
Abstract
Abstract
Background:
The expression of inflammatory cytokines in lung tissue plays an important role in immune function of the lung. In this study, we tested whether aerosol delivery of the gene of interferon-γ (IFNγ) could affect inflammatory cytokine expression in mouse lung.
Methods:
Murine IFNγ-expressing plasmids (pcDNA-IFNγ) complexed with polyethylenimine (PEI) (PEI/pcDNA-IFNγ) were constructed, and their transfection efficiency was assessed in vivo using real-time quantitative RT-PCR and enzyme-linked immunosorbent assay. After aerosol administration of the plasmid complexes and confirmation of the IFNγ plasmid location in lung tissue, we measured mRNA levels of the inflammatory cytokines interleukin-1 (IL-1), IL-6, IL-10, tumor necrosis factor-α (TNF-α), and granulocyte-macrophage colony-stimulating factor (GM-CSF) on days 1 to 7 in mouse lung tissues using real-time RT-PCR.
Results:
IFNγ mRNA expression in mouse lung was significantly increased 24 hr after a single aerosol administration of PEI/pcDNA-IFNγ and gradually decreased over the next 5 days, whereas the mRNA expressions of IL-1, IL-6, and GM-CSF were markedly decreased, but not those of IL-10 and TNF-α.
Conclusions:
PEI/IFNγ gene therapy delivered by aerosol has immune-regulating potential by suppressing lung cytokine mRNA expression, and therefore may alleviate lung disease.
Introduction
D
Aerosol delivery of gene therapy to the lung has vast potential for the treatment of pulmonary diseases. Gene complexes delivered by aerosol have been shown to have broad therapeutic activities.(7–9) In this study, we delivered branched polyethylenimine (PEI)/pcDNA-IFNγ plasmid complexes via aerosol to test the expression of IFNγ in murine lung tissues and to observe changes in mRNA expression of several inflammatory cytokines.
Materials and Methods
Animals
Female Bacl/C mice (5–7 weeks old) were purchased from Shenyang Pharmaceutical University, China. Housing and feeding of the mice and experimented procedures complied with Chinese regulations of the Administration of Affairs concerning experimental animals. Mice were housed five in a cage and kept on a 12-hr light/dark cycle.
Reagents
Branched 25-kDa PEI was purchased from Sigma-Aldrich Chemical GmbH (Steinheim, Germany). A PEI stock solution was prepared at a concentration of 4.3 mg/mL (0.1 M in nitrogen) in 150 mM NaCl. The solution was neutralized with HCl to pH 7.2 and filtered (pore size 0.2 μm). Escherichia coli DH5α cells were obtained from TaKaRa Biotechnology (Dalian, China) and grown in Luria-Bertani medium at 37°C. Endonucleases EcoRI and BamHI, T4 DNA ligase, and Ex Taq were purchased from TaKaRa Biotechnology.
Plasmid construction
Total mRNA was extracted from Bacl/C mouse lung using TRIzol reagent in accordance with the instructions from the provider (Invitrogen). Reverse transcription (RT) of the mRNA to generate the cDNA pool was performed with oligo-(dT) primers. The mouse IFNγ gene was amplified via polymerase chain reaction (PCR) with forward primer (FP) 5’-CATGGAATTCATGAACGCTACACACT GC-3’ and reverse primer (RP) 5’-CATGGGATCCTCAGCAGCGACTCCTTTT-3’. The PCR product and eukaryotic expressing plasmid pcDNA3.1(–) (Invitrogen) were cut with the endonucleases EcoRI and BamHI and directly cloned to create the recombinant vector pcDNA-IFNγ. The recombinants were screened using PCR and identified via restriction endonuclease digestion analysis. The positive clones were identified by gene sequencing.(10)
The plasmids were replicated in cells of the high-copy DH5α strain of E. coli and isolated through alkaline lysis, followed by purification in accordance with instructions in the PureYield Plasmid Midiprep System kit (Promega). The purity of the plasmids was verified by resolution in an agarose gel, and absorbance readings at 260 and 280 nm were measured with a BioPhotometer (Eppendorf, Hamburg, Germany) to ensure that their ratio fell within the acceptable range.
