
Abstract
Abstract
Background:
Corticosteroids are the most efficacious anti-inflammatory drugs for asthma therapy; however, steroids are not always completedly effective for asthma. Studies have shown Mycobacterium bovis Bacille Calmette-Guérin (BCG) and other mycobacterial infections suppress airway hyperresponsiveness and inflammation in asthma. We use a murine model of Ovalbumin (OVA)-induced asthma to study whether nebulized inhalation of inactivated Mycobacterium phlei can alleviate asthmatic airway inflammation through influencing cytokine production and determine whether it can prevent and treat asthma.
Methods:
Fifth male Balb/c mice were randomly divided into four groups: normal control group (A), asthma model group (B0, B3, B4, B5), the treatment group (C0, C3, C4, C5), and prevention group (D). Mice were sensitizated and challenged with Ovalbumin to make a murine asthma model. Group C were given treatment of aerosol Mycobacterium phlei once daily after OVA challenge. Groups C3, C4, and C5 were treated for 3 days, 4 days, and 5 days, respectively. Group D inhaled the solution of inactivated Mycobacterium phlei daily before each time of OVA challenge. All the animals were killed and lung tissue and bronchoalveolar lavage fluid (BALF) were harvested. Pathological HE staining and AB-PAS staining were done to measure lung inflammation and mucus production. Total cell numbers and differential cell count in BALF were performed. Cytokines IL-4, IL-10, and IFN-γ levels in BALF were quantified by ELISA.
Results:
In groups C4, C5, and D, IL-4 production in BALF was decreased and IL-10 and IFN-γ were increased (p<0.05).The number of total inflammatory cells and the mean percentage of eosinophils and lymphocytes in the BALF of group D, group C4, and group C5 was lower than in the corresponding group B (p<0.05). Histological examination of the lungs showed airway inflammation of group D and group C5 were attenuated.
Conclusion:
The inhalation of Mycobacterium phlei can reduce airway inflammation in asthmatic mice. This ability was associated with its immunomodulatory effect on regulating IL-4, IL-10, and IFN-γ secretion. Aerosol administration of inactivated Mycobacterium phlei may be accepted as an alternative method with less risk of adverse reactions in treatment of asthma.
Introduction
Materials and Methods
Animals
Specific pathogen-free male Balb/c mice, 4–6 weeks of age, were obtained from Laboratory Animal Center of Guangxi Medical University (Nanning,Guangxi, China) and were housed under standard laboratory conditions in a pathogen-free cage. They were manitained in an air-conditioned room at 23±3°C, 55.5±10% humidity, fed a standard laboratory diet with ad libitum access to food and water.
Animal experimental protocols
Fifty male Balb/c mice were randomly categorized into four experimental groups: the normal control group (group A), the asthma model groups (group B0, B3, B4, B5), the treatment groups (group C0, C3, C4, C5), the prevention group (group D). All experiments were performed using five mice per group. For the asthma model groups, the treatment groups and the prevention group, the mice were sensitisated and challenged with Ovalbumin to make a murine asthma model. The treatment groups inhaled the atomized solution of inactivated Mycobacterium phlei after the challenge. To examine the effects of different treatment course, mice in group C3, C4, and C5 were treated for 3 days, 4 days, and 5 days, respectively. Mice in the prevention group inhaled the atomized solution of inactivated Mycobacterium phlei daily before each time's challenge.
