
Abstract
Abstract
Background:
The long-term survival of lung cancer patients treated with conventional therapies remains poor and has changed little in decades. The need for novel approaches remains urgent. Aerosol-mediated delivery of genes has potential for the treatment of a broad spectrum of pulmonary disorders and may offer numerous advantages over invasive modes of delivery.
Methods:
The potential effects of aerosol-delivered lentiviral-based short hairpin AIMP2 lacking exon 2 (shDX2) on lung tumorigenesis were studied. Lentiviral-based shDX2 was delivered into AIMP2+/– mice through a nose-only inhalation system twice a week for 4 weeks.
Results and Conclusions:
The effects of shDX2 on lung cancer progression and the Akt1-mTOR-p70S6K signaling pathway were evaluated. Long-term repeated delivery of lentiviral-based shDX2 suppressed lung tumor progression significantly by inhibiting Akt1-related signals and decreasing both protein synthesis and angiogenesis. In vivo, the aerosol-mediated application of lentiviral-based short hairpin RNAs was successful in achieving potent and specific knockdown of the target. The collective results indicate the therapeutic potential of the repeated delivery of shDX2 for lung cancer treatment and prevention.
Introduction
The delivery of genes via aerosol holds promise for the treatment of a broad spectrum of pulmonary disorders and offers numerous advantages over more invasive modes of delivery.(5) Gene delivery into target cells is necessary in regenerative medicine, and viral vectors have been widely used as attractive gene transfer vehicles. The most commonly used viral vectors are those derived from adenoviruses and retroviruses. Adenoviral vectors have high efficiency of gene delivery, but suffer from the temporary expression of the transgene and the potent immune response directed against adenoviral structural proteins.(6) Retroviral vectors integrate into the genome, leading to long-term and stable transgene expression.(7) Among the retroviral vectors, lentiviruses have the ability of active transport of their preintegration complexes through the nucleopore, which makes them suitable for the infection of nondividing cells.(8) RNA interference (RNAi) is the pathway by which short interfering RNA (siRNA) or short hairpin RNA (shRNA) is used to inactivate the expression of target genes.(9) Whereas the siRNA-mediated knockdown of gene expression is transient in nature, viral-based shRNAs enable long-term silencing for more complex pathway analysis and screening.(10) Many proteins are too abundant or stable for phenotypic changes to be assessed in transient knockdown experiments.(11) However, with lentiviral delivery, the shRNA sequence stably integrates into the host chromosome for long-term reduction of mRNA, extending the assay time and allowing for the measurement of protein reduction.(12)
Aminoacyl-tRNA synthetases (ARSs) are enzymes that are essential for protein synthesis, by virtue of their catalysis of the ligation of specific amino acids to transfer RNAs. In higher eukaryotic systems, nine different ARSs form a macromolecular complex with three nonenzymatic factors called ARS-interacting multifunctional proteins (AIMPs).(13,14) Many of the ARSs that form the multi-ARS complex also have noncanonical functions. Like ARSs, ARS-associated factors in the multi-ARS complex play diverse roles in processes other than protein synthesis.(15) Although it has not been determined whether the complex-forming propensity of these enzymes has a relationship to their noncanonical activities, this macromolecular enzyme complex may work as molecular repository for the multifunctional housekeeping enzymes.
AIMPs are also involved in various intracellular and extracellular signaling pathways. Among them, AIMP2 plays a critical role for the assembly of the multi-ARS complex.(16) Genetic disruption of AIMP2 induces overexpression of c-myc, which causes neonatal lethality in mice through the overproliferation of alveolar epithelial cells and transforming growth factor-β.(17) In addition, AIMP2 harbored in the translation machinery acts as novel tumor suppressor through the combination of its proapoptotic activity via p53 and antiproliferative activities.(18)
AIMP2-DX2 has a deletion of exon 2 in AIMP2; the amino acid sequence corresponding to residues 46–114 is partially or completely deleted to generate a deletion variant of AIMP2 that is capable of forming a heterodimer with AIMP2. The heterodimer can inhibit the normal function of AIMP2 and promote degradation of AIMP2. This heterodimer is specifically expressed in cancer cells, in particular, in lung and liver cancer cells. An in vivo study provided evidence that AIMP2-DX2 was related to the genesis of lung and liver cancer.(19)
Therefore, the lentiviral-mediated installation of specific shRNA to suppress the expression of AIMP2-DX2 may be a rational therapeutic strategy for the treatment of cancer. Toward this end, we used lentiviral vector–based shRNA delivery via aerosol to efficiently suppress AIMP2-DX2 expression in the lungs of mice.
The role of suppressed AIMP2-DX2 in the control of cell growth is unknown. In this study, we investigated the functional significance and molecular behavior of suppressed AIMP2-DX2 in the control of cell proliferation, as well as tumorigenesis in the mouse lung.
Materials and Methods
Materials
Monoclonal antibodies against Akt1 and phospho-Akt at Ser473 were produced using a general method described elsewhere.(20) Antibodies against phospho-Akt at Thr308, PDK1, p70S6K, phospho-p70S6K (Thr389), 4E-BP1, phospho-4E-BP1 (Ser65), eIF4E, phospho-eIF4E (Ser209), vascular endothelial growth factor (VEGF), and proliferating cell nuclear antigen (PCNA) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody was obtained from BD Biotechnology (San Jose, CA). mTOR, phospho-mTOR (Ser2448), and CD31 [platelet endothelial cell adhesion molecule-1 (PECAM-1)] antibodies were obtained from Cell Signaling Technology (Beverly, MA). Monoclonal AIMP2-DX2 antibody was prepared as described previously.(19) pENTR/U6™ entry vector, pLenti6/BLOCK-iT™-DEST vector kits, and lentiviral expression plasmids were purchased from Invitrogen (Carlsbad, CA). Benzo[a]pyrene [B(a)P; 99% pure] was purchased from Sigma-Aldrich (St. Louis, MO).
Lentivirus-AIMP2-DX2 shRNA (shDX2) cloning
Five siRNA sequences targeting mouse AIMP2-DX2 mRNA were designed.(19) siRNA-1(5′-GCGGGCCACGTGC AGGACTA-3′) and siRNA-2(5′-CACGTGCAGGACTATG GGGC-3′) were superior in knocking down mAIMP2-DX2 mRNA expression in transient transfection experiments. An shRNA generated based on this siRNA sequence was cloned into the pENTR/U6 entry vector (Invitrogen). The shRNA expressing lentivirus vector (pLenti6/BLOCK-iT-DEST vector) was constructed following the manufacturer's protocol (BLOCK-iT Lentiviral RNAi Expression System; Invitrogen). Constructed AIMP2-DX2 shRNA targeting the first sequence was designated shDX2-1, and AIMP-DX2 shRNA targeting the second sequence was designated shDX2.
Preparation of lentivirus-shDX2 and titration
A total of 5×106 293T cells were seeded in 10-cm-diameter dishes 24 hr prior to transfection in Dulbecco's modified Eagle's medium (DMEM) (WelGENE, Daegu, Korea) with 10% fetal bovine serum and penicillin (100 IU/mL) in an incubator having a 5% CO2 atmosphere. A total of 12 μg of plasmid DNA was used for the transfection of one dish: 3 μg of expression plasmid and 9 μg of transfer vector plasmid viral power packaging plasmids (pLP1, pLP2, pLP/VSV-G). An equal volume of Lipofectamine reagent (Invitrogen) was added to this DNA mixture, and the complexes were incubated at room temperature for 20 min before transfection. After 24 hr, the medium (10 mL) was replaced with DMEM. The viral supernatant was collected after 48 hr. The viral concentration was determined using an HIV-1 p24ELISA kit (PerkinElmer, Boston, MA).
In vivo aerosol delivery of lentivirus-shDX2
Experiments were performed on 6-week-old male AIMP2+/– mice (n=6 per group). The breeding mice were obtained from Seoul National University (Seoul, Korea) and were kept in a laboratory animal facility with temperature and relative humidity maintained at 23±2°C and 50±20%, respectively, under a 12-hr light/dark cycle. All methods used in this study were approved by the Animal Care and Use Committee at Seoul National University (SNU-060727-6). For generating lung tumor, B(a)P (100 mg/kg of body weight; Sigma-Aldrich) was injected into 6-week-old AIMP2+/– mice once a week for 2 weeks.(19) Two weeks after the final B(a)P injection, the mice were randomized into a control group (untreated), scramble control group (lentivirus-scramble), and shDX2-treated group (lentivirus-shDX2). For aerosol delivery, AIMP2+/– mice were exposed to aerosol containing lentivirus-shDX2 solution (50 mL) that contained 40 ng/mL of lentivirus-AIMP2-DX2 shRNA. AIMP2+/– mice were allowed to inhale the aerosolized lentivirus-AIMP2-DX2 shRNA from the nebulizer for 30 min twice a week. After 1 month, the mice were sacrificed and lung samples were collected for further analysis.
Western blot analysis
Protein concentration of the homogenized lysates was measured using a Bradford kit (Bio-Rad, Hercules, CA), and equal amounts (50 μg) of protein were separated by 10–15% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Amersham Pharmacia, Cambridge, UK). The membranes were blocked for 1 hr in Tris-buffered saline–Tween 20 containing 5% skim milk, and immunoblotting was done by incubating overnight at 48°C, and then with secondary antibody conjugated to horseradish peroxidase (HRP) for 3 hr at room temperature or overnight at 48°C. After washing, the bands of interest were visualized using a LAS-3000 luminescent image analyzer (Fujifilm, Tokyo, Japan), and quantification of western blot analysis was done by using the MultiGauge version 2.02 program (Fujifilm).
Histopathological analysis and immunohistochemistry (IHC)
The lung tissues were fixed in 10% neutral buffered formalin-fixed, paraffin-embedded tissue section and were cut at a thickness of 5 μm. For histological analysis, the tissue sections were stained with hematoxylin and eosin. For IHC, the tissue sections were deparaffinized in xylene and rehydrated through alcohol gradients, then washed and incubated in 3% hydrogen peroxide (AppliChem, Darmstadt, Germany) for 30 min to quench endogenous peroxidase activity. After washing in phosphate-buffered saline (PBS), the tissue sections were incubated with 3% bovine serum albumin in PBS for 1 hr at room temperature to block the unspecific binding sites. Primary antibody was applied to the tissue sections overnight at 4°C. The following day, the tissue sections were washed and incubated with secondary HRP-conjugated antibodies (1:50) for 2 hr at room temperature. After washing, tissue sections were counterstained with Mayer′s Hematoxylin (DAKO, Carpinteria, CA) and washed with xylene. Cover slips were mounted using Permount (Fisher), and the slides were reviewed using a light microscope (Carl Zeiss, Thornwood, NY). Quantification of staining intensity of AIMP2-DX2, phospho-Akt at Thr308, p70S6K, and VEGF was performed using In Studio version 3.01 (Pixera, San Jose, CA). Staining intensity was assessed by counting the number of positive cells in randomly selected fields viewed with appropriate magnification of objective lens.
Data analysis
Quantification of western blot analysis was done using the Multi Gauge version 2.02 program (Fujifilm). All results are given as means±SE. The results were analyzed by Student's t test (Graphpad Software, San Diego, CA). p<0.05 was considered significant and p<0.01 highly significant compared with corresponding control values.
Results
Lentivirus-shDX2 inhibits alveolar/bronchiolar hyperplasia
The two shRNA-DX2 targets were aerosol-delivered to AIMP2+/– mice (Fig. 1A). shDX2-2 reduced AIMP-DX2 expression compared with shDX2-1 (Fig. 1B). shDX2-2 was referred to sbDX2 and used for further experiments (Fig. 2). IHC analysis showed that shDX2 suppressed AIMP2-DX2 expression (Fig. 1C). In addition, alveolar/bronchiolar hyperplasia incidences and grade were significantly inhibited by shDX2, as shown in Table 1 and Fig. 1E.
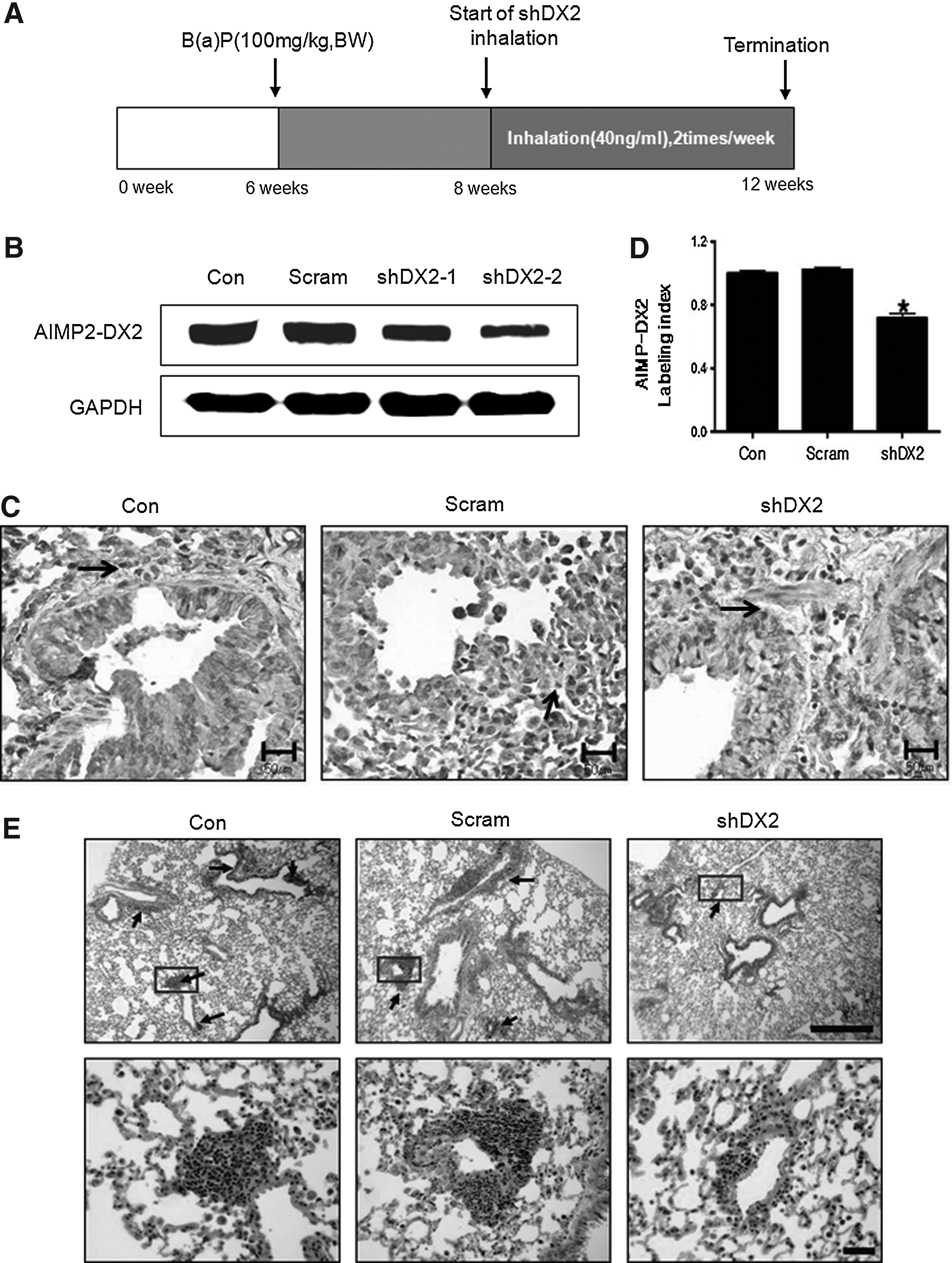
Down-regulation of AIMP2-DX2 protein expression.

Western blot and IHC analyses of Akt signals in the lungs of AIMP2+/– mice.
The AIMP2+/− male mice were given two intraperitoneal injections of B(a)P and exposed to aerosols containing lentivirus-shDX2 twice a week for a total of 4 weeks. At the end of the test period, AIMP2+/− mice were sacrificed, and then lung samples were collected. Simultaneously, the lungs from six mice were fixed in 10% neutral buffered formalin for histopathological examination. Incidence and multiplicity of lung cell hyperplasia lesions were compared.
Con, control; Scramble, Scrambled control; shDX2, shDX2-delivered group.
Moderate alveolar/bronchiolar hyperplasia.
Severe alveolar/bronchiolar hyperplasia.
Mild alveolar/bronchiolar hyperplasia.
p<0.05 compared with control.
Lentivirus-shDX2 suppresses Akt activity
To determine whether aerosol-delivered lentivirus-shDX2 would affect Akt1 activity, we measured the protein expression levels of Akt1, phospho-Akt (Thr308), phospho-Akt (Ser473), and PDK1 by Western blot. Lentivirus-shDX2 significantly decreased phospho-Akt at Thr308 and PDK1 protein levels, whereas phospho-Akt at Ser473 and total Akt protein levels remained unchanged (Fig. 2A). Densitometric analysis confirmed the western blot results (Fig. 2B). Also, IHC analysis showed that shDX2 suppressed phosphorylation of Akt at Thr308 (Fig. 2C). Densitometry analysis clearly reconfirmed the IHC result (Fig. 2D).
Lentivirus-shDX2 inhibits Akt-related signaling (mTOR/p70S6K)
To investigate the effects of aerosol-delivered lentivirus-shDX2 on the Akt-related signaling pathway, we measured the expression levels of mTOR, phospho-mTOR (Ser2448), p70S6K, and phospho-p70S6K (Thr389) by Western blot. Our results showed that lentivirus-shDX2 decreased mTOR, phospho-mTOR (Ser2448), p70S6K, and phospho-p70S6K (Thr389) protein expression (Fig. 3A). Densitometry analysis confirmed the Western blot results (Fig. 3B). IHC analysis revealed that lentivirus-shDX2 inhibited p70S6K (Fig. 3C). Densitometry analysis clearly reconfirmed the IHC result (Fig. 3D).

Western blot and IHC analyses of Akt-related proteins.
Lentivirus-shDX2 inhibits eIF4E and 4E-BP1
To examine the effects of lentivirus-shDX2 on protein translation initiation factor, we measured the expression levels of phospho-4E-BP1 (Ser65), phospho-eIF4E (Ser209), 4E-BP1, and eIF4E by western blot. Lentivirus-shDX2 slightly decreased phospho-4E-BP1 at Ser65, phospho-eIF4E at Ser209, and total 4E-BP1 and eIF4E expression (Fig. 4A). However, there was no statistical significance. Densitometry analysis confirmed the result of Western blot (Fig. 4B).
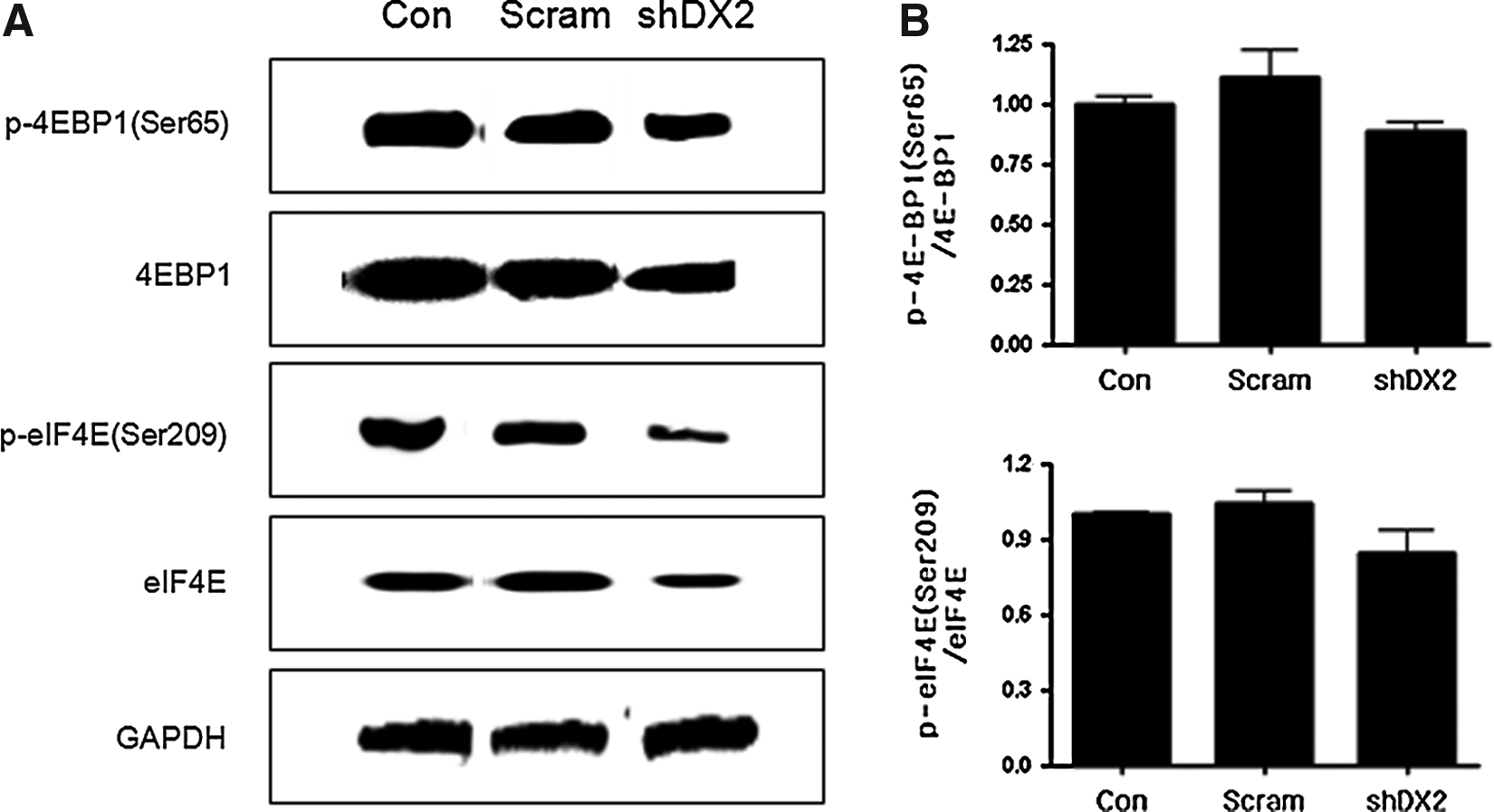
Western blot analysis of 4E-BP1, phospho-4E-BP1, eIF4E, and phospho-eIF4E.
Lentivirus-shDX2 inhibits cancer cell proliferation
VEGF is an important regulator of endothelial cell proliferation and migration.(21) Appropriately, we measured the expression level of VEGF. Lentivirus-shDX2 significantly inhibited VEGF protein expression (Fig. 5A and B). This finding was also confirmed by IHC and densitometry analyses (Fig. 5C and D). Moreover, the effect of shDX2 on cell proliferation was investigated. Decreased levels of PCNA (cell proliferation marker) and CD31 (endothelial cell marker) were detected by Western blot analyses in shDX2-delivered mice (Fig. 5E). Densitometry analyses further confirmed the result of western blot (Figs. 5F).
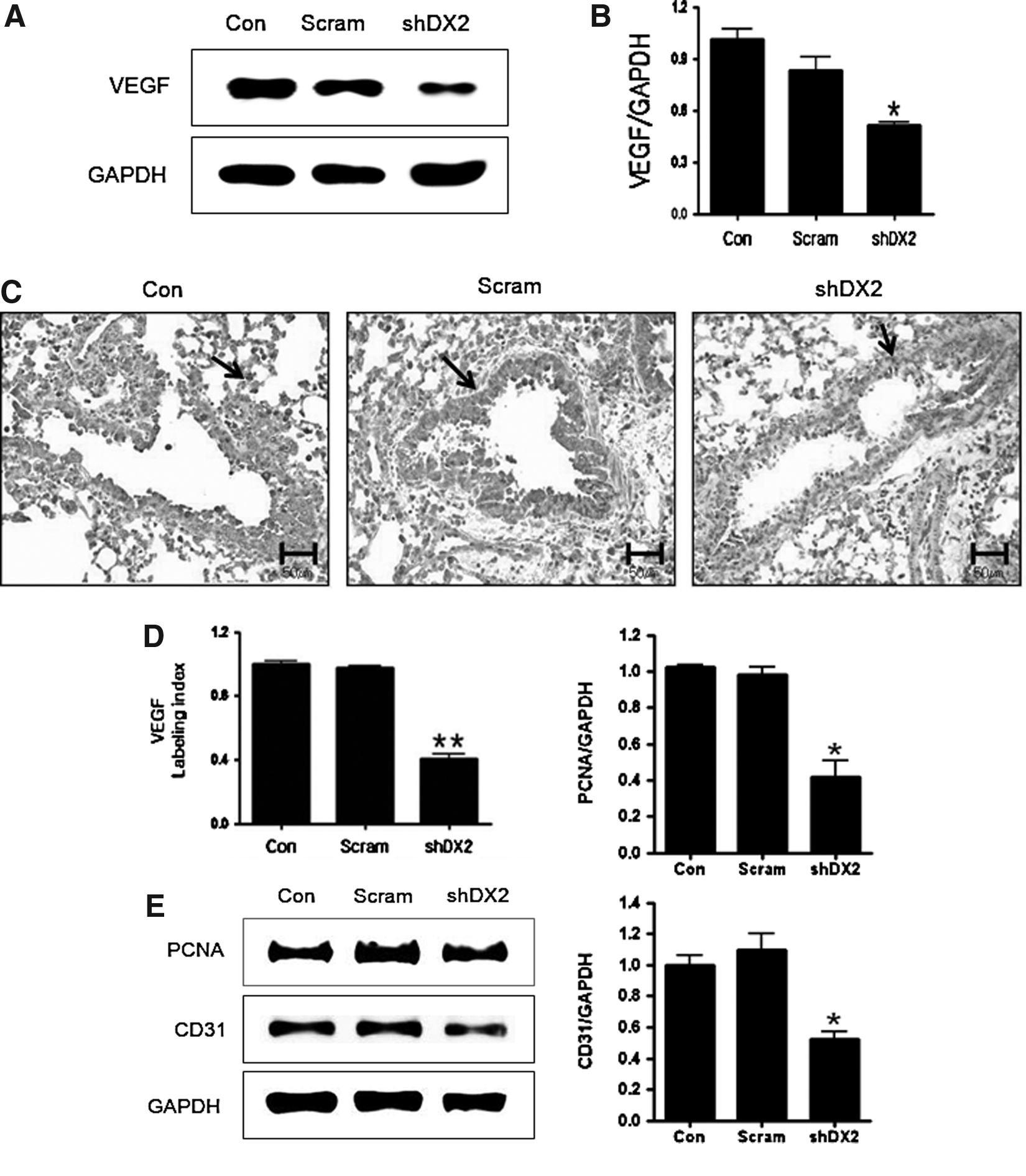
Western blot analysis of VEGF, PCNA, and CD31.
Discussion
ARSs are essential enzymes that join amino acids to transfer RNAs, thereby linking the genetic code to specific amino acids.(22) ARSs form a functional macromolecular protein complex. This complex consists of nine different ARSs and three nonenzymatic factors (AIMP1, 2, and 3).
Among them, AIMP3 was previously shown to be a haploin-sufficient tumor suppressor, which functions in the maintenance of chromosome integrity. Although AIMP2 plays a scaffolding role in the assembly of the whole complex, it also controls a major signaling pathway that is critically involved in cell death and proliferation.(17) Also, AIMP promotes cell death via the activation of p53 and apoptotic signal of tumor necrosis factor-α.(18)
The aberrant variant of AIMP2 lacking exon 2 (AIMP2-DX2) is specifically expressed in lung cancer cell lines and tissues.(23) For this reason, mice lacking AIMP2 were neonatal lethal due to lung failure resulting from overproliferation of lung epithelial cells. In addition, AIMP2 heterozygous mice with reduced expression level of AIMP2 were highly susceptible to tumorigenesis.(24) However, the inhibitory effect of AIMP2 on the development of lung tumors is still unknown. To see whether tumor development could be controlled by suppressing the expression of AIMP2-DX2, we delivered shRNA AIMP2-DX2 into AIMP2+/– mice for a month. Aerosol-delivered shDX2 significantly decreased AIMP2-DX2 protein expression (Fig. 1B, C, and D) and inhibited lung hyperplasia in the lungs of AIMP2+/– mice (Fig. 1E and Table 1). The results support the idea that the suppression of AIMP2-DX2 may play a role in suppressing lung tumorigenesis.
Akt1 kinases control essential cellular functions, including proliferation and protein translation, and have been proposed as promising targets for the treatment of angiogenesis-dependent pathologies, such as cancer.(23) Also, Akt1 is implicated in the treatment resistance of non–small-cell lung cancer (NSCLC), suggesting that Akt1 inhibition is a key factor for specific cancer cell death.(24) Our results indicate that lentivirus shDX2 inhibits the expression of phosphorylated Akt at Thr308 (Fig. 2). This observation suggests that shDX2 can inhibit Akt activity through the selective suppression of Akt phosphorylation at Thr308.
Deregulation of mTOR/p70S6K may play an important role in cancer development, as well as in many other diseases. This pathway is activated by Akt through phosphorylation.(25) Approximately 90% of NSCLC are associated with activation of the Akt-mTOR-p70S6K pathway, and the activation of this pathway promotes cellular survival and proliferation.(26) Overexpression of p70S6K also has been found in many tumor cells and results in increased tumor size in vivo, whereas inhibition of p70S6K prevents cell proliferation.(27) Our results indicate that shDX2 suppresses phosphorylated mTOR at Ser2448 and p70S6K phosphorylation at Thr389, suggesting that shDX2 can regulate cancer growth and proliferation through Akt-related signals (Fig. 3A and B).
4E-BP1 is another mTOR substrate that inhibits the function of eIF4E. 4E-BP1 binds directly to eIF4E to inhibit complex assembly for translation initiation.(28) Overexpression of eIF4E in experimental models dramatically alters cellular morphology, enhances proliferation, and induces cellular transformation, tumorigenesis, and metastasis. Both eIF4E and 4E-BP1 are frequently overexpressed in lung cancer.(29,30–31) This suggests that inhibition of key factors of protein translation initiation complex through blockade of the mTOR-p70S6K-eIF4E pathway by shDX2 may reduce protein synthesis, leading to the inhibition of lung cancer cell proliferation as well as progression.
mTOR also regulates other cellular processes, including RNA polymerases, protein phosphatases, and VEGF signaling.(21) One potent marker for angiogenesis is VEGF, which induces endothelial cell proliferation and migration. The present observation that shDX2 decreased the expression levels of VEGF, PCNA (cell proliferation marker), and CD31 (endothelial cell marker) supports the suggestion that shDX2 may regulate endothelial cell proliferation (Fig. 5).
In summary, the collective results indicate that suppression of AIMP2-DX2 may be effective in regulating cell proliferation through the inhibition of the Akt1-mTOR-p70S6K pathway. Thus, AIMP2-DX2 has potential as a novel therapeutic target in the treatment and prevention of cancer.
Footnotes
Acknowledgments
This work was supported by National Research Foundation grants (NRF-2012-0001116 and 2012M3A9C4048819) from the Ministry of Education, Science and Technology (MEST). M.H. Cho was also partially supported by the Research Institute for Veterinary Science, Seoul National University.
Author Disclosure Statement
The authors declare that no conflicts of interest exist.