellulosic matrix with minimal degradation of cellulose and hemicellulose. Unlike mineral acids such as sulfuric acid, oxalic acid’s recovery is easier through standard organic acid techniques, such as ion exchange and adsorption, which, despite its higher cost, makes its use more feasible. 29 Oxalic acid has proven effective for pretreating a range of biomasses, including oil palm, 30 yellow poplar, 29 corn cobs, 31 maple wood, 32 and olive biomass. 33
Therefore, this study aims to evaluate using oxalic acid, an organic acid, as a replacement for traditional mineral acids in the pretreatment of brewers’ spent grain. The goal is to produce a hydrolysate rich in pentoses while minimizing inhibitor formation. The hydrolysate will then be fermented using three S. cerevisiae strains: the industrial strain PE-2 and two genetically modified strains, MP-P5 and JDY-01, to assess carbohydrate consumption and the production of organic compounds.
Materials and Methods
BIOMASS
The brewer's spent grain was sourced from pilsner malt and supplied by Basaltica Cervejaria (Chapecó, SC, Brazil). After brewing, the material was collected from a single batch and stored in airtight bags. The malt bagasse was transported to the laboratory and dried in an air-circulating oven at 55 ± 1°C until constant weight (approximately 72 hours). After drying, the biomass was ground using a Willye knife mill to achieve particles with a maximum diameter of 0.6 mm (AmericanLab model AL-032S). The conditioned biomass was stored in hermetically sealed packaging at a laboratory temperature of approximately 20°C. All tests were performed on a single batch of bagasse.
ACID PRETREATMENT
An acid pretreatment was performed to enhance the solubilization of xylose from brewer's spent grain. Three concentrations of oxalic acid were initially tested: 3% m/v (OA3%), 4% m/v (OA4%), and 5% m/v (OA5). Additionally, a sulfuric acid concentration of 1% v/v (SA1) was tested under the same experimental conditions described by Dresch et al. 34
Briefly, 20 g of biomass was processed at a solid-to-liquid ratio of 1:10. The vials were sealed and autoclaved for 120 minutes at 126°C and 1.4 bar. Afterward, the sample was vacuum filtered using a Büchner funnel. The liquid fraction was analyzed for carbohydrates, acetyl groups, and inhibitors via high-performance liquid chromatography (HPLC; Shimadzu, Brazil). The solid fraction was resuspended in 200 mL of distilled water, neutralized with 12% (m/v) sodium hydroxide (NaOH), filtered, and dried in an oven at 105°C until constant weight. The solid fraction was then subjected to physicochemical characterization.
Based on the optimal results from these preliminary pretreatments, a series of tests were designed to assess the effects of different oxalic acid concentrations (3.5–5.5%) and reaction times (24–136 minutes; Table 1). The experimental methodology used was identical to that of preliminary pretreatment. The xylose yield in the hydrolysate and the formation of inhibitory compounds (e.g., furfural, 5-HMF, and acetic acid) were evaluated.
Variables and Levels Employed in the Rotational Central Composite Design for the Biomass Pretreatment
BREWER'S SPENT GRAIN FERMENTATION
Fermentation tests were conducted using three strains of S. cerevisiae: PE-2, MP-P5, and JDY-01. The MP-P5 and JDY-01 strains are genetically modified derivatives of PE-2, as previously described by Tadioto et al. 19 Preinoculation of the three yeast strains followed the methodology outlined by Vargas et al. 35
To evaluate the effects of fermentation temperature and hydrolysate pH on carbohydrate consumption, xylitol accumulation (by the strain PE-2), ethanol production (by the strain MP-P5 and JDY-01), and volatile organic compounds (VOCs) production (by all three yeast strains), a 22 rotational central composite design (RCCD) was conducted in triplicate at the central point (Table 2).
Variables and Levels of the RCCD for the Fermentation Tests with the Saccharomyces cerevisiae Strains: PE-2, MP-P5, and JDY-1
The hemicellulosic hydrolysate was initially concentrated to approximately 50% of its original volume using a vacuum rotary evaporator (TE-213, Tecnal, Piracicaba, SP, Brazil). The concentrated hydrolysate was supplemented with 1% yeast extract and 2% bacteriological peptone, neutralized with a 2 M NaOH solution, and filtered through a sterile 0.22 μm membrane using a vacuum filtration system. A 50 mL aliquot of this prepared hydrolysate was transferred to 250 mL Erlenmeyer flasks, inoculated with 1 μL of preinoculum from each yeast strain, and incubated for 48 hours at 145 rpm on a rotary shaker under the temperature and pH conditions outlined in the experimental design (Table 2).
Samples were collected in duplicate at zero time and every 12 hours. The collected samples were centrifuged at 9,000 rpm for 5 minutes, and the resulting supernatants were filtered through 0.22 μm filters. The filtered supernatants were subjected to HPLC to analyze carbohydrate consumption, xylitol, and ethanol production.