Establishment of a murine model of asthma and the treatment
A murine model of asthma was established according to a modification of the methods proposed by North et al.(8) Mice were sensitized by intraperitoneal administration of 25 μg of Ovalbumin (OVA) (grade V, Sigma-Aldrich, St. Louis, MO, USA) emulsified in 1 mg of aluminum hydroxide (Chengdu Kelong Chemical Reagent Factory, Chengdu, China) in a total volume of 200 μL on days 0, 7, and 14. After the initial sensitization, the mice were challenged for 20 min with 2% OVA once daily by an ultrasonic nebulizer (WH-2000, Guangzhou, China) in a closed chamber from days 21 to 28. The normal control group were received only saline (instead of OVA) at both sensitization and airway challenge stages. After the challenge, mice in the treatment groups (C3, C4, and C5) inhaled the solution of inactivated Mycobacterium phlei (each Amp injection of 1.72 μg Mycobacterium phlei dissolved in 10 mL saline) using a nebulizer once daily. The Mycobacterium phlei F.U.36 injection adopts Mycobacterium phlei strain, cultured by solid and liquid, and then choose the fine cultured active ingredient. This product is low concentration with the quantity of 1 mL in an ampoule each, 1.72 μg, 4.6×106 bacterium included. Shake well before using. Do not use it if there are some clots or foreign bodies that cannot be shaken up. Group C3, C4, and C5 were treated for 3 days, 4 days, and 5 days, respectively. The prevention group received inhalation of the atomized solution of inactivated Mycobacterium phlei daily before each time challenge. The animals of group A, group B0, group C0, and group D were killed after the last OVA challenge. Specimens (lung tissue and bronchoalveolar lavage fluid) were harvested. After the treatment groups had inhale inactivated Mycobacterium phlei for 3 days, the mice of group C3 were sacrificed and those of group B3 were sacrificed correspondingly. The mice of group C4 and group B4 were sacrificed after the inhale for 4 days, group C5 and group B5 for 5 days.
Preparation bronchoalveolar lavage fluid (BALF)
The mice were sacrificed with an overdose of pentobarbital sodium (100 mg/kg body weight, i.p.), fixed in the supine position. The trachea was immediately exposed after anesthesia and carefully intubated. The catheter was secured with ligatures. Prewarmed phosphate-buffered saline (PBS) solution (at 37°C) was slowly infused into the lungs and withdrawn gentlely and Repeated three times (500 μL×3). The BALF was pooled and kept at 4°C.
Cell fractionation and differential cell count in BALF
The BAL fluid was centrifuged at 1500 rpm for 2 min at 4°C. The supernatant was kept at 20°C for determination of cytokine levels. Cell pellet was resuspended in 100 μL PBS. Total cell numbers in BALF were performed using a haemacytometer. Smears of BAL cells were prepared using a cytospin. A differential cell count was performed using Wright-Giemsa staining classified as eosinophils, neutrophils, and mononuclear cells on the basis of morphologic criteria and staining characteristics. At least 200 cells were counted per slide.
Histological examination of murine lung tissue
Lungs were isolated from the mice after BAL, fixed with 10% formalin for 24 h, and embedded in paraffin. Specimens were cut into 4-μm sections. The microsections were stained with hematoxylin-eosin (HE) and alcian blue/Periodic acid Schiff (AB-PAS). Cell infiltration and mucus production were measured. Total lung inflammation was defined semiquantitatively as the sum of the peribronchial and perivascular scores. The scoring system was as follows: 0, none; 1, mild; 2, moderate; 3, marked; and 4, severe. The adopted grading system of Goblet cell hyperplasia in the airway epithelium was as follows: 0, no goblet cells; 1, <25%; 2, 25–50%; 3, 50–75%, and 4, >75%.(9)
Measurement of cytokines in BALF with enzyme-linked immunosorbent assay (ELISA)
Interleukin (IL-4, IL-10) and IFN-γ production from BALF was measured by ELISA according to the manufacturer's instructions with a monoclonal antibody-based mouse IL-4, IL-10, and IFN-γ ELISA kit (R&D Systems, Minneapolis, MN, USA).The absorbance was measured at 450 nm by a microplate ELISA reader (Bio-Rad Laborotaries, Hercules, CA, USA).
Statistical analysis
Statistical analyses were performed using SPSS version 17.0 statistic software. Multiple groups were compared by one-way analysis of variance (ANOVA) followed by posttesting with LSD's multiple comparison of means. Histological scores were compared by the Kruskal-Wallis test. Statistical significance was set at P<0.05.
Results
Cell counts in BALF
Compared with group A, the mice of group B(B0, B3, B4, B5) caused a significant increase in the number of total inflammatory cells in the BALF (p<0.01). The mean percentage of eosinophils and lymphocytes in the BALF of group B (B0, B3, B4, B5) was significantly increased compared with group A (p<0.01). The total number of cells in the BALF was significantly less in group D, group C4, and group C5 compared with group B0, group B4, and group B5, respectively (p<0.05, p<0.01, p<0.01) (Fig. 1A). The mean percentage of eosinophils in the BALF of group D, group C4, and group C5 was significantly lower than in the group B0, group B4, and group B5, respectively (p<0.05, p<0.01, p<0.01) (Fig. 1B).
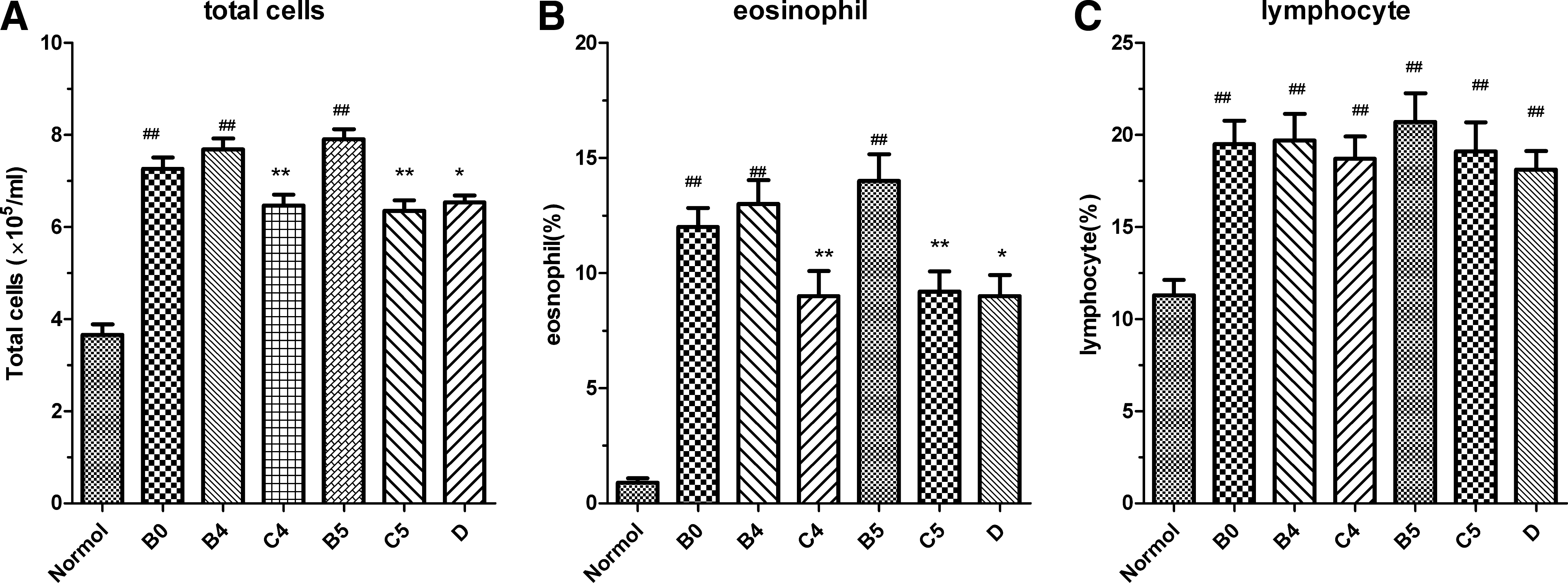
Cell counts in BALF. BALF were prepared and cell fractionation and differential cell count in BALF were performed, as described in Materials and Methods. (
Histological study of lung tissues (HE and AB/ PAS staining)
Histological examination of the lungs from the normal control group mice challenged with saline showed normal tissue,with no inflammatory cells. In the asthma model groups mice challenged with ovalbumin, histological staining showed a significant infiltration of total inflammatory cells around airways and blood vessels. The majority of the infiltrated inflammatory cells were eosinophils and lymphocytes. There were more goblet cell hyperplasia in airway epithelia and more thickened basement membranes were present than in the normal control mice. The administration of inactivated Mycobacterium phlei reduced the infiltration of inflammatory cells in the peribronchial and perivascular areas compared with the asthma model mice (Fig. 2A and B). The Inflammation scores in group B5 mice and group C5 mice were 3.6±0.89 and 2.4±1.14, respectively (n=5, p<0.05). The Inflammation scores in group B0 mice and group D mice were 3.4±0.89 and 2.6±1.14, respectively (n=5, p<0.05) (Fig. 2C). Mycobacterium phlei also reduced the hyperplasia of goblet cells. The mucus scores in group B5 mice and group C5 mice were 3.4±1.34 and 2.2±0.83, respectively (n=5, p<0.05). The mucus scores in group B0 mice and group D mice were 3.2±0.83 and 2.4±1.14, respectively (n=5, <0.05) (Fig. 2D).

Histological study of lung tissues (HE and AB-PAS staining). Lung samples were fixed with 10% formalin for 24 h, and embedded in paraffin. Specimens were cut into 4-μm sections. The microsections were stained with hematoxylin-eosin (HE) and alcian blue/Periodic acid Schiff (AB-PAS) to measure lung inflammation and mucus production. (

Cytokines (IL-4, IL-10, and IFN-γ) levels in BALF
The levels of IL-4 in BAL fluid of the asthma model groups were largely higher compared with those in the normal control group (p<0.01). The IL-10 and IFN-γ levels in the asthma model groups were lower compared with the normal control group(p<0.01, p<0.05). Inactivated Mycobacterium phlei decreased the levels of IL-4 in BALF and increased the levels of IL-10 and IFN-γ in BALF compared with the asthma model groups.The levels of IL-4 in BALF of groups C4 and C5 were significantly lower than the corresponding asthma model group (groups B4 and B5, respectively) (15.35±3.53 vs. 28.41±4.45, 14.48±4.21 vs. 30.52±3.57, respectively, p<0.01). The IFN-γ and IL-10 levels in BALF (groups C4 and C5) were significantly increased compared with the group B4 and B5, respectively (243.80±58.78 vs. 166.20±54.25, 248.40±62.53 vs. 157.20±51.74, respectively, p<0.05; 70.00±6.28 vs. 46.53±4.41, 70.72±5.40 vs. 45.47±4.14, respectively, p<0.01). IL-4 levels were significantly reduced in group D compared with the asthma model group B0 (14.60±2.89 vs. 26.81±3.59, p<0.01). IFN-γ and IL-10 productions in BALF of group D were increased compared with the asthma model group B0 (259.00±43.29 vs. 175.20±55.35,P<0.05; 71.36±6.46 vs. 47.68±4.47, p<0.01) (Fig. 3).
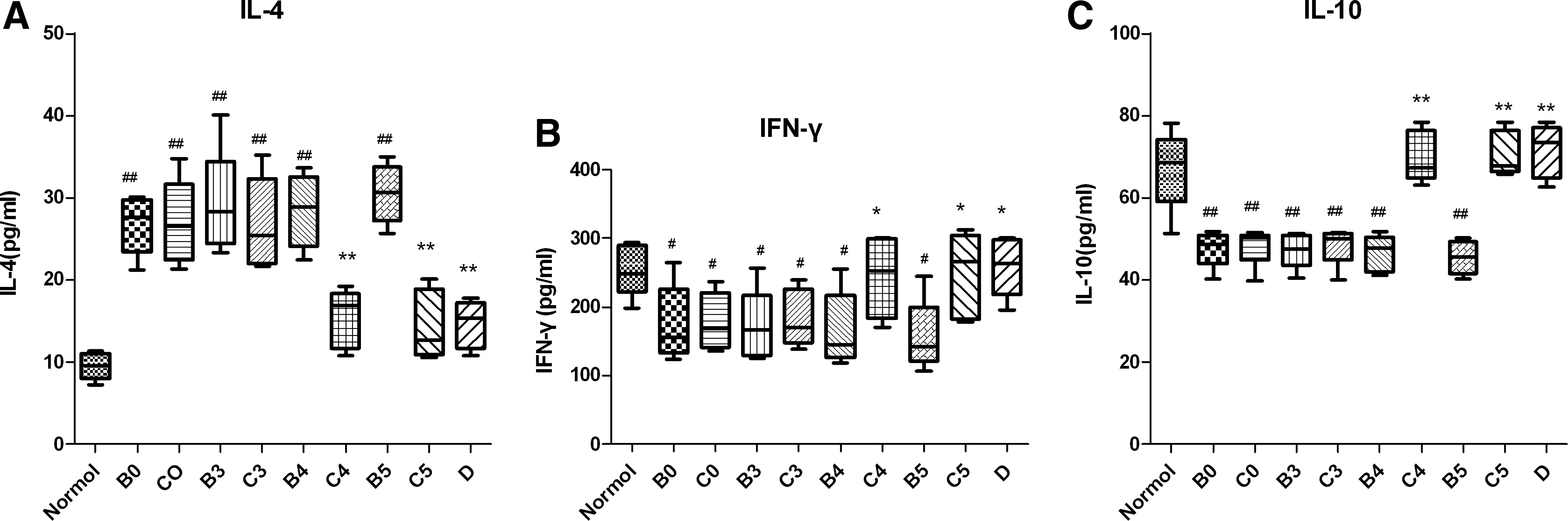
Cytokines (IL-4, IL-10, and IFN-γ). Levels in BALF. Interleukin (IL-4, IL-10) and IFN-γ production from BALF was measured by ELISA, as described in Materials and Methods. (
Discussion
Asthma is a chronic lung disease with symptoms of coughing, chest tightness, dyspnea, and wheezing, caused by reversible airflow obstruction occurring in response to various stimuli.(10) Pathophysiological performance includes variable degrees of airflow obstruction, airway hyperresponsiveness, and airway inflammation. As one of the most common chronic diseases worldwide, the prevalence of asthma has increased over the past several decades.(11) Asthma needs continuous medical treatment and thus resultes in a large burden on society. Anti-inflammatory therapy is the cornerstone of asthma therapy. As the typical anti-inflammatory drugs, corticosteroids are the most efficacious for asthma. ICS has become the first preference of asthma therapy. However,in many patients asthma is not well controlled despite the use of ICS.