Design of PEI/pcDNA-IFNγ complexes
An optimized method for the design of modified PEI was performed as previously reported.(11) In brief, the complex was prepared at a ratio of 10:1 nM for PEI nitrogen:DNA phosphate. Two milligrams of control plasmids pcDNA-IFNγ/pcDNA3.1(–) dissolved in water was added slowly into the PEI solution and vortexed. There was 2 mg of plasmid for one nebulizer dose in a chamber. The mixed solution was incubated at room temperature for 15 min before being used.
In vivo transfection with PEI/pcDNA-IFNγ via aerosol
PEI/pcDNA-IFNγ or PEI/pcDNA3.1(–) was aerosolized using the PARI TurboBOY N nebulizer (PARI GmbH, Starnberg, Germany). Aerosol was passed through the sealed plastic cage (12×10×22 cm) that housed five 5- to 7-week-old mice. The control groups included five untreated mice and five mice treated with PEI/pcDNA3.1 only. Mice were housed in the nebulization chamber and administered a single dose of 2 mg of PEI/pcDNA3.1 (pcDNA) or aerosol PEI/pcDNA-IFNγ (pcDNA-IFN) in aerosol form, or they were not aerosolized (control).
To determine the persistence of IFNγ expression and the changes in expression of the other cytokines, mice were anesthetized and euthanized at different time points after a single aerosol delivery, and lungs were harvested and homogenized. There were five mice in each group per time point. Total RNA was extracted using TRIzol reagent, based on the instructions from the provider (Invitrogen). The mRNA levels of interleukin-1 (IL-1), IL-6, tumor necrosis factor-α (TNF-α), IFNγ, and granulocyte-macrophage colony-stimulating factor (GM-CSF) were determined by real-time quantitative RT-PCR (qRT-PCR) as described below.
In situ hybridization
To detect the localization of pcDNA-IFNγ following transfection, the in situ hybridization (ISH) method was used. In brief, the IFNγ PCR product (FP, 5’-ATCAGGCCATCAGCAACAAC-3’; RP, 5’-ATCAGCAGCGACTCCTTT TC-3’) was used as a probe and was labeled with biotin. Biotin-11-dUTP was incorporated into the DNA using a biotin random prime DNA labeling kit (Beyotime Institute Biotechnology, Jiangsu, China). Five-micrometer tissue sections on dimethyldichlorosilane-coated glass slides were rapidly dewaxed, cleared, and air-dried. After digestion with proteinase K, 20 ng of biotinylated probes was dropped onto the slides and distributed with a coverslip. The preparations were denatured with the DNA probe for 10 min in a moisture chamber at 85° in a water bath. The moisture chamber was transferred to a 37° water bath for hybridization. The coverslip was removed after overnight, and the slides were washed for 2×15 min in 50% formamide/2×standard saline citrate (SSC), 2×5 min in 2×SSC, and 3×5 min in 0.1% Tween-20/2×SSC in sequence, and then were rinsed in PBS for 3 min (all solutions pH 7.0). The slides were covered with streptavidin–horseradish peroxidase complex (diluted 1:200) and incubated for 30 min. After a rinsing with PBS, the slides were stained with DAB (0.5 mg of diaminobenzidine/5 mL 30% H2O2/1 mL PBS). After using neutral red-stained preparations, dark-brown hybridization signals were detected.
qRT-PCR
qRT-PCR was performed using a sequence detection system (ABI PRISM 7000; Applied Biosystems, Life Technologies, Grand Island, NY), and all reaction components were purchased from the same source (SYBR PrimeScript RT-PCR Kit obtained from TaKaRa Biotechnology). The standard experiments were carried out based on the instructions of the provider. All reactions were performed in triplicate. The threshold for the cycle threshold analysis of all samples was set automatically by the ABI PRISM 7000 software. The mRNA expressions of the indicated genes were normalized relative to β-actin mRNA. The primers were designed as previously reported(12) and are shown in Table 1.
Enzyme-linked immunosorbent assay
The concentrations of IFNγ in BALF were quantified by enzyme-linked immunosorbent assay (R&D Systems, BD Biosciences, San Jose, CA, and eBiosciences, San Diego, CA). Two milligrams of PEI/pcDNA-IFNγ or PEI/pcDNA3.1(–) complexes was nebulized in each group with five mice. One half milliliter of BALF was harvested from each mouse after 24 hr.
Statistical analyses
All data are presented as the means±standard error of the mean. The statistical comparison between groups was carried out using Student's t test. A p value of <0.05 was considered statistically significant.