Quantification of VOCs
To determine VOCs, the fermented hydrolysates were transferred to Falcon tubes and centrifuged at 9,000 rpm for 10 minutes. A 20 mL supernatant sample was collected and mixed with 6.67 mL of dichloromethane (1/3 v/v). The mixture was agitated for 30 minutes at 145 rpm at room temperature. A liquid–liquid extraction was then performed using a separation funnel, following the methodology proposed by Roque et al. 36
The extracted samples were analyzed using gas chromatography coupled with mass spectrometer (GC-MS), model GCMS-QP2010Ultra (Shimadzu). A 2-μL sample aliquot was injected into an NST05ms capillary column (30 m × 0.25 mm × 0.25 μm), with helium as the mobile phase at a 14 mL/minute flow rate. The injector temperature was set at 150°C, with a split ratio of 1:10. The oven temperature was programmed as follows: 100°C for 6 minutes, then ramped from 100°C to 150°C at a rate of 4°C/minute, followed by an increase from 150°C to 220°C at a rate of 8°C/minute, and held at 220°C for up to 40 minutes. The mass spectrometer operated with an ionization source temperature of 200°C and an interface temperature of 220°C in scanning mode (m/z range: 35–500). 37 The NIST08s library was used for mass spectrum analysis. Results were reported with a limit of 50 peaks, presenting the area of each peak for compounds with a retention time in the column. The difference between the compounds in the culture medium before inoculation was also evaluated.
The production of each compound was determined semiquantitatively through the areas of each peak, which represent the relative abundance of each substance to the others, as described by Di Francesco et al. 38
BIOMASS CHARACTERIZATION
The moisture, ash, extractives, total lignin, cellulose, and hemicellulose contents were quantified following the National Renewable Energy Laboratory (NREL) methods. 39 The moisture content was determined by drying 6 g of biomass in an oven at 105°C until a constant weight was achieved. 40 The ash content was determined by incinerating 1 g of the sample at 800°C for 2 hours. 41 The extractive content was assessed using Soxhlet extraction, where the sample was sequentially washed with water and ethanol to isolate water-soluble and ethanol-soluble components. 41
Cellulose, hemicellulose, and lignin were extracted and analyzed using the NREL protocol described by Sluiter et al.
39
Briefly, 0.3 g of extractive-free biomass was hydrolyzed with 3.0 mL of 72% (v/v) sulfuric acid (H2SO4) in a water bath at 30°C for 1 hour. The samples were diluted to 4% (v/v) sulfuric acid by adding 84 mL of deionized water, autoclaved at 121°C for 1 hour, and vacuum filtered. The solid fraction, representing acid-insoluble lignin, was washed with 500 mL of deionized water and dried at 105°C to a constant weight. The liquid fraction was divided into two parts: one aliquot was used to measure acid-soluble lignin using a UV-Vis spectrometer (UV-1800, Nova Instruments, Brazil). The remaining portion was neutralized with calcium carbonate powder to a pH of 5–6, filtered through 0.2 µm syringe filters, and analyzed by HPLC for carbohydrates, inhibitors, acetyl groups, and acid-soluble lignin.
34,35
The proportions of cellulose and hemicellulose were estimated using Equations 1 and 2, respectively.
Protein content was measured using the Kjeldahl method, as detailed in the Adolfo Lutz Institute Manual.
42
Briefly, 0.2 g of biomass was added to digestion tubes containing 1 mL of H2O2, 2 mL of H2SO4, and 0.7 g of a digestion mixture (100 g of Na2SO4; 10 g of CuSO4; and 1 g of selenium). The mixture was digested at 350°C until a clear green coloration. Following digestion, the sample was diluted to 25 mL in a volumetric flask, neutralized with NaOH (10 mol/L), and distilled using a nitrogen distiller (Marconi, Model MA-036, Brazil). Nitrogen content was quantified through titration with H2SO4 (0.025 mol/L) using a boric acid solution (H3BO3) as the indicator. For the blank, a test was performed without biomass. The percentage of protein was calculated using Equation 3.
STATISTICAL ANALYSIS
The results obtained were evaluated using analysis of variance, followed by Tukey’s test with a 95% confidence interval (p < 0.05) in GraphPad Prism Software (version 9.5.1, StatSoft Inc.). The experimental design and RCCD were structured and analyzed using Protimiza Experimental Design® Software (Campinas, SP, Brazil, version: https://experimental-design.protimiza.com.br).
Results and Discussion
BREWER'S SPENT GRAIN CHARACTERIZATION
The physicochemical composition of the brewer’s spent grains used in this study is shown in Table 3. The composition of malt bagasse is dependent upon the variety of barley utilized and the processing techniques employed during beer production, with values ranging from 3.48% to 30.00%. 43 –46 In this study, the brewery spent grain exhibited a relatively higher lignin content, accounting for 32.22% of its total composition. Such discrepancies can be attributed to variations in malt varieties, brewing processes, and methodologies employed, which have evolved alongside advancements in quantification methods, instrumentation, and techniques. Furthermore, the lignin content of various biorefining feedstocks, including barley, a source of malt bagasse, can be significantly impacted by extreme temperatures, nitrogen-rich nutrition, and water scarcity during cultivation. 47
The Physicochemical Composition of the Malt Bagasse Used in this Study Compared with that Reported in Other Studies Found in the Literature
Sum of acid-soluble lignin and acid-insoluble lignin.