The pathophysiological characteristics and inflammatory process of asthma are determined by an imbalance in the Th1/Th2 immune pattern and the the Th2-biased immune response.(12) In the response,cytokines are believed to take part in a crucial role.(13) T helper-2 (Th2) cells produce interleukin-4 (IL-4), IL-5, and IL-13 and so on. Th2 cytokine IL-4 promotes IgG1 and IgE production.(14) IL-5 enhances eosinophil accumulation,(15) and IL-13 enhance mucus secretion and airway hyperresponsiveness.(16) In contrast, IFN-γ is considered to promote Th1 cell-mediated immune responses. In case of asthma, IFN-γ inhibits the development of airway hyperreactivity in the lungs induced by Th2 responses.(17) Targeting the activation of Th2 cells or resetting the Th1/Th2 imbalance to prevent and treat asthma have become a therapeutic strategy in asthma.
There are other mechanisms involved in the pathogenesis of allergic diseases through the Th1/Th2 imbalance. It has been proposed that an immune suppression of regulatory T cells (Treg) or the induction of immunologic tolerance as factors in the development of allergies.(18) Treg play an important role in controlling Th2 biased responses both in animal models and in asthmatic humans.(19,20) IL-10, known as an important immunoregulatory cytokine, can inhibit allergic inflammation. IL-10 is mainly produced by The type 1 regulatory T(Tr1) cells.(21) It is most likely that the combination of reduced Th1 cytokine (IFN-γ) and reduced Treg cytokine (IL-10) secondary to reduced microbial exposures during childhood is responsible for the Th2-skewed immune response.
Many bacteria and their components can exert modulating effects on immune cells, including mycobacteria, LPS-(Lipopolysaccharide) and CpG-ODNs (oligonucleotides containing CpG motifs). Mycobacterium phlei being a species of Mycobacterium genus, is a nonpathogenic bacteria to human. Mycobacteria have the potential to protect against development of Th2 diseases. Some studies found that Mycobacterium bovis Bacille Calmette-Guerin (BCG) could inhibit airway inflammation and reduce airway hyperresponsiveness in animal asthmatic models.(22,23) Mutius et al.(24) showed that an increase in the tuberculosis notification rates was associated with a decrease in the prevalence of asthma. Ozdemir et al.(25) demonstrated that immunization by Mycobacterium vaccae of newborn mice prevent some of the chronic changes of airways due to asthma and suggested M. vaccae have suppressive effects on asthma. Mycobacteria is a strong inducer of Th1 type immune response, so it can reverse the Th1/Th2 imbalance in asthma. Futhermore, it is reported that BCG can induce immune-regulatory responses.(26) It remains to be explored whether some of the mycobacterial components improve Treg cells populatoin in asthma. Ahrens B et al.(27) reported that BCG-induced suppression of Th2-type allergic airway inflammation seems to be associated with an upregulation of both Treg and Th1 function. Mycobacterium phlei may have the same effect. Researchers have been committed to find less side effects and more potent therapeutic agents for the treatment of asthma. Being a nonpathogenic bacteria to human, Mycobacterium phlei may be a cheaper alternative of potential use in immunomodulatory therapy of asthma with less risk of adverse reactions.
In the present study, we found that inhaled inactivated Mycobacterium phlei attenuated the increased number of eosinophils and lymphocytes in BALF, suppressed both the infiltration of inflammatory cells and hyperplasia of goblet cells to the airways in a murine model of asthma. Our study demonstrated that inhaled inactivated Mycobacterium phlei has anti-inflammatory properties to asthmatic airway inflammation in the mouse model. The total number of cells in BALF was significantly less in in treatment groups (group C4 and C5) and in prevention group compared with the model group. The percentage of eosinophils was significantly lower in groups C4 and C5 and group D compared with group B. HE staining, AB-PAS staining revealed that peribronchial and perivascular infiltration of inflammatory cells and mucus production was significantly decreased by using inhaled inactivated Mycobacterium phlei. Importantly, in this study we obsevered that inhaled inactivated Mycobacterium phlei affect the immune response through influencing cytokine production (IL-4, IL-10, and IFN-γ). We investigated the production of IL-4 and IFN-γ in the BALF to evaluate the status of the Th1/Th2 balance. The inhibition of inflammation was associated with increased IFN-γ levels and decreased IL-4 levels in the lung. Our results illustrate that inhaled inactivated Mycobacterium phlei can alter the balance of the Th1/Th2 responses in the asthma. Furthermore, IL-10 is a potent anti-inflammatory cytokine. We observed that, in groups C4, C5, and group D, IL-10 production were increased compared with the corresponding model group. This result show that inhibitory effect on airway inflammation of inhaled inactivated Mycobacterium phlei may associated with increased IL-10 secretion. Akdis et al.(21) observed that there is an increase in the frequency of IL-4-secreting, allergen-specific T cells, and a reduction in the frequency of IL-10-secreting, allergen specific Treg cells in allergic individuals compared with nonallergic individuals. These results indicate that the balance between allergen-specific Tr1 cells and Th2 cells may be decisive in the development of allergy. Treg cells play an important role in suppressing the immune responses to allergens and preventing allergic disease. Zuany-Amorim et al.(5) showed that treatment of mice with SRP299, a killed Mycobacterium vaccae-suspension, enhances allergen-specific CD4+CD45RB(Lo) regulatory T cells and its inhibition of airway inflammation was mediated through interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β). It is possible that inhaled inactivated Mycobacterium phlei may have the potential to trigger Treg cells in prevention and treatment of asthma.
We use aerosol administration of inactivated Mycobacterium phlei through the respiratory tract, which is a safe, efficient, and noninvasive mucosal route. It is thought that modulation of local (mucosal) T cell responses may significantly impact airway inflammation and airway hyperresponsiveness.(28) Vaccine via the mucosal route would even more stimulate vigorous cell-mediated immune responses in humans.(29) γδT cells constitute 1–5% of the total blood lymphocytes but are up to 50% of the T cells in the mucosal tissues.(30) Our previous study proved γδT cells is involved in the pathogenesis of asthma.(31) Because they are most commonly known for their response to mycobacterium and their locations at mucosal sites, it is possible that γδT cells are also partly involved in the process of inhaled inactivated Mycobacterium phlei to protect against asthma.
Thus, the results of our study suggest that inhaled inactivated Mycobacterium phlei can attenuate airway inflammation in a murine model of asthma. It has both preventive effects and therapeutic effects on asthma. This ability depends its immunomodulatory mechanisms of restoring the balance of the immune system. Inactivated Mycobacterium phlei inhalation may accepted as an alternative method with less risk of adverse reactions in treatment of asthma. However, additional studies are needed to characterize the precise mechanism of it in asthma. Questions remain about how long the protective effect of vaccination on asthmatic airway inflammation lasts. Further studies are needed to define the time course and the use of inhaled immunotherapy in asthma.
Footnotes
Acknowledgments
We thank Ms. Yi Jianhua and Li Honghua for their valuable technical assistance in histopathologic examination.
Author Disclosure Statement
The authors have no conflicts of interests to declare.