Results
Regional lung location of IFNγ plasmid after aerosol PEI/pcDNA-IFNγ
We nebulized PEI/pcDNA-IFNγ and PEI/pcDNA3.1(–) to the lungs of mice using the above method. Twenty-four hours after aerosol application, we detected the IFNγ plasmid location by the ISH method. At low magnification, hybridization signals of IFNγ plasmid were detectable distributed in the alveolar (Fig. 1B) but not in the aerosol PEI/pcDNA3.1(–) lungs (Fig. 1A). At high magnification, hybridization signals of IFNγ plasmid were found in the alveolar macrophage (Fig. 1C) and alveolar epithelium (Fig. 1D) in the lungs of mice following PEI/pcDNA-IFNγ aerosol application.
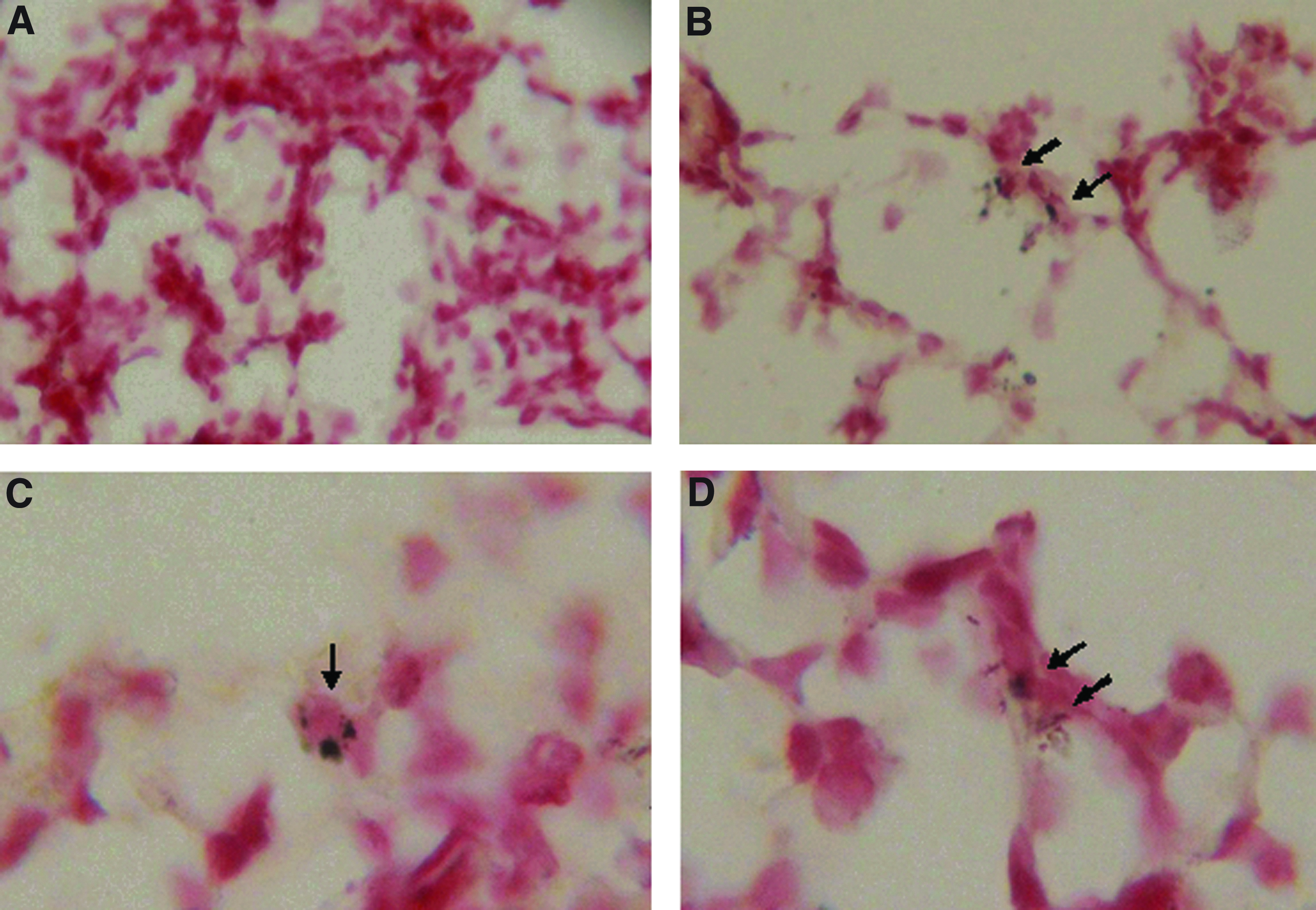
Lung tissue location of IFNγ plasmid after transfection. Two milligrams of control plasmids pcDNA3.1(–) or pcDNA-IFNγ was nebulized into a plastic box that housed the mice. Twenty-four hours after nebulization, mice were killed and paraffin sections of the lungs were detected for location of pcDNA plasmid using the ISH method.