The cellulose content observed in this study (18.34%) is consistent with the value reported by Ribeiro-Sanches et al. 46 which was 17.5%. Similarly, the hemicellulose content aligns with the findings reported in the literature (Table 3). Hemicellulose is composed of pentose sugars, including xylose and arabinose, which are valuable compounds for second-generation biorefineries. In these facilities, microorganisms can convert these monomers into value-added products, such as xylooligosaccharides, 11 xylitol, 13 and furan derivatives, 18 supporting the circular economy.
Additionally, brewer's spent grain is a rich source of protein, particularly essential amino acids, with a protein content that typically ranges from 14.00% to 30.00%. Consequently, it is predominantly employed in the food industry. 48 –50 The biomass analyzed in this study exhibited a total protein content of approximately 14.15%, which is consistent with the values reported in the literature. Further investigation into the potential of isolating protein from malt bagasse for use in human food, particularly as a substitute for animal-derived protein isolates commonly used in dietary supplements, is recommended, particularly given the high availability of this byproduct. 51
PRETREATMENT WITH OXALIC ACID TO SOLUBILIZE HEMICELLULOSES
To assess the solubilization of hemicellulose, preliminary tests were initially performed utilizing three concentrations of oxalic acid (3%, 4%, and 5%, m/v) and one concentration of sulfuric acid (1%, v/v). The results are presented in Figure 1.
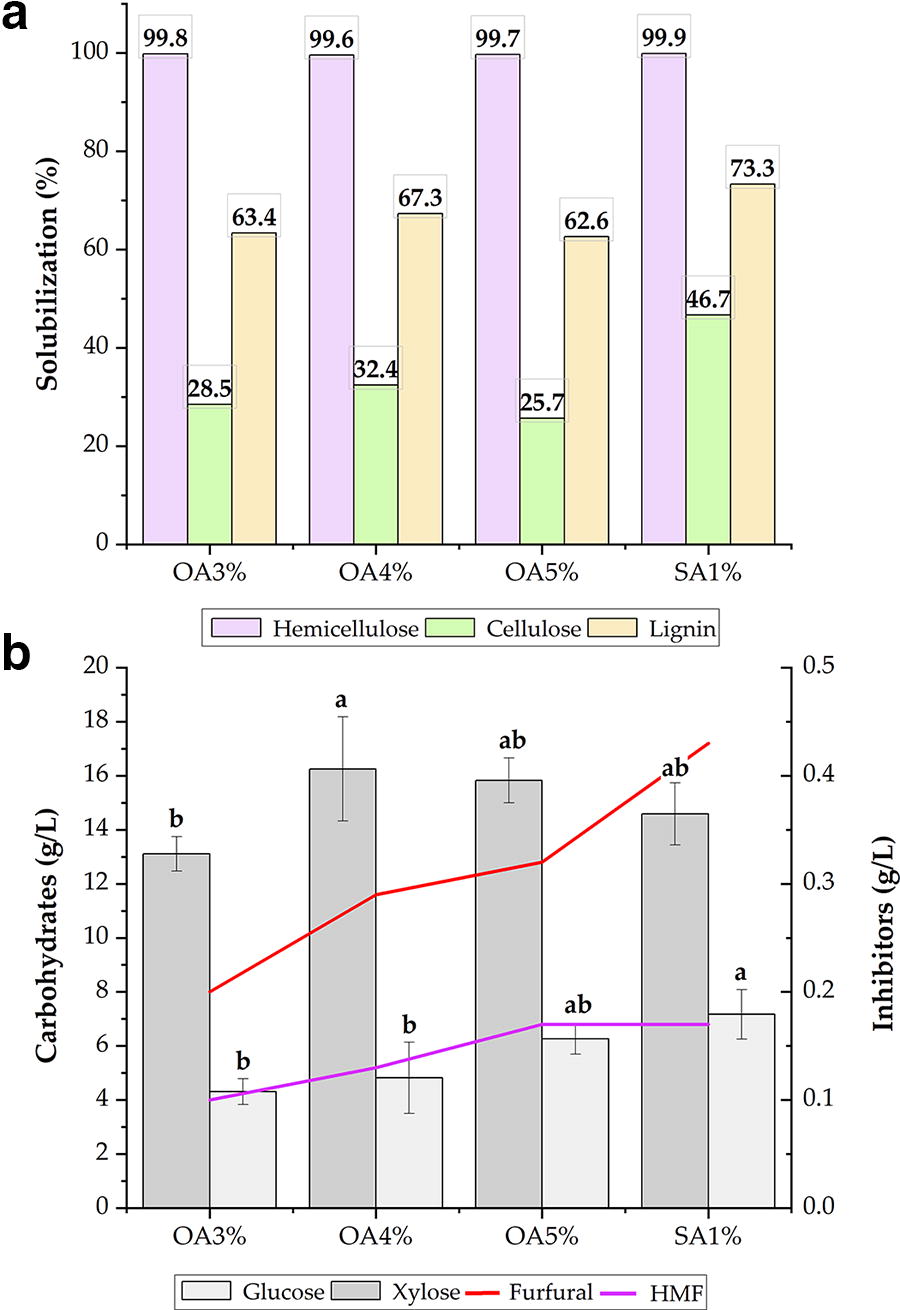
The results are presented in terms of the solubilization of hemicellulose, cellulose, and lignin
All tests demonstrated solubilization efficiency of over 99.0% for the hemicellulose in malt bagasse (Fig. 1a), resulting in hydrolysates with xylose concentrations ranging from 13 g/L to 16 g/L (Fig. 1b). The oxalic acid treatments at concentrations of 4% and 5% yielded the most favorable results in terms of xylose concentration in the medium. Furthermore, the oxalic acid treatments resulted in significantly lower levels of inhibitory compounds compared with sulfuric acid treatment, particularly HMF, which reached 0.32 g/L in the hydrolysis with 5% oxalic acid, compared with 0.43 g/L in the sulfuric acid treatment at 1%. In a similar study, Wilkinson et al. 52 reported hydrolysates with 0.44 g/L of HMF and 0.41 g/L of furfural in pretreatments using 1% hydrochloric acid. This finding demonstrates that oxalic acid is considerably less aggressive in degrading carbohydrates in biomass than other acids, such as hydrochloric acid. The formation of these compounds is closely related to the reaction time, as pentoses and hexoses are exposed to acid after solubilization, which results in their degradation.
A study conducted by Sanchez and Bautista 53 demonstrated that furfural exerts a pronounced inhibitory effect on the fermentative and respiratory metabolism of the yeast S. cerevisiae at concentrations of 1.5 g/L, resulting in complete inhibition at this level. Concentrations over 2 g/L have been demonstrated to induce toxicity in furfural comparable to that observed with HMF. Furfural is typically present in higher concentrations than HMF in most lignocellulosic hydrolysates and has been observed to exert a more pronounced toxicity effect on the majority of industrial microorganisms. Furthermore, furfural has been demonstrated to potentially enhance the toxicity of other compounds in culture media. 54 Consequently, the concentrations of furfural and HMF in all pretreatments can be regarded as low.
The most favorable results for the preliminary tests on hemicellulose solubilization were observed when an oxalic acid solution was employed at concentrations between 4% and 5%. Subsequently, pretreatment trials were conducted on malt bagasse to assess the impact of varying oxalic acid concentration on the pretreatment process. The variables under investigation included the concentration of oxalic acid (3.5–5.5%) and the reaction time (24–136 minutes). The results demonstrated that all tests successfully solubilized more than 99% of the hemicellulose present in the malt bagasse (Fig. 2a). However, some tests yielded a greater quantity of inhibitory compounds as the reaction time increased (Fig. 2b).
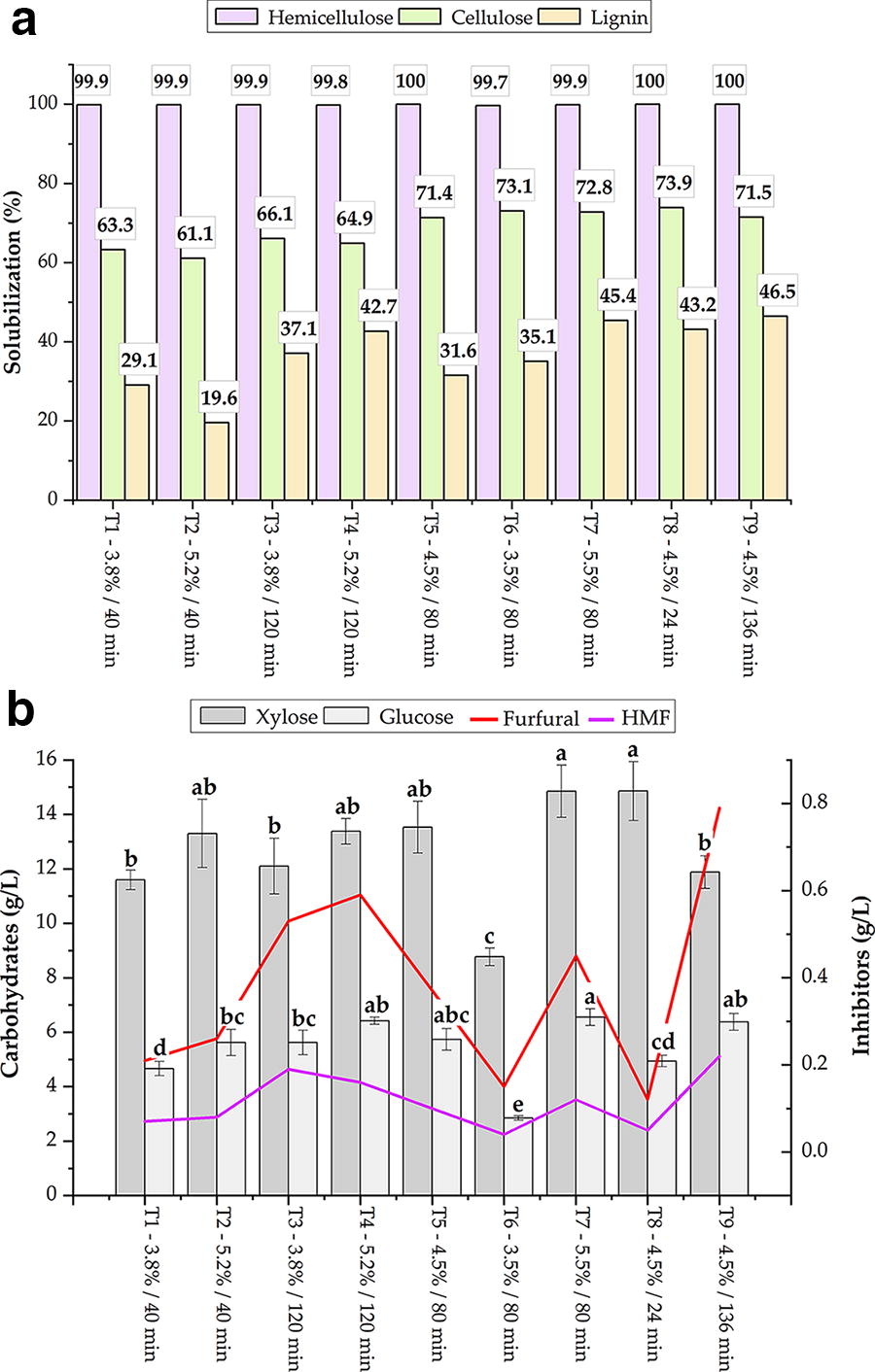
Results of RCCD for the biomass pretreatment with oxalic acid in terms of the solubilization of hemicellulose, cellulose, and lignin
With regard to the concentration of xylose in the hydrolyzate, the tests that yielded the most promising results were T8 (4.5% oxalic acid; 24 minutes of hydrolysis) and T7 (5.5% oxalic acid; 80 minutes of hydrolysis). Both tests yielded comparable results in this regard, with an average xylose concentration of 14 g/L. Bukhari et al. 30 reported similar results when treating oil palm trunk biomass with 4% oxalic acid, resulting in a hydrolysate with approximately 15 g/L of xylose. It is crucial to consider the potential impact of inhibitory substances on the fermentation process. The furfural and HMF, which are formed during the degradation of xylose and glucose, respectively, have the potential to inhibit microbial activity. In this context, the T8 demonstrated the lowest levels of furfural and HMF formation levels, with 0.12 g/L and 0.05 g/L, respectively. This can be attributed to the shorter reaction time of 24 minutes, which limited the exposure of solubilized hemicellulose to prolonged high temperatures and oxalic acid. This, in consequence, reduced the degradation of xylose and glucose monomers into inhibitory compounds.
Given that there was no significant difference in xylose solubilization and concentration between the tests, and the fact that the concentration of inhibitory compounds remained below levels that could potentially impair fermentation, it was decided to proceed under the conditions of the T8 test: the solution was autoclaved at 126°C and 1.4 bar for 24 minutes, using 4.5% oxalic acid.
FERMENTATION AND EVALUATION OF ORGANIC COMPOUND PRODUCTION BY THE YEASTS
Consumption of carbohydrates and production of xylitol and ethanol
Following concentration in a rotary evaporator, the hydrolysate intended for fermentation exhibited a glucose concentration of 5.4 g/L, a xylose concentration of 22.98 g/L, and an arabinose concentration of 14.09 g/L. Furthermore, the hydrolysate exhibited minimal inhibitory effects, with concentrations of 1.12 g/L of acetic acid, 0.01 g/L of furfural, and 0.06 g/L of HMF. Acetic acid has been demonstrated to exert a pronounced inhibitory effect on microbial growth at concentrations exceeding 5 g/L. 55 However, the concentrated hydrolysate intended for fermentation exhibited an average acetic acid content of 1.12 g/L, which falls well below the critical threshold. Consequently, it was concluded that the fermentation tests could be conducted without significant issues. Furthermore, the concentration of furfural was observed to decrease following the hydrolysate concentration stage. This reduction may be attributed to the evaporation of furfural during the concentration process, as a consequence of the reduced vapor pressure exerted by the rotary evaporator.
In the majority of experiments, fermentation tests conducted with the three S. cerevisiae strains did not result in the production of ethanol or xylitol. The only exception was experiment F12, in which the MP-P5 strain produced 1.76 g/L of ethanol following a 48-hour fermentation period. At the outset of the experiment, the yeast consumed the entirety of the available glucose, as anticipated, given its proclivity for this carbohydrate. 56 Over the course last 12 hours, it commenced the consumption of xylose (approximately 1.0 g/L; Table 4). The introduction of the requisite enzyme-encoding genes into the genome of this strain via genetic modification enables it to ferment xylose. 19
Results of the Initial and Final Concentrations of Glucose, Xylose, and Arabinose for the Three Saccharomyces cerevisiae Strains (PE-2, MP-P5, and JDY-01)
Tests that present different superscript letters for the initial and final sugar concentration differ significantly (p < 0.05).
The ethanol yield of the F12 test was 0.35 gethanol/gglucose/xylose, which is below the theoretical maximum of 0.51 gethanol/gglucose. 57 This lower yield was expected, as an external factor may have impaired fermentation, and achieving the maximum yield is challenging even under optimal conditions. It was not expected that this strain would produce xylitol, given that its genetic modifications were designed to facilitate ethanol production from pentose consumption. By the conclusion of the experiment, approximately 26 g/L of sugar remained in the medium. It is important to note that if the fermentation had been extended beyond 48 hours, it is possible that all carbohydrates would have been consumed.
Additionally, the test demonstrated a notable elevation in the glycerol concentration within the medium, from 0.42 g/L to 0.90 g/L. The observed increase in glycerol concentration may be attributed to stress induced by low pH, which has the potential to damage the cell wall and trigger defense mechanisms. Such mechanisms include the activation of the cell wall integrity pathway and the high-osmolarity glycerol pathway. The latter pathway involves the accumulation of glycerol within the cell, which helps to maintain cellular tone. 58 –60
The tests F1 and F11, utilizing yeast PE-2, F20, F21, and F22 with yeast MP-P5, and F23, F31, F32, and F33 with yeast JDY-01, demonstrated a slight decline in glucose concentration over the 48-hour fermentation period. A t-test revealed that the initial and final glucose values were significantly different, indicating that the yeast may have consumed glucose while adapting to the environment. However, this adaptation, which is referred to as the lag phase, did not result in the production of ethanol. The duration of the lag phase can vary considerably, depending on the specific environmental conditions to which the microorganism is exposed. 61 It is important to note that an extended lag phase does not necessarily indicate cellular stress. As demonstrated by Hamill et al., 62 long lag phases are a typical feature of native species and are more likely to be related to thorough environmental adaptation than to cellular stress.
The tests that did not yield ethanol or xylitol and exhibited no carbohydrate consumption were likely influenced by factors that inhibited yeast activity. These factors may include the initial pH of the hydrolysate and the residual presence of oxalic acid, which was used in the pretreatment step, or its conjugate base, oxalate. The presence of oxalate may be attributed to the pH adjustment of the medium, which was performed in accordance with the experimental design.
The pH of the medium is a critical factor affecting the cellular and metabolic viability of yeast during fermentation. The majority of strains of S. cerevisiae demonstrate growth at pH values between 2.5 and 8.5, with the specific range dependent on factors such as temperature, oxygen presence, and the particular strain under consideration. The optimal pH range for yeast growth is between 4.0 and 6.0, with enhanced yields frequently observed in acidic environments. 63 It has been demonstrated that yeast performance is significantly impaired in environments with extremely acidic pH levels (less than 2.5). Such conditions can result in an extended lag phase, microbial growth inhibition, and even irreversible damage to the cell wall and DNA, which may render the yeast completely unviable. 59,62,63 This may explain why the F5, F16, and F27 failed to exhibit carbohydrate consumption or the production of ethanol and xylitol after 48 hours, regardless of the yeast strain used.
Yeast is particularly susceptible to stress in alkaline environments with pH levels above 8.0. In such conditions, yeast activates various adaptive mechanisms to survive, which can disrupt nutrient homeostasis, impairing the uptake and utilization of iron, copper, phosphate, and glucose. 64,65 Given that tests F2, F4, F6, F13, F15, F17, F24, F26, and F28 were conducted at initial pH values of 9.0 and 10.0, it was anticipated that the yeast cells would encounter significant challenges in either producing or consuming metabolites during these experiments.
However, the majority of tests conducted at pH and temperature levels that are typically conducive to yeast performance did not yield the anticipated results with regard to carbohydrate consumption or production of ethanol and xylitol. Given that temperature ranges of 15–40°C did not appear to be a limiting factor and that pH values between 3.0 and 6.0 generally favor yeast activity, it became essential to analyze the hydrolysate used for fermentation in detail to identify any factors that might have affected the microorganisms’ metabolism. It was determined that the concentrations of oxalate and sodium (in both associated or undissociated forms) in the medium were pivotal in impeding the fermentative performance of the three S. cerevisiae strains. These concentrations disrupt yeast metabolism and prevent the yeasts from adapting to stress conditions. The sodium concentration in the hydrolysates across different pH ranges was determined using flame atomic absorption spectrometry, and the following values were obtained: 3.78 g/L (pH 1.8), 5.83 g/L (pH 3.0), 13.53 g/L (pH 6.0), 14.28 g/L (pH 9.0), and 15.33 g/L (pH 10.2).
Oxalic acid, employed during the pretreatment process to facilitate the dissolution of hemicellulose from malt bagasse, is a dicarboxylic organic acid with dissociation constants of pKa1 = 1.25 and pKa2 = 4.27. It is widely distributed in nature and can exist in either its acid or its conjugate base form, namely oxalate, depending on the pH of the medium. The formation of oxalate salts is contingent upon the availability of metal ions and the solubility of the resulting compounds. 28
The hydrolysate utilized in the fermentation stage exhibited an approximate concentration of 45 g/L of oxalic acid, derived from a 4.5% (m/v) oxalic acid solution employed to solubilize hemicelluloses from malt bagasse. At a neutral pH, oxalic acid is fully dissociated into oxalate, which can form salts depending on the chemical species present. The pH of the hemicellulose hydrolysate was adjusted using a 2M NaOH solution. The solution undergoes dissociation, releasing sodium ions and resulting in the formation of sodium oxalate. While this is not problematic at low concentrations, the two pKa values of oxalic acid create buffering regions: one between pH 0.25–2.25 and another between pH 3.27–5.27. 28 To overcome these buffering ranges, a substantial quantity of 2M NaOH solution was required to attain the desired pH levels for the experiments, resulting in a high sodium ion concentration in the medium (13.53 g/L). For instance, the pH was adjusted to 6.0 in 800 mL of hemicellulosic hydrolysate, necessitating approximately 450 mL of 2M NaOH, which was subsequently employed in the fermentation process.
To address the issue affecting yeast metabolism, a new method for neutralizing the hydrolysate was tested with the objective of precipitating the oxalate present in the solution. The hydrolysate, prepared under optimal conditions for hemicellulose solubilization (T8—4.5% oxalic acid for 24 minutes), was adjusted to pH of 6.0 using powdered calcium hydroxide. This resulted in the precipitation of the oxalate as calcium oxalate, which was subsequently separated from the liquid fraction through vacuum filtration. Following filtration, the liquid fraction was found to contain 15.97 g/L of xylose, 4.83 g/L of glucose, and low levels of inhibitory compounds, specifically 0.002 g/L of furfural and 0.005 g/L of HMF. Furthermore, the concentration of salts in the solution was markedly reduced (1.2 mg/L of sodium and 0.7 mg/L of calcium) compared with the preceding neutralization utilizing 2M NaOH (13.53 g/L of sodium). Subsequently, the hydrolysate was supplemented and fermented using the commercial yeast S. cerevisiae PE-2, under conditions of optimal pH and temperature (6.0 at 30°C). Notwithstanding the aforementioned attempt to precipitate the oxalate, no ethanol production was observed.
It should be noted that while oxalic acid is highly effective at solubilizing hemicelluloses in malt bagasse and produces fewer inhibitory compounds from carbohydrate degradation, concentrations above 3.0% (w/v) can create challenges during subsequent neutralization and fermentation with microorganisms. A comparable result was observed by Bukhari et al. 30 wherein the pretreatment of oil palm biomass with 3.0% and 4.0% (w/v) oxalic acid led to an enhanced xylose yield but impeded fermentation by Actinobacillus succinogenes for succinic acid production. The study further demonstrated that while 1.0% (w/v) oxalic acid facilitated the production of 12.69 g/L of succinic acid, an increase in concentration to 2.0% (w/v) resulted in a significant decline in productivity, with an approximate reduction of 45%. These findings are consistent with those of Jung et al. 66 who reported that concentrations above 3.0% (w/v) of maleic acid, another organic acid, significantly inhibited the metabolism of S. cerevisiae D5A during bioethanol production. To obviate the occurrence of analogous metabolic disruptions during fermentation, it is advised that oxalic acid concentrations be constrained to a maximum concentration of 3.0%. An alternative is the use of the fungus Paecilomyces variotii FN89 for biological detoxification, which has demonstrated efficacy in degrading 68.0% of oxalic acid in wheat and corn straw pretreatments without a reduction in carbohydrate concentrations. This approach resulted in a maximum yield of 0.47 gethanol/gglucose/xylose following fermentation with S. cerevisiae XH7. 67
Production of VOCs
The extracts obtained from the liquid–liquid extraction of the supernatants collected from 33 postfermentation hemicellulosic hydrolysates were subjected to analysis using GC-MS. Additionally, the hydrolysates were subjected to analysis prior to fermentation across a range of pH values, with the objective of differentiating between the compounds present in the culture medium and those produced during fermentation. Given that the hydrolysates were supplemented with peptone and yeast extract and contained residual molecules from dead cells, certain detections were not considered. Specifically, alkane compounds derived from fatty acids, amino acids, and other substances, such as Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)-, were not considered as they were not products of the strains’ metabolism. The primary VOCs identified in the hydrolysates following fermentation are presented in Table 5.
Volatile Organic Compounds Identified in the Hydrolysate After 48 Hours of Fermentation by the PE-2, MP-P5, and JDY-01 Strains of the Yeast Saccharomyces cerevisiae
A semiquantitative analysis of the chromatograms indicated that 4-hydroxy-2-methylacetophenone is a prominent compound. This phenolic substance has been demonstrated to process notable antioxidant, antitumor, analgesic, and antiseptic properties. 67,68 The substance was identified in notable quantities in tests conducted at pH 6.0 using the three yeast strains: PE2 (tests F9, F10, F11), MP-P5 (tests F20, F21), and JDY-01 (tests F31, F32, F33). 4-Hydroxy-2-methylacetophenone is a constituent of a class of biologically active compounds designated as sesquiterpene lactones. Although it is predominantly a secondary metabolite in diverse plant species, it is also present in certain fungi and has been the subject of growing interest concerning its potential medicinal uses.
In the F26 assay using the JDY-01 strain, a peak corresponding to 44.11% of linoleic acid was observed. S. cerevisiae is capable of producing saturated and monounsaturated fatty acids, which typically contain 16 or 18 carbon atoms and up to two double bonds. 70 In a related study, You et al. 71 enhanced fatty acid production in this yeast by overexpressing acetyl-CoA carboxylase, thioesterase, and malic acid. This approach resulted in a 2.4-fold increase in fatty acid productivity, which was further enhanced by 4.7-fold when calcium nitrate was employed as a nitrogen source. Given the significant presence of linoleic acid in the F26 test, it is worth considering the JDY-01 strain for applications beyond bioethanol production.
The presence of pantolactone was detected in small amounts in most postfermentation extracts (Table 5). This compound may be present within the yeast cells or excreted during fermentation. Zea et al. 72 observed that S. cerevisiae accumulates and excretes pantolactone during the maturation of Sherry-type wines, where it plays a crucial role in forming the characteristic veil of the beverage. Pantolactone is an intermediate in the biosynthesis of pantothenic acid (vitamin B5), essential for the Krebs cycle, lipid metabolism, and yeast cell growth. 73 Moreover, it is employed extensively in the cosmetics industry as a humectant. 74
2-Phenylethanol, a primary alcohol characterized by its strong rose-like aroma, was detected in multiple tests conducted with the three yeast strains: PE2 (tests F01, F03, F10, F11), MP-P5 (tests F12, F14, F17, F18, F19, F20, F21), and JDY-01 (tests F23, F27, F29, F30, F31, F32, F33). It is widely used in the pharmaceutical, cosmetic, and food industries due to its antimicrobial and antifungal properties, making it useful as a preservative. 75 2-Phenylethanol can be produced through two pathways in yeast metabolism: the Shikimate pathway and the Ehrlich pathway. The latter pathway involves the catabolism of amino acids, primarily phenylalanine (which is present due to the addition of peptone to the hydrolysates), leading to the release of 2-Phenylethanol. 76,77 This compound was notably present in the F12 test at 21%, accompanied by ethanol and other related compounds, such as indole-3-ethanol and ethanol 2-(pentyloxy) acetate.
Similarly, 1-propanol can be synthesized from arginine, phenylalanine, glutamine, lysine, and tryptophan, as well as indole-3-ethanol from tryptophan. 78 –80 While these VOCs can also be produced via the Shikimate pathway, the glucose consumption observed in some experiments (especially in F12 and F32, where 100% and 37.5% of the hexose was consumed, respectively) did not appear to be related to the production of these compounds. This is because in the experiments in which this sugar was not consumed, the aforementioned VOCs were also detected. It can be inferred that glucose was utilized to obtain pyruvate, which was subsequently fermented into ethanol or converted into acetyl-CoA to provide the necessary energy for cell maintenance in these experiments. It is also noteworthy that higher alcohols, particularly 2-phenylethanol and indole-3-ethanol, have been proposed as signaling molecules for invasive growth in S. cerevisiae. The molecules have been shown to stimulate the expression of the FLO11 gene, which is responsible for biofilm formation, flocculation, and pseudohyphal growth by yeast when there is scarcity or difficulty in obtaining resources for growth, 81 –83 as observed in the present study.
Notwithstanding the metabolic challenges encountered by the three S. cerevisiae strains, they all successfully produced compounds of biotechnological interest. This serves to reinforce the notion that S. cerevisiae occupies a position as a “biofactory” with substantial and prospective industrial applications that remain to be fully elucidated.
Conclusion
The use of organic acids as substitutes for mineral acids has been widely studied; however, the specific effect of oxalic acid on the solubility and fermentability of the hemicellulose fraction in lignocellulosic biomasses remains inconclusive. In this study, 4.5% oxalic acid was effective in solubilizing over 99% of the hemicellulose fraction of brewer’s spent grain during pretreatment, with minimal production of inhibitory compounds such as 5-HMF, furfural, and acetic acid. However, this concentration significantly impaired the metabolism of the three S. cerevisiae strains due to oxalate dissociation during hydrolysate pH adjustment. Based on the available information and the existing literature, it may be advisable to consider utilizing a maximum of 2% of any organic acid in instances where the objective is to subject the hydrolysate to fermentation. Given the evidence, it is hypothesized that higher concentrations may prove critical for the work of the yeasts. Despite the metabolic impairment, the production of VOCs showed promising results. Notable results included 2-phenylethanol, pantolactone, and 4-hydroxy-2-methylacetophenone. Additionally, the F26 assay with the JDY-01 strain revealed a prominent peak for linoleic acid (44.1%), a long-chain fatty acid essential in the diet of most living organisms.
Footnotes
Acknowledgments
The authors sincerely thank the Federal University of Fronteira Sul (UFFS) and the Coordination of Superior Level Staff Improvement (CAPES). A.R.S. and J.M. thank the Federal University of Fronteira Sul for the scholarship. The authors are also grateful to Dr. Boris U. Stambuk (UFSC) for providing the yeast strains.
Authors’ Contributions
A.R.S.: Conceptualization, methodology, formal analysis, investigation, data curation, writing—original draft, and visualization. A.P.D.: Conceptualization, methodology, formal analysis, investigation, writing—original draft, writing—review and editing, and visualization. J.M.: Investigation. A.G.: Writing—original draft. O.F.: Formal analysis. R.M.D.: Methodology and writing—review and editing. H.T.: Writing—review and editing. G.M.M.: Writing—review and editing. S.L.A.J.: Methodology, resources, and writing—review and editing. J.P.B.: Conceptualization, methodology, supervision, project administration, resources, and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Research Promotion Program from the Federal University of Fronteira Sul (UFFS), grant # PES-2023-0521, the Brazilian Funding Agencies: Brazilian National Council for Scientific and Technological Development (CNPq-302484/2022–1, 308830/2023–7), Coordination of the Superior Level Staff Improvement (CAPES), the Research and Innovation Funding Agency of the State of Santa Catarina (FAPESC, grant number 2023TR000234), the support of the Bioprocess and Biotechnology for Food Research Center (Biofood), which is funded through the Research Support Foundation of Rio Grande do Sul (2), and Federal University of Santa Catarina (UFSC) for the financial support.