
Abstract
An efficient conversion of xylose along with glucose is needed for the sustainable production of lignocellulosic biofuels. However, the assimilation of xylose is still a major loophole for economical 2G ethanol production. Evolutionary adaptation is known to provide stability to the strains in a challenging environment. As an extension of an earlier study, the strains Kluyveromyces marxianus NIRE-K1.1 and K. marxianus NIRE-K3.1 (KmNIRE-K3.1) were subjected to secondary adaptation on optimized minimal media (MM) with the aim to enhance xylose utilization for ethanol production. Both the strains were adapted till a saturated improvement in xylose uptake, that is, 54 generations on MM medium containing xylose. Xylose utilization increased from 14.21% to 45.80% and 10.55% to 45.31%, in evolved strains KmNIRE-K1.2 and KmNIRE-K3.2, respectively. Specific xylose reductase activity also increased 2.04- and 3.36-folds in KmNIRE-K1.2 and KmNIRE-K3.2, respectively. Xylitol dehydrogenase activity was also increased by 2.82- and 1.35-folds in KmNIRE-K1.2 and KmNIRE-K3.2, respectively. Further, a decrease in redox imbalance was also observed in evolved strains, and hence, reduced in xylitol production was observed during growth and fermentation. Xylose uptake rate increased by 2.53- and 1.5-folds in KmNIRE-K1.2 and KmNIRE-K3.2, respectively with 2.20- and 6.46-folds higher ethanol concentrations, with 2.25- and 5.86-folds higher volumetric productivity, respectively. This study has demonstrated the role of evolutionary adaptation in developing robust yeast strains. KmNIRE-K1.2 and KmNIRE-K3.2 have shown enhanced ethanol production, enzyme activities, and less by-product formation such as xylitol during xylose metabolism.
Introduction
The annual availability of lignocellulosic biomass in India is about 308 million tonnes (MT) including 122 MT of rice straw and 109 MT of wheat straw. The efficient conversion of lignocellulosic biomass to bioethanol has been a major concern of researchers since the last three decades. 1,2 Although, many processes have been developed for cellulosic ethanol production but their economic viability is still challenged due to incomplete conversion of biomass to valuable products. 3 Lignocellulosic biomass contains C6 and C5 sugars including glucose and xylose as major sugar, that is, 90% of total sugar, and simultaneous conversion of both sugars into ethanol is still a loophole. 4
Further, the optimal temperatures for maximum cellulase activity and fermentation by conventional yeast Saccharomyces cerevisiae and other mesophilic yeasts are about 30°C, respectively, and have wide differences to perform in a single step. 5 In this context, simultaneous saccharification and fermentation (SSF) is one of the integrated approaches in lignocellulosic ethanol production which has various advantages, that is, high productivity of ethanol, and lower cellulase inhibition due to glucose accumulation. 6
Nowadays, thermotolerant ethanologenic yeast K. marxianus has attracted researchers to solve afore said issue due to its thermotolerance and the broad range of substrate specificity including xylose. 7 –9 It can grow up to 50°C and produce ethanol while most of the yeasts cannot survive at this temperature. 5 There are several process advantages linked with thermophilic/thermotolerant yeast for bioethanol production such as broad substrate utilization range, higher SSF rates, minimized contamination risk, lower costs of pumping and stirring, and no cooling problems, less energy requirement for mixing and product recovery. Further, various efforts have been made to engineer the K. marxianus xylose metabolic pathway including heterologous expression of xyl1 (encodes for xylose reductase [XR]) and xyl2 (encodes for xylitol dehydrogenase [XDH]) genes from Scheffersomyces stipitis or xyl11 from Orpinomyces sp. 5,10 However, the ethanol yield and productivity of these strains at higher temperatures are observed to be very low on xylose as the carbon source.
In our previous studies, we have reported the evolutionary adaptation of K. marxianus NIRE-K1 (KmNIRE-K1) and K. marxianus NIRE-K3 (KmNIRE-K3) in a rich medium with xylose as a carbon source (yeast extract, peptone, xylose, [YEPX]). 7,8 Xylose utilization in evolved strains KmNIRE-K1.1 and KmNIRE-K3.1 increased 3.42- and 3.80-folds, respectively as compared with the parent strain. Also, ethanol titer increased 5.11- and 9.72-folds in KmNIRE-K1.1 and KmNIRE-K3.1, respectively. 7,8 However, the xylose assimilation of adapted strains was not significant on minimal medium during fermentation, which is usually considered for the cost-economics of the process. In the present study, both the primary adapted cultures KmNIRE-K1.1 and KmNIRE-K3.1 were further adapted in a minimal medium containing xylose as the carbon source. The developed KmNIRE-K1.2 and KmNIRE-K3.2 strains showed enhanced xylose utilization and ethanol fermentation capability in a minimal medium containing xylose as the carbon source. The effect of secondary evolutionary adaptation on the activity of metabolic enzymes involved in xylose assimilation including XR and XDH is also explored. The changes in gene expression levels of xyl1 and xyl2 genes and co-enzyme quantification are also examined. Further, the growth and fermentation kinetics of primary and secondary adapted strains are compared.
Materials and Methods
MICROORGANISMS AND MEDIA
Primary adapted strains KmNIRE-K1.1 and KmNIRE-K3.1 were used for secondary evolutionary adaptation. 7,8 Secondary adaptation of KmNIRE-K1.1 and KmNIRE-K3.1 were performed on minimal media containing xylose as the carbon source (MMX) ([g/L]: yeast extract, 5; ammonium sulfate, 1.23; di-potassium hydrogen phosphate, 1.35; magnesium sulfate 1; sodium di-hydrogen phosphate,1.05; xylose, 20; pH, 5.5) and MMX2 ([g/L]: yeast extract, 4.81; ammonium sulfate, 1.99; di-potassium hydrogen phosphate, 1.10; magnesium sulfate, 0.95; sodium di-hydrogen phosphate, 1.05; xylose, 20; pH, 5.5), respectively. 11
EVOLUTIONARY ADAPTATION
Evolutionary adaptation in repetitive batch culture was carried out through sequential transfer in MMX medium in 100 mL of cotton-plugged Erlenmeyer flask with 20 mL of medium. The initial batch was performed by culturing loopful cells from a phytagel plate in an Erlenmeyer flask and incubated for 24 hours at 45°C in an Innova 43/43R Shaker (orbital shaker incubator) (New Brunswick, USA) at 150 rpm. Further, 1% inoculum was used for consecutive batches for subsequent growth. A 1 mL sample was withdrawn every 24 hours in each cycle for analysis of residual xylose, dry cell weight (DCW), and metabolites (ethanol and xylitol) produced during the growth. The adaptation was continued until no further improvement was observed in terms of xylose utilization and DCW.
GROWTH AND FERMENTATION CONDITION
A growth study was carried out in a bioreactor of 3 L volume (NBS BioFlo-CelliGen 115, New Brunswick, USA) under aerobic conditions with a working volume of 2 L. The temperature and pH were controlled at 45°C and 5.5, respectively, throughout the growth study. The 0.1 N HCl and 0.5 N NaOH were used as acid and base, respectively, to maintain pH. The agitation was controlled at 120 rpm and aeration was controlled at 1 L/min. The samples were collected at an interval of 2 hours and analyzed for DCW, xylose concentration, and extracellular metabolites.
Fermentation was carried out in a bioreactor of 3 L volume with a working volume of 2 L under a controlled temperature of 45°C and pH 5.5, similar to growth conditions. The agitation was maintained at 120 rpm with no aeration. Inoculum was prepared by growing the strains overnight in MMX medium at 45°C. Cells were harvested by centrifugation at 5,000x g for 10 minutes and used as inoculum for fermentation in a bioreactor with a similar composition as the growth medium except for a xylose concentration of 30 g xylose/L. The bioreactor was loaded with 2 L minimal medium and sterilized at 121°C for 15 minutes. The initial cell mass concentration was kept at ∼2.5 g cells/L. The samples were collected at intervals of 6 hours and analyzed for DCW, xylose concentration, ethanol, and byproducts. The growth and fermentation studies were carried out on a triplicate basis and sample distributions were described through the mean and standard deviation (SD).
ENZYME ASSAY
The strains were grown in a medium containing 20 g xylose/L and/or 20 g glucose/L overnight at 45°C followed by harvested at 3,000x g for 10 minutes at 4°C in a centrifuge. Protein extraction was done by using yeast buster protein extraction reagent (Novagen, Germany) and the procedure was followed according to the manufacturer’s instructions. Enzyme activity of XR and XDH were determined according to Ikeuchi et al.
12
using a spectrophotometer at A340 for KmNIRE-K1.1, KmNIRE-K1.2, KmNIRE-K3.1, and KmNIRE-K3.2 at variable temperatures ranging from 30°C to 50°C. One unit (U) of XR activity is defined as the amount of enzyme required to oxidize 1.0 μmol nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) to nicotinamide adenine dinucleotide phosphate (NADP+) for the reduction of d-xylose to xylitol in 1 minute, and one unit (U) of XDH activity is defined as the amount of enzyme required to reduce 1.0 μmol nicotinamide adenine dinucleotide (NAD+) to nicotinamide adenine dinucleotide hydrogen (NADH) for oxidation of xylitol to xylulose in 1 minute. Enzyme activities were calculated as per the formulae given by Ikeuchi et al.
12
:
where TV is total volume; V is volume of cell extract, ε is molar extinction coefficient (6.22 l mmol−1 for a path length of 1.0 cm); and CF is dilution of cell extract.
CO-ENZYME QUANTIFICATION
NADPH/NADP+ quantification
Intracellular co-enzyme concentration in yeast cells was determined after growing cells to log phase. A 2 mL sample was withdrawn and centrifuged at 2,000x g for 5 minutes and the cell pellet was washed with 20 mM phosphate buffer (pH 7.0). NADPH/NADP+ was extracted from 1 × 105 cells by using NADPH/NADP+ extraction buffers (Sigma-Aldrich, USA), and followed the manufacturer’s instructions. The co-enzyme was quantified calorimetrically at A450 in a UV-visible spectrophotometer by using NADPH/NADP+ (Sigma-Aldrich) and NADPH/NADP+ quantification kit (#MAK038, Sigma-Aldrich) by following the manufacturer’s instructions. All the results were performed in triplicate and the mean value was calculated. Furthermore, NADPH standards “0, 2, 4, 6, 8, and 10 μL” of the 10 pmol/μL were prepared to quantify the NADH at the cell extract. The total NADPH was measured at the absorbance 450 nm (A450). The ratio of NADP/NADPH in a sample may be determined by the following equation:
NADPH is an amount of NADPH in an unknown sample (pmol).
NAD+/NADH quantification
Intracellular co-enzyme concentration in yeast cells was determined after growing cells to log phase. A 2 mL sample was withdrawn and centrifuged at 2000x g for 5 minutes and the cell pellet was washed with 20 mM phosphate buffer (pH 7.0). NAD+/NADH+ was extracted from 2 × 105 cells by using NAD+/NADH+ extraction buffers (Sigma-Aldrich), and followed the manufacturer’s instructions. The co-enzyme was quantified calorimetrically at A450 in a UV-visible spectrophotometer by using NAD+/NADH (Sigma-Aldrich) and NAD+/NADH quantification kit (#MAK037 Sigma-Aldrich) by following the manufacturer’s instructions. All the results were performed in triplicate and the mean value was calculated. Furthermore, NADH standards “0, 2, 4, 6, 8, and 10 μL” of the 1 μM were prepared to quantify the NADH at the cell extract. The total NADPH was measured at the absorbance of 450 nm (A450). The ratio of NADP/NADPH in a sample may be determined by the following equation:
NADH is an amount of NADH in an unknown sample (pmol).
REAL-TIME POLYMERASE CHAIN REACTION
Real-time polymerase chain reaction (RT-PCR) was performed to determine the relative expression of xyl1 and xyl2 genes in KmNIRE-K1.1, KmNIRE-K1.2, KmNIRE-K3.1, and KmNIRE-K3.2. Total RNA was isolated by using an Aurum total RNA kit (Biorad, USA) and purified according to the manufacturer’s instructions, followed by cDNA synthesized using an iScript cDNA synthesis kit (Biorad, Canada). RT-PCR was performed in CFX connect real-time system (Biorad, India) with iTaq universal syber green supermix (Biorad, Canada). Kmatc1 encoding for actin was used as a control. The list of primers used in the gene expression analysis were reported in Supplementary Table S1 as the supplementary material.
ANALYTICAL METHODS AND KINETICS
The concentration of xylose, ethanol, xylitol, glycerol, and acetic acid were estimated using a high-pressure liquid chromatography system (Agilent Technologies, USA) with HiPlex H column with 1 mM H2SO4 as the mobile phase and flow rate of 0.7 mL/min at 57°C and detected by refractive index detector at 50°C. Yeast cell concentration was measured through DCW. Protein concentrations in cell extracts were estimated using Lowry’s method. Growth and fermentation kinetic parameters were calculated using the formulae by Bailey and Ollis.
13
STATISTICAL ANALYSIS
All the values have been expressed as mean ± SD of three independent experiments with three replicates each. The results were statistically analyzed using a one-way analysis of variance with the Holm–Sidak method using SigmaPlot (Systat Software, San Jose, CA).
Results and Discussion
EVOLUTIONARY ADAPTATION OF KMNIRE-K1.1 AND KMNIRE-K3.1
KmNIRE-K1.1 and KmNIRE-K3.1 were subjected for secondary adaptation in MMX medium and improvements were monitored in the form of DCW and sugar consumption in the broth after completion of each batch as shown in Figure 1. There was an almost twofold increase in xylose utilization after the first 10 batches in both strains. Moreover, KmNIRE-K1.1 and KmNIRE-K3.1 utilized ∼50% and 45% of total sugar, respectively after 50 batches. After 50 batches, no significant improvement was observed in the cell growth of both strains and hence, the sub-culturing was stopped after 54 cycles. There was a 3.5-fold increase in xylose utilization in adapted KmNIRE-K1.1 (Fig. 1A) and named the adapted culture as KmNIRE-K1.2. Similarly, a 4.5-fold increase was observed in xylose utilization by adapted KmNIRE-K3.1 (Fig. 1B) and named the adapted culture as KmNIRE-K3.2. DCW of KmNIRE-K1.2 increased exponentially and was about a fourfold increase, while, a slight improvement (1.17-fold) was observed in DCW in KmNIRE-K3.2.
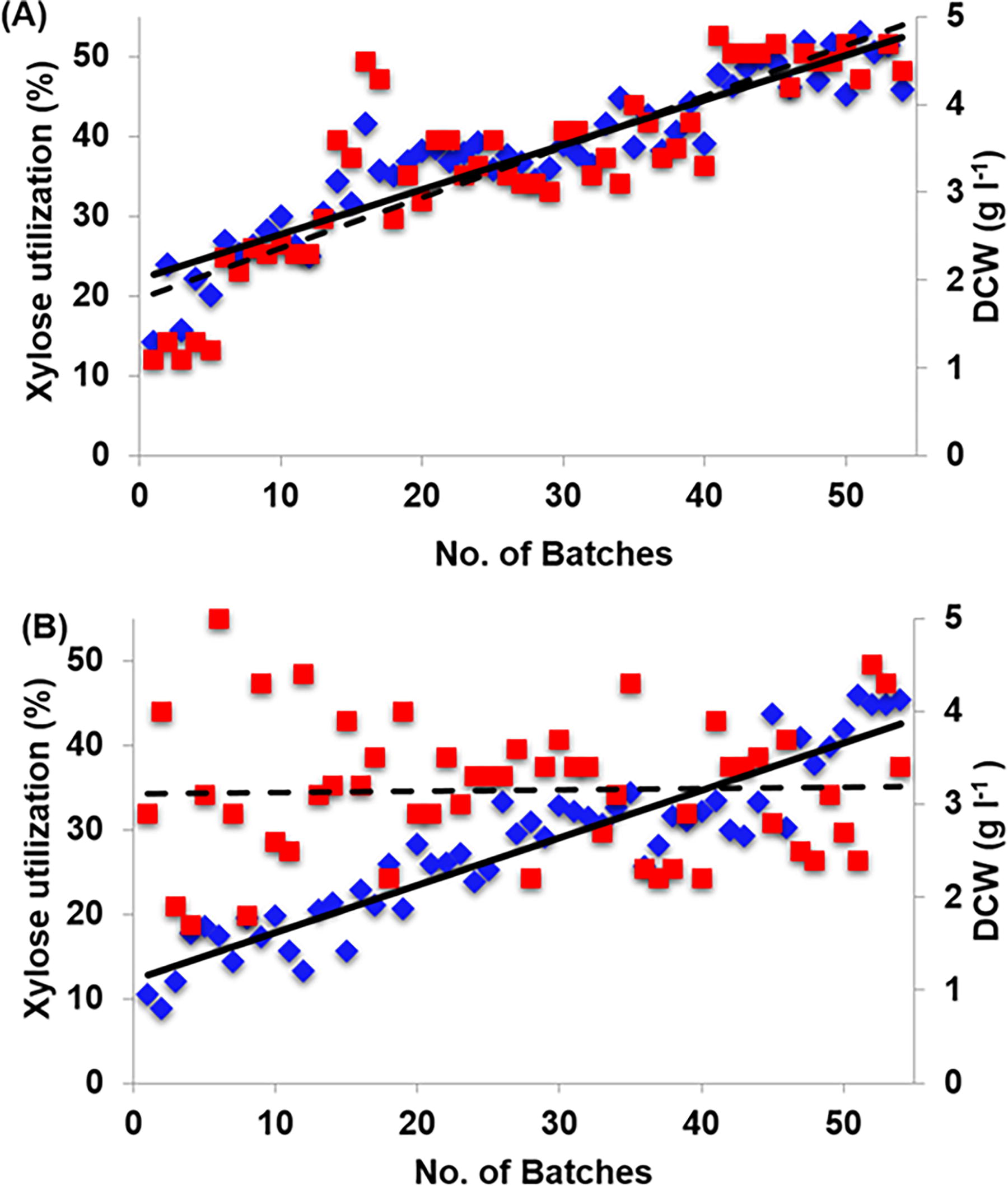
Secondary adaptation of K. marxianus strains KmNIRE-K1.1 and KmNIRE-K3.1 in MMX medium containing 20 g/L xylose. The graphs display (♦) xylose utilization and (■) dry cell weight (DCW) over time, with solid lines (—) representing the trend in xylose utilization and dashed lines (‐‐‐) representing the trend in DCW. Subfigures
As we reported in our previous studies, KmNIRE-K1.1 and KmNIRE-K3.1 were developed through primary adaptation of wild-type strains using the YEPX medium. 7,8 A significant improvement was observed in adapted cultures, but still, it was not significant in terms of xylose utilization and ethanol fermentation when grown in MMX. Hence, the primarily adapted strains were secondarily adapted in the MMX medium for further improvement. In the current study, the importance of adaptation in MM for xylose metabolism in thermotolerant yeast K. marxianus was shown. The strain improvement was proved in comparison of parental (KmNIRE-K1.1 and KmNIRE-K3.1) and evolved strains (KmNIRE-K1.2 and KmNIRE-K3.2) with metabolic enzyme (XR and XDH) activity, co-enzyme quantification (NAD+/NADH and NADPH/NADP+), gene expression (xyl1 and xyl2) and growth and fermentation. The carbon source availability impacts the evolutionary adaptation of yeast strains. In the present study, xylose was the sole carbon source similar to another study, where xylose utilizing S. cerevisiae developed through the combination of genetic engineering and adaptation. They showed improved xylose utilization with a higher specific growth rate (0.225 h−1) than that of the native recombinant strain (0.055 h−1) by using xylose as a carbon source under aerobic conditions. 14 Martín et al. 15 developed an adapted strain of genetically engineered S. cerevisiae through cultivation on a medium containing the inhibitors including aliphatic acids, furaldehydes, and phenolic compounds for 353 hours. Apart from the adaptation of culture on inhibitors, the xylose consumption also increased significantly through adaption. In another study, Mavrommati et al. 16 described an innovative adaptive laboratory evolution (ALE) strategy for the acquisition of S. cerevisiae populations more tolerant to ethanol toxicity, where populations derived after 100 generations of evolution and produced 94.77 and 98.67 g/L of ethanol for S. cerevisiae CFB and S. cerevisiae BLR, respectively, in comparison with their parental strain (84.91 and 78.09 g/L of ethanol). Liu et al. 17 developed a strain S. cerevisiae NRRL Y-50049 by using an evolutionary adaption that could detoxify furfural and hydroxymethylfurfural (HMF) while in-situ ethanol production. Similarly, Dhaliwal et al. 18 adapted thermotolerant yeast Pichia kudriavzevii on galactose medium and observed 30% enhanced ethanol production from sugarcane juice. Similarly, the present study based on secondary adaptation of both strains in MMX medium proved to be a possible approach for enhanced xylose utilization. García-Río et al. 19 evolved a cocoa strain (S. cerevisiae) in a sugary defined medium at high temperature to improve both fermentation and growth capacity. Adaptive evolution at high temperatures (40°C) enabled us to unveil the molecular basis underlying the improved phenotype by analyzing the whole genome sequence of the evolved strain. Adaptation to high-temperature conditions occurred at different genomic levels and promoted aneuploidies, segmental duplication, and single nucleotide variants in the evolved strain.
ENZYME ASSAYS OF ADAPTED STRAINS
The enzyme assays were conducted to determine the effect of adaptation on the specific activities of metabolic enzymes i.e., XR and XDH in secondary adapted strains KmNIRE-K1.2 and KmNIRE-K3.2. The enzyme activities of XR and XDH were determined in the crude enzyme extracts obtained from cells grown in glucose, xylose, and glucose/xylose mixture individually. The enzyme activity was determined at different temperatures varying from 30°C to 50°C to validate the thermo-stability of XR and XDH enzymes.
XYLOSE REDUCTASE ASSAY
XR activity for xylose grown KmNIRE-K1.2 was observed to be 1.14-, 1.03-, 1.24-, 1.27-, and 1.20-folds higher as compared with xylose grown KmNIRE-K1.1 at 30°C, 35°C, 40°C, 45°C, and 50°C, respectively. These activities were found to be 2.26-, 1.61-, 1.50-, 1.58-, and 1.28-folds higher in glucose/xylose mixture grown KmNIRE-K1.2 than that of glucose/xylose mixture grown KmNIRE-K1.1 at 30°C, 35°C, 40°C, 45°C, and 50°C, respectively (Fig. 2A). On comparing XR activities in KmNIRE-K1.2 grown in xylose and glucose/xylose mixture, it was found higher in glucose/xylose mixture at all the temperatures between 30°C and 50°C and maximum activity was observed at 50°C. XR activity in KmNIRE-K1.2 was about 1.34-fold higher in glucose/xylose mixture than only xylose at 45°C.
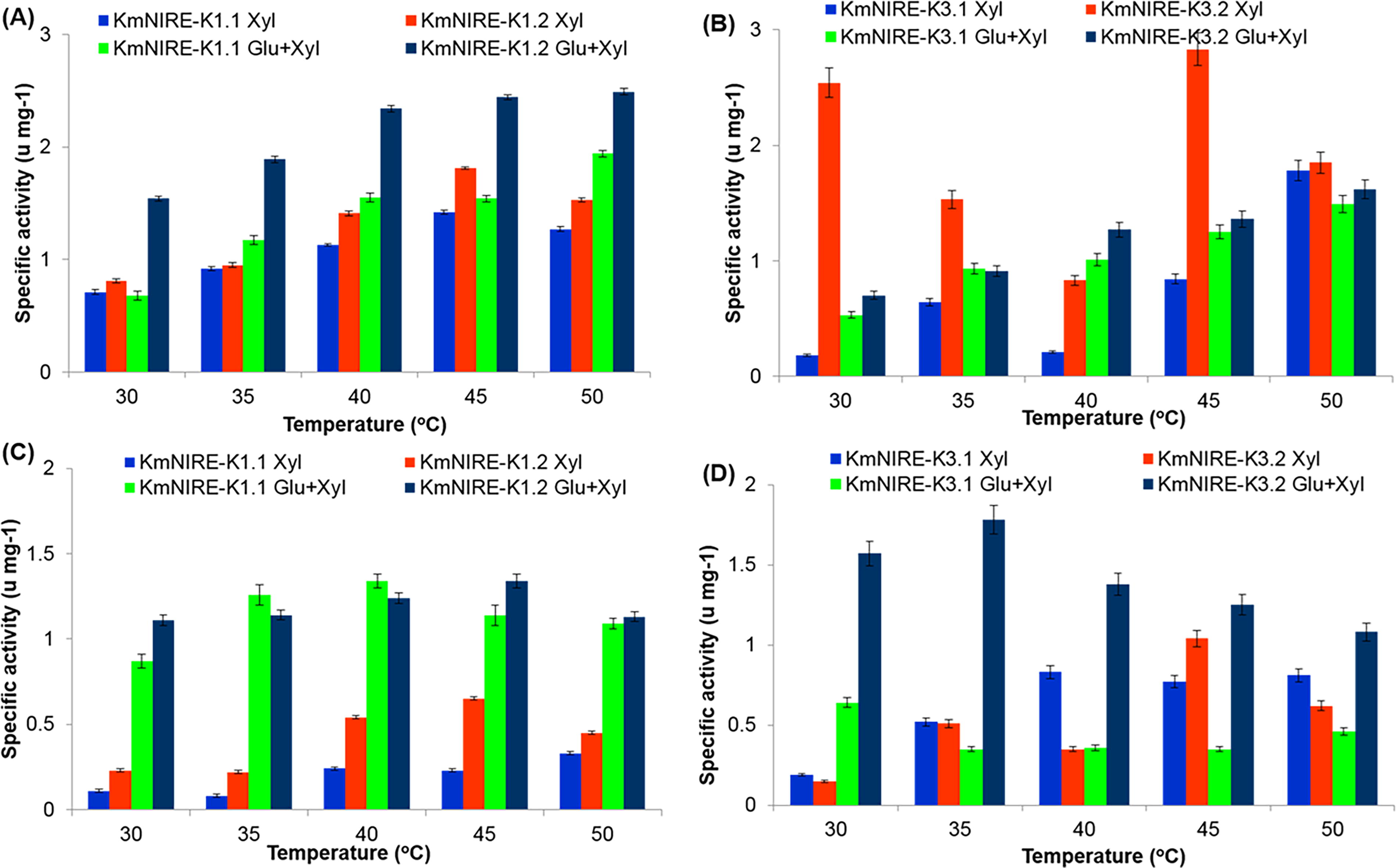
Specific activities of xylose reductase (XR) and xylitol dehydrogenase (XDH) in strains KmNIRE-K1.1, KmNIRE-K3.1, KmNIRE-K1.2, and KmNIRE-K3.2 grown at varying temperatures in media with xylose and a glucose/xylose mixture. Subfigures
Similarly, XR activities in KmNIRE-K3.2 were found to be 14.11-, 2.39-, 3.95-, 3.37-, and 1.04-folds higher than that of KmNIRE-K3.1 at 30°C, 35°C, 40°C, 45°C, and 50°C, respectively in xylose. These activities were 1.32-, 1.26-, 1.09-, and 1.08-folds higher in KmNIRE-K3.2 than that of KmNIRE-K3.1 at 30°C, 40°C, 45°C, and 50°C, respectively when grown in glucose/xylose mixture (Fig. 2B). However, no change of XR activity was observed at 35°C grown in glucose/xylose mixture using KmNIRE-K3.2 and KmNIRE-K3.1. Maximum activity was found at 50°C in glucose/xylose mixture grown KmNIRE-K3.2 (Fig. 2B). KmNIRE-K3.2 showed higher XR activity at 45°C, that is, 2.83 U/mg in xylose which is 2.04-fold higher than XR activity in glucose/xylose mixture at 45°C.
Increased XR activity on evolved strains may have been caused by the increased xylose influx after adaptation to xylose, which will increase substrate availability for the next enzyme in the metabolic pathway, that is, XDH. On comparing XR activities in both the xylose grown evolved strains, KmNIRE-K3.2 was found to be 1.53-fold higher than that of KmNIRE-K1.2 at 45°C. However, XR activity was found to be 1.79-fold higher in KmNIRE-K1.2 grown in glucose/xylose mixture than that of KmNIRE-K3.2 grown in glucose/xylose mixture at 45°C. The higher XR activity in cells grown in glucose/xylose mixture might be due to the upregulation of xyl1 gene in the presence of xylose, which enhances xylose transport. 20–21 Jeppsson et al. 22 have obtained the activity 6.07 U/mg for XR in S. cerevisiae TMB2360 grown in xylose as the sole carbon source. Similarly, Khattab et al. 23 also estimated the activity of XR in genetically modified S. cerevisae strains SK-N2, SK-N3, and SK-N4, which were found to be 1.50, 2.52, and 1.11 U/mg, respectively, when grown with xylose as the carbon source. Further, the activity of XR in thermotolerant K. marxianus YZJ088 has been reported as 0.35 U/mg 6 Similarly, Dasgupta et al. 24 have calculated an activity of XR in Kluyveromyces sp. IIPE453 as 1.036 ± 0.006 U/mg, when grown in xylose.
XDH ASSAY
XDH activities in KmNIRE-K1.2 grown in xylose as the sole carbon source were found to be 2.09-, 2.75-, 2.25-, 2.83-, 1.36-folds higher than that of KmNIRE-K1.1 at 30°C, 35°C, 40°C, 45°C, and 50°C, respectively (Fig. 2C). The maximum XDH activity in KmNIRE-K1.2 grown in xylose as the sole carbon source was observed as 0.65 U/mg at 45°C. On the contrary, XDH activities in KmNIRE-K1.2 were found as 1.28-, 1.18-, and 1.04-folds higher as compared with KmNIRE-K1.1 at 30°C, 45°C, and 50°C, respectively, grown at glucose/xylose mixture. The maximum XDH activity of 1.34 ± 0.04 U/mg was found at 45°C in KmNIRE-K1.2. On comparing XDH activities of KmNIRE-K1.2 grown in xylose and glucose/xylose mixture, the activity was higher at all the temperatures between 30°C and 50°C. XDH activity in KmNIRE-K1.2 was found to be 2.06-fold higher in glucose/xylose mixture than only xylose at 45°C.
Similarly, XDH activities in KmNIRE-K3.2 was found to be 1.04 ± 0.01 U/mg, which was 1.35-fold higher than KmNIRE-K3.1 at 45°C in xylose (Fig. 2D). On the contrary, XDH activities in KmNIRE-K3.2 were found to be 2.45-, 5.09-, 3.83-, 3.57-, and 2.35-folds higher than that of KmNIRE-K3.1 at 30°C, 35°C, 40°C, 45°C, and 50°C, respectively, in glucose/xylose mixture. However, the adapted strain showed maximum XDH activity of 1.78 ± 0.06 U/mg at 35°C. It was also observed that XDH activity KmNIRE-K3.2 was found to be 1.20-fold higher in glucose/xylose mixture than xylose at 45°C.
On comparing the activities of XDH in both the evolved strains, the activity of XDH in KmNIRE-K3.2 was found to be 1.6-fold higher than that of KmNIRE-K1.2 at 45°C in xylose. However, XDH activity in KmNIRE-K1.2 was found 1.1-fold higher than that of KmNIRE-K3.2 at 45°C in a glucose/xylose mixture. The increased XDH activities were observed after secondary adapted strains, which might be due to the combined effect of enhanced XR activity and decreased efflux of xylitol from cells. Yablochkova et al. 25 have compared the activity of XDH in different native xylose utilizing yeast strains such as Candida silvanoru VGI-II, C. tropicalis Y-456, K. marxianus Y-488, C. intermedia TabII, and C. shehatae Y-1632 and reported as 4.32, 1.16, 0.02, 3.83 and 13.95 U/mg, respectively. Goshima et al. 26 have reported XDH activity as 1.11 U/mg for K. marxianus DMB3-7, grown in xylose as the carbon source. Zhang et al. 6 have obtained 0.45 U/mg XDH activity in genetically modified K. marxianus YZJ088. Similarly, Zhang et al. 27 have estimated 0.60 ± 0.10 U/mg XDH activity in modified yeast S. cerevisiae W32N55.
CO-ENZYME QUANTIFICATION
Co-enzyme concentration was also determined during the growth of KmNIRE-K1.1, KmNIRE-K1.2, KmNIRE-K3.1, and KmNIRE-K3.2 in xylose to ensure the improvement in redox balance, which may support the improvement in xylose metabolism after adaptation. NADPH/NADP+ and NAD+/NADH ratios play an important role in xylose assimilation as the redox imbalance results in xylitol efflux. 6,28 XR oxidizes NADPH to NADP+ for conversion of xylose to xylitol and XDH reduces NAD+ to NADH to convert xylitol to xylulose, which is further phosphorylated to xylulose-5-phosphate by XK. KmNIRE-K1.2 showed improved concentrations of NAD+ and NADH, that is, 1814.09 ± 6.98 and 754.95 ± 3.72 nmol/g DCW, respectively, whereas, in KmNIRE-K1.1, these concentrations were 1052.64 ± 5.69 and 237.57 ± 5.69 nmol/g DCW, respectively (Table 1). Similarly, the concentrations of NAD+ and NADH were also increased in KmNIRE-K3.2, that is, 1142.94 ± 5.92 and 1838.7 ± 4.65 nmol/g DCW, respectively, as compared with KmNIRE-K3.1, which were 714.77 ± 4.96 and 958.4 ± 4.34 nmol/gDCW, respectively (Table 1). Furthermore, the ratio of NAD+/NADH in KmNIRE-K1.2 was 2.4, whereas in KmNIRE-K1.1, it was 4.43. Similarly, there was a value of 0.62 of NAD+/NADH ratio in KmNIRE-K3.2, whereas, in KmNIRE-K3.1, it was 0.75. The decreased ratio of NAD+/NADH showed a greater reduction of NAD+ to NADH, which might have been caused by the activity of XDH in evolved strains during xylitol to xylulose production with the help of NAD+ co-enzyme.
NAD+/NADH And NADP+/NADPH Quantifications of Primary and Secondary Adapted Strains
K. marxianus, Kluyveromyces marxianus.
Quantification of co-enzymes involved in xylose metabolism, that is, NAD+, NADH, NADPH, and NADP+, have also supported the fact of improvement with higher values of NADP+ and NADH in both the evolved strains during xylose metabolism. Moreover, the ratio of NADP+/NADPH has been observed higher in evolved strains which reveal better conversion of xylose to xylitol, which ultimately improved XR activity. Similarly, the lower value of NAD+/NADH ratio supports the fact of higher conversion of xylitol to xylulose due to better activity of XDH. Concentrations of NADPH and NADP+ in KmNIRE-K1.2 were found to be 62.71 ± 1.9 and 138.33 ± 2.69 nmol/g DCW, respectively, whereas, KmNIRE-K1.1 showed concentrations of 94.26 ± 4.21 and 165.9 ± 5.29 nmol/g DCW, respectively. Similarly, the concentrations of NADPH and NADP+ in KmNIRE-K3.2 were found to be 105.71 ± 4.95 and 165.85 ± 4.14 nmol/g DCW, respectively, which were found to be 153.12 ± 2.67 and 82.99 ± 5.17 nmol/g DCW, respectively in KmNIRE-K1.1. Moreover, the ratios of NADP+/NADPH were 1.76 and 2.21 in KmNIRE-K1.1 and KmNIRE-K1.2, respectively, and 0.54 and 1.56 in KmNIRE-K3.1 and KmNIRE-K3.2, respectively. The data show higher conversion of NADPH to NADP+ in adapted strains and resulted in higher XR activity. Moreover, the ratio of NADP+/NADPH has been increased 1.25- and 2.89-folds in KmNIRE-K1.2 and KmNIRE-K3.2 as compared with their parental strains, respectively, which may be due to higher XR activity during xylose to xylitol production. In a similar way, Zhang et al. 27 engineered K. marxianus YZJ088 with NADPH preferring XR, NADP+ preferring XDH, and NADH-dependent GLN1 and reported the increased co-enzyme concentration by 6.72-fold as compared with parent strain K. marxianus NBRC1777. Moreover, NADH/NAD+ and NADPH/NADP+ ratios were also higher, that is, 0.89 and 0.80 than K. marxianus NBRC1777, that is, 0.28 and 0.69, respectively. 27 In another study, Wei et al. 29 developed FPS1 deleted strain S. cerevisiae SR8-fps1Δ to enhance ethanol production. SR8-fps1Δ showed 10%–30% enhanced ethanol yield together with increased NADP+/NADPH ratio as compared with parent strain S. cerevisiae SR8. However, the NAD+/NADH ratio was similar to the parent strain. 29 Overall, the data depicted that the redox imbalance was reduced due to adaptation on MM containing xylose, which will decrease xylitol efflux and ultimately increase the xylose to ethanol production in evolved strains KmNIRE-K1.2 and NIRE-K3.2.
COMPARATIVE GENE EXPRESSION ANALYSIS OF PRIMARY AND SECONDARY ADAPTED STRAINS
The expression of endogenous genes xyl1 and xyl2 were analyzed using RT-PCR. The relative level of gene expression was determined between KmNIRE-K1.1 and KmNIRE-K1.2, and KmNIRE-K3.1 and KmNIRE-K3.2. The expression level of xyl1 and xyl2 in KmNIRE-K1.2 was 2.23- and 7.73-folds higher, respectively, than that of KmNIRE-K1.1 (Fig. 3A). Similarly, the expression level of xyl1 and xyl2 in KmNIRE-K3.2 were 2.53- and 6.99-folds higher, respectively, then that of KmNIRE-K3.1 (Fig. 3B). Increased expression of xyl1 and xyl2 in secondary adapted cultures KmNIRE-K1.2 and KmNIRE-K3.2 may be due to enhanced xylose uptake, which is supposed to trigger the higher metabolic enzyme activity.
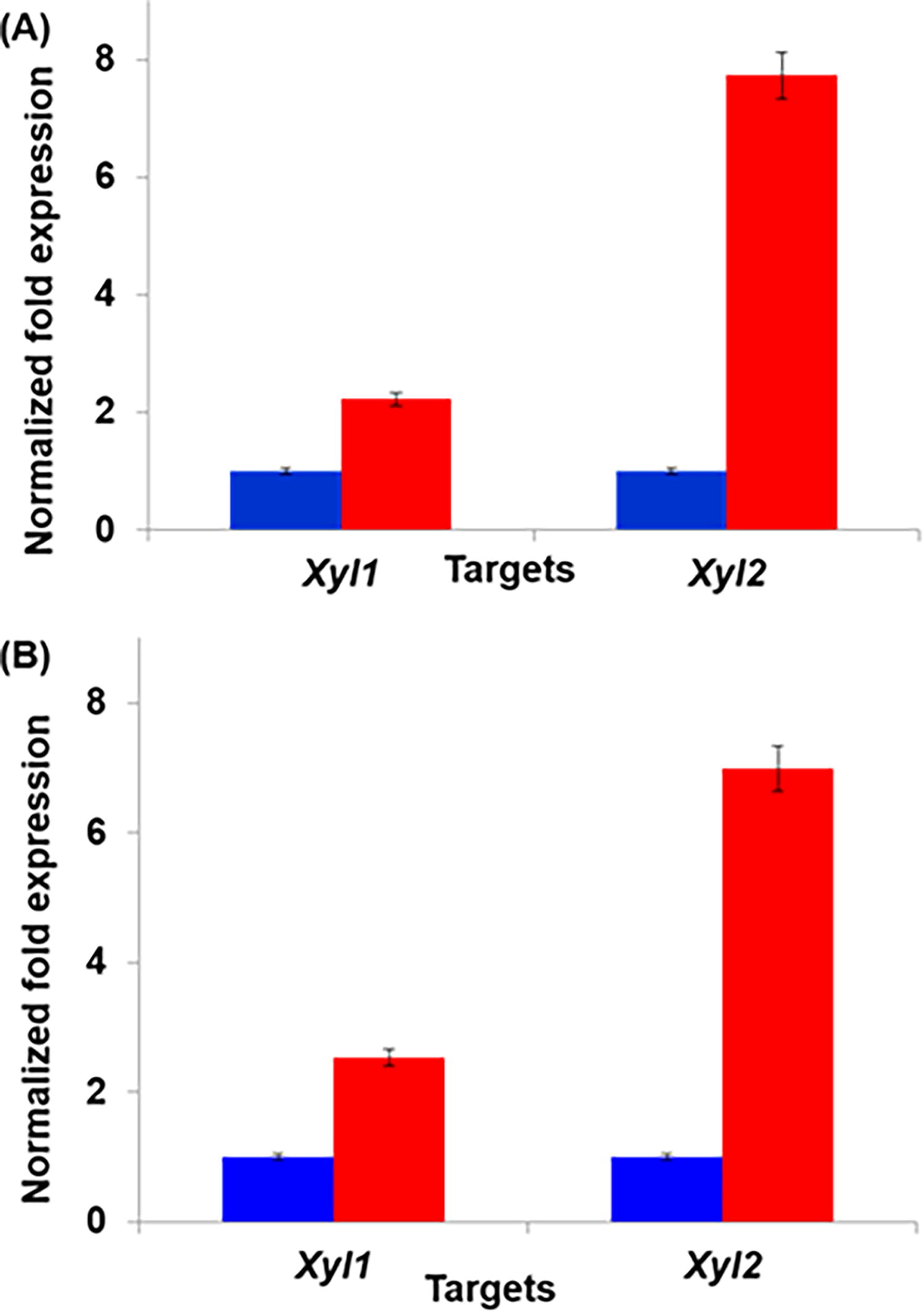
Comparative gene expression analysis of xyl1 (coding for XR) and xyl2 (coding for XDH) in adapted K. marxianus strains. Subfigure
Both the evolved strains KmNIRE-K1.2 and KmNIRE-K3.2 have shown higher gene expression of xyl1 and xyl2, which resulted in improved XR and XDH activity. The researchers have reported various studies of the relative expression of xyl1, xyl2, and other xylose metabolic genes. Li et al. 30 metabolically remodeled industrial S. cerevisiae for the purpose of utilizing both glucose and xylose with high efficiency, where heterologous gene xylA from Piromyces and homologous genes related to xylose utilization were selected to construct expression cassettes and integrated into the genome. In another study, Qi et al. 31 analyzed gene expression of xylA and xks1 of evolutionary adapted recombinant S. cerevisiae SyBE003 and reported 0.53- and 3-folds relative expression, respectively, than that of SyBE002. Biswas et al. 32 investigated the effect of different carbon sources on the expression level of the xyl2 gene of Debaryomyces hansenii and reported 7.5-fold higher expression of the xyl2 gene in xylose grown cells as compared with glucose grown cells. Similarly, Yamasaki-Yashiki et al. 33 also reported higher expression of xyl2 in Rhizomucor pusillus grown in xylose as compared with glucose. In another study, expression of xyl1 in R. pusillus NBRC 4578 was found to be higher in the cells grown in xylose and arabinose than that of grown in glucose. 34
COMPARATIVE GROWTH PROFILING OF PRIMARY AND SECONDARY ADAPTED STRAINS
Growth profiling of primary and secondary adapted cells of KmNIRE-K1 and KmNIRE-K3 were performed in a 3 L Bioflow Bioreactor containing 2 L MMX at 45°C, 5.5 pH, and 120 rpm. KmNIRE-K1.2 utilized 22.08 g xylose/L in 22 hours, whereas KmNIRE-K1.1 utilized the same amount of sugar in 28 hours (Fig. 4A). A significant difference was observed in maximum specific growth rate between the KmNIRE-K1.2 (0.22 ± 0.02 h−1) and KmNIRE-K1.1 (0.07 ± 0.007 h−1), which was about 3.14-fold higher in KmNIRE-K1.2 as compared with KmNIRE-K1.1. Similarly, KmNIRE-K3.2 and KmNIRE-K3.1 utilized about 20 g xylose/L in 16 and 22 hours, respectively (Fig. 4B). However, KmNIRE-K3.2 and KmNIRE-K3.1 showed almost similar maximum specific growth rate, that is, 0.18 ± 0.02 and 0.20 ± 0.02 h−1 (Table 2). On comparing the maximum specific growth rate of KmNIRE-K1.2 and KmNIRE-K3.2, it was found to be 1.22-fold higher in KmNIRE-K1.2 than that of KmNIRE-K3.2.
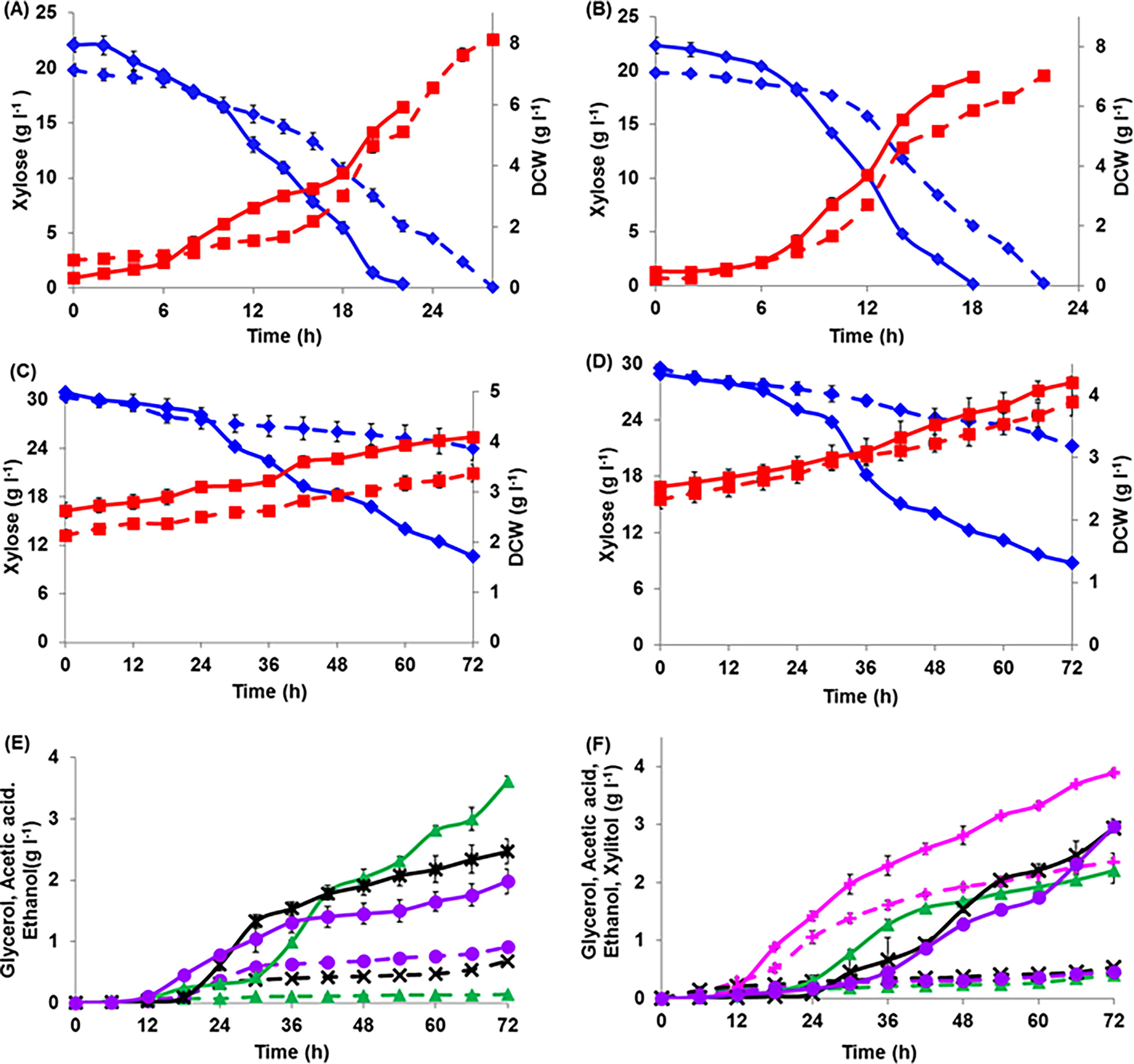
Growth and fermentation profiles of strains KmNIRE-K1.1, KmNIRE-K1.2, KmNIRE-K3.1, and KmNIRE-K3.2 on xylose as a carbon source. The growth curves (♦) for xylose, (■) DCW, (▲) glycerol, (×) acetic acid, (●) ethanol, and (+) xylitol are plotted, with solid (—) and dashed (‐‐‐) lines differentiating between strains. Subfigures
Growth and Fermentation Kinetics of KmNIRE-K1.1, KmNIRE-K1.2, KmNIRE-K3.1, and KmNIRE-K3.2
nd, not detected.
A shorter lag phase was observed in KmNIRE-K1.2 (6 hours) as compared with KmNIRE-K1.1 (10 hours). Similarly, KmNIRE-K3.2 also showed a shorter lag phase, that is, 6 hours than KmNIRE-K3.1 (8 hours). The lag phase during microbial growth usually increases due to changes in the environment, which could be reduced through adaptation in the same media. 35 ALE experimentally selects targeted phenotypes by allowing cells containing beneficial random genetic mutations to replicate more quickly in the presence of environmental stress. 36 Hence, the reduction in the lag phase after adaptation may be due to the stabilization of regulatory mechanisms. 7,37 Variation in the growth environment results in slow metabolic activity because transcription is not adapted for new changes. 37 However, after adaptation to a new environment, microorganisms show improved growth, that is, a shorter lag phase. 7,37 A shorter lag phase was also found after adaptation with various inhibitors including furfural, HMF, and acetic acid. 38 –40 Engineered strains also showed a shorter lag phase after evolutionary adaptation. 38,41,42
During growth in the MMX medium xylose uptake rate increased by 2.53-fold in KmNIRE-K1.2 (0.48 ± 0.038 g/[g·h]) as compared with KmNIRE-K1.1. Similarly, the xylose uptake rate of KmNIRE-K3.2 was found to be 1.5-fold higher than that of KmNIRE-K3.1 (Table 2). However, the cell mass yield was 1.38-fold lower in KmNIRE-K1.2 as compared to KmNIRE-K1.1. Similarly, the cell mass yield of KmNIRE-K3.2 was also slightly lower, that is, 0.30 ± 0.01 g/g as compared with KmNIRE-K3.1. The lower cell mass yield may be due to the formation of other metabolite, that is, ethanol under aerobic conditions. 43 However, the ethanol was not detected in the broth, which is supposed to escape ethanol from the vent due to air sparging. Sonderegger et al. 20 developed S. cerevisiae TMB3001C1 and TMB3001C5 through adaptation with 460 generations and reported improved maximum growth rate as 0.64 ± 0.001 and 0.119 ± 0.001 h−1, with xylose uptake rate of 0.13 ± 0.00 and 0.27 ± 0.02 g/[g·h], respectively, on 5 g xylose/L initial concentration. Similarly, Zhou et al. 44 applied a three-stage evolutionary adaptation on engineered S. cerevisiae H131-A3 and reported an improved specific growth rate of 0.12 h−1 on xylose as the sole source of carbon. Several improved features like enhanced fermentation rates, improved ethanol tolerance, and yield have also been reported through adaptation. 16 The current study shows that the evolutionary adaption in a minimal medium has proved to be a better approach for the improvement of yeast strain in context to utilize xylose present in lignocellulosic biomass.
FERMENTATION PROFILING OF ADAPTED STRAIN
Fermentation batches using primary and secondary adapted cells of KmNIRE-K1 and KmNIRE-K3 were carried out in a 3 L Bioreactor containing 2 L salt medium MMX at 45°C temperature, 5.5 pH, and 120 rpm agitation. After 72 hours of fermentation, KmNIRE-K1.2 utilized 20.26 ± 0.48 g xylose/L and produced 4.1 ± 0.09 g DCW/L from 30.92 ± 0.32 g xylose/L and 2.63 ± 0.05 g DCW/L of initial, respectively (Fig. 4C). However, KmNIRE-K1.1 utilized only 6.38 ± 0.72 g xylose/L and produced 3.37 ± 0.06 g DCW/L using initial xylose concentration of 30.38 ± 0.77 g/L and initial DCW of 2.13 ± 0.15 g/L (Fig. 4C). Simultaneously, the maximum concentration of 1.98 ± 0.2 g ethanol/L was obtained by KmNIRE-K1.2 which was 2.11-fold higher than that of KmNIRE-K1.1 (0.91 ± 0.02 g ethanol/L) (Fig. 4E). Moreover, KmNIRE-K1.2, also produced 3.61 ± 0.09 g glycerol and 2.47 ± 0.2 g acetic acid/L, whereas, KmNIRE-K1.1 produced only 0.14 ± 0.02 g glycerol/L and 0.68 ± 0.02 g acetic acid/L, respectively.
KmNIRE-K3.2 utilized 20.18 ± 0.48 g xylose/L and produced 4.2 ± 0.1 g DCW/L from 28.95 ± 0.61 g xylose/L and 2.53 ± 0.06 g DCW/L of initial, respectively, in 72 hours, whereas, KmNIRE-K3.1 utilized only 8.31 ± 0.52 g xylose/L and produced 3.89 ± 0.22 g DCW/L of 29.59 ± 0.64 g xylose/L and 2.33 ± 0.15 g DCW/L of initial (Fig. 4D). The maximum ethanol concentration produced by KmNIRE-K3.2 was found to be 2.97 ± 0.12 g ethanol/L in 72 hours, which was 6.45-fold higher than that of KmNIRE-K3.1 (0.46 ± 0.02 g ethanol/L) (Fig. 4F). Furthermore, KmNIRE-K3.2 also produced 3.89 ± 0.06 g xylitol/L, 2.2 ± 0.21 g glycerol/L, and 2.94 ± 0.02 g acetic acid/L, respectively, as by-product together with ethanol production, which were 1.65-, 5.51-, 5.44-folds higher than that of KmNIRE-K3.1, that is, xylitol (2.35 ± 0.15 g/L), glycerol (0.4 ± 0.02 g/L) and acetic acid (0.54 ± 0.04 g/L), respectively. Similarly, solid agar adapted S. stipitis CBS 6054 cells showed increased ethanol production (19.4 g ethanol/L) than that of the native strain (16.3 g/L). 45
Followed by growth and fermentation analysis showed improved xylose influx, and growth rate, and finally higher ethanol yield and productivity have been observed in evolved strains on xylose as a carbon source. The comparative growth and fermentation kinetics of all the evolved strains are presented in Table 2. The specific sugar consumption rate (qs ) in KmNIRE-K1.2 (0.085 ± 0.004 g/[g·h]) was 2.83-fold higher than that of KmNIRE-K1.1, whereas the specific product (ethanol) formation rate (qp ) was found to be twofold higher in KmNIRE-K1.2 (0.008 ± 0.0001 g/[g·h]) than that of KmNIRE-K1.1. Similarly, the specific sugar consumption rate (qs ) in KmNIRE-K3.2 (0.085 ± 0.003 g//[g·h]) was 1.7-fold higher than that of KmNIRE-K3.1, whereas, the specific product (ethanol) formation rate (qp ) was found to be 4.34-fold higher in KmNIRE-K3.2 (0.013 ± 0.001 g/[g·h]) than that of KmNIRE-K3.1. On the contrary, the ethanol yield (Yp/s ) of KmNIRE-K1.2 (0.1 ± 0.006 g/g) was nearly similar to KmNIRE-K1.1 (0.14 ± 0.02 g/g). However, ethanol yield (Yp/s ) was 2.43-fold higher in KmNIRE-K3.2 (0.14 ± 0.02 g/g) as compared with KmNIRE-K3.1, whereas, xylitol yield (Yp/s ) in KmNIRE-K3.2 (0.19 ± 0.002 g/g) was 1.47-fold lower than that of KmNIRE-K3.1. Adapted cells of Candida guilliermondii showed a threefold higher xylitol yield after adaptation in rice straw hydrolysate. 46 The volumetric substrate uptake (Qs ) and volumetric productivity (ethanol) (Qp , g/[L·h]) were 3.18- and 2.25-folds higher in KmNIRE-K1.2 (Qs = 0.28 ± 0.006 , Qp = 0.027 ± 0.002) than that of KmNIRE-K1.1, respectively. Similarly, the volumetric substrate uptake (Qs ) in KmNIRE-K3.2 (0.28 ± 0.006) was 2.34-fold higher than that of KmNIRE-K3.1. Moreover, volumetric productivities (Qp ) for ethanol and xylitol were 5.86- and 1.8-folds higher in KmNIRE-K3.2 (Qp(ethanol) = 0.041 ± 0.001; Qp(xylitol) = 0.0540 ± 0.001) than that of KmNIRE-K3.1, respectively. Furthermore, the final xylose to ethanol conversion (%) in KmNIRE-K1.2 was 19.51 ± 1.8%, whereas it was 28.84 ± 4.1% in KmNIRE-K1.1. Similarly, xylose to ethanol and xylitol conversions (%) in KmNIRE-K3.2 were 29.39 ± 0.5% and 19.19 ± 0.03%, respectively, whereas it was 11.56 ± 0.43% and 28.68 ± 0.28%, respectively, in KmNIRE-K3.1.Moreover, the cell mass yields (Yx/s ) in KmNIRE-K1.2 (0.072 ± 0.006 g/g) and KmNIRE-K3.2 (0.08 ± 0.003 g/g) were 2.63- and 2.38-folds lower than that of KmNIRE-K1.1 and KmNIRE-K3.1, respectively. Comparable results of fermentation have also been reported by a few researchers in support of the evolutionary adaptation strategy. Variants of S. stipitis were obtained with tolerance to hardwood spent sulfite liquor (HSSL) when evolutionary engineering was performed. 47 HSSL tolerance of evolved strain increased from 20% to 60% with increased substrate uptake rate (0.22 g/[L·h]) and ethanol yield (0.16 g/g). Several researchers have adopted this strategy to stabilize engineered strains. Xylose utilizing S. cerevisiae showed a 6% higher relative flux in the pentose phosphate pathway after application of long-term adaptation. 48 The developed strain produced 53 g/L of ethanol by utilizing 40 and 80 g/L of xylose and glucose, respectively, in 24 hours. In another study by Saengphing et al., 49 aimed to improve furfural tolerance in a previously isolated Spathaspora passalidarum CMUWF1 − 2, which presented thermotolerance and no detectable glucose repression, via ALE. Ultimately, the lag phase of adapted strain AF2.5 was two times shorter than that of CMUWF1 − 2 in the presence of 2.0 g/L furfural, which allowed the highest ethanol titers to be reached in a shorter period. Nielsen et al. 50 applied short-term adaptation on xylose utilizing S. cerevisiae during propagation. Adapted culture showed enhanced xylose utilization along with a 30% improved ethanol yield. Moreover, cell vitality and viability also increased by more than 10% and 20%, respectively, in adapted culture.
Evolutionary adaptation provides stability to the strains K. marxianus NIRE-K1.1 and K. marxianus NIRE-K3.1 with xylose uptake rate increased by 2.53- and 1.5-folds in KmNIRE-K1.2 and KmNIRE-K3.2, respectively with 2.20- and 6.46-folds higher ethanol concentration, with 2.25- and 5.86-folds higher volumetric productivity, respectively, for ethanol production.
Conclusions
In conclusion, the present study proved the fact that non-targeted engineering as an evolutionary adaptation improved the xylose metabolism in both the strains KmNIRE-K1.2 and KmNIRE-K3.2. By overall comparison between evolved strains, KmNIRE-K3.2 has been considered as a better strain due to higher metabolic enzyme activity, xylose uptake rate, and ethanol yield than KmNIRE-K1.2 at 45°C on xylose as the sole carbon source. However, further improvement is also suggested to reduce acetic acid and glycerol efflux during fermentation, which could further increase ethanol production. Moreover, protoplast fusion may also be attempted with potential strain to increase xylose influx and metabolism. It is also revealed that evolved strain KmNIRE-K3.2 may be considered as a potential candidate for sustainable simultaneous SSF process for second-generation bioethanol production using lignocellulosic biomass. In the future, this study will definitely directly or indirectly impact energy, promoting new industries, generating employment, and sustainable and inclusive economic growth in rural India.
Footnotes
Acknowledgments
One of the authors (N.K.S.) is very thankful to Sardar Swaran Singh National Institute of Bio-Energy, Kapurthala for providing a Senior Research Fellowship. The authors also gratefully acknowledge the Ministry of New and Renewable Energy, Govt. of India, for providing in-house support and encouragement to carry out the research activities. The authors acknowledge Research Square for the preprint (![]() ) before the publication.
) before the publication.
Authors’ Contributions
N.K.S. performed all experimentation and wrote the original draft of the article. N.K.S., S.B., R.A., A.S., and S.K. reviewed and edited the article. S.K. was involved in conceptualization, reviewing, and supervision.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Data Availability
Data will be made available on request.
Funding Statement
No funding was received for this article.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.