Expression of IFNγ mRNA and protein after aerosol treatment with PEI/pcDNA-IFNγ plasmid complexes
To assess the transfection efficiency of the aerosolized gene therapy treatment in murine lung tissue, the expression level of IFNγ mRNA was determined using qRT-PCR 24 hr after transfection of PEI/pcDNA-IFNγ complexes. In the mice treated with aerosolized PEI/pcDNA-IFNγ, IFNγ mRNA levels were significantly increased (60-fold) compared with those of the nonaerosolized mice or those treated with PEI/pcDNA3.1 (p<0.05 for both; n=5 for each treatment group; Fig. 2A). There were no significant differences in IFNγ mRNA levels between the PEI/pcDNA3.1 and the nonaerosolized groups.
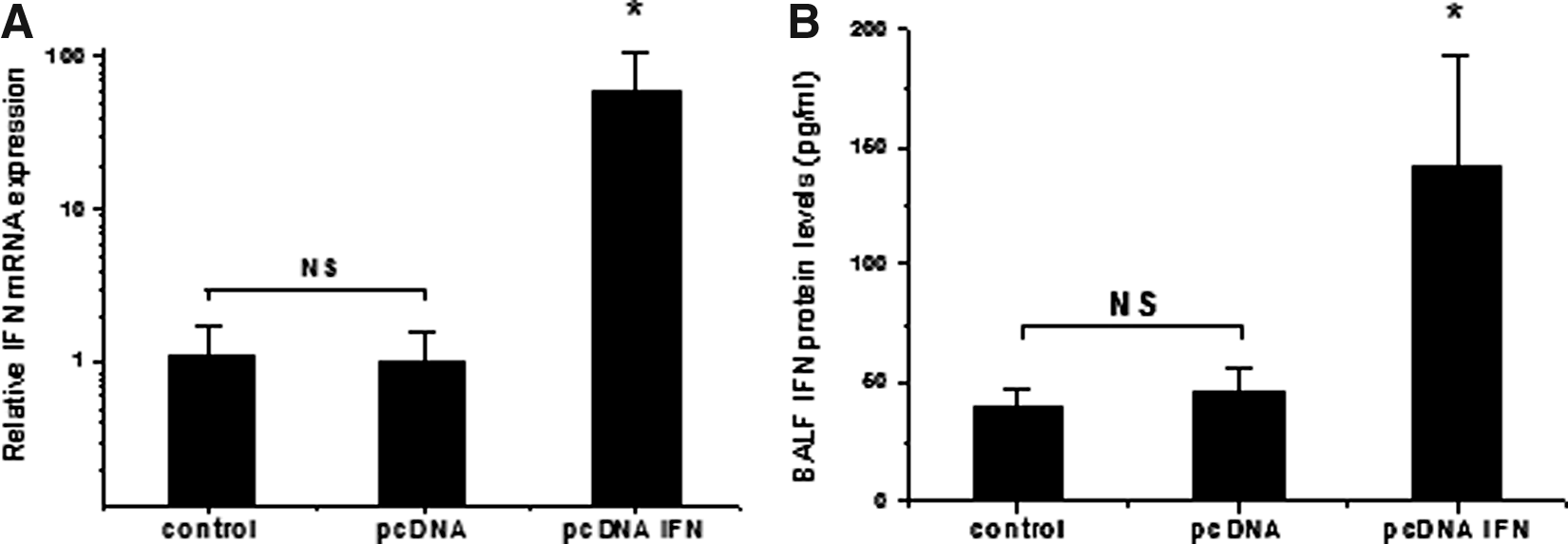
The expression levels of IFNγ mRNA and protein in different treatment groups.
Further experimental analyses revealed that aerosol PEI/pcDNA-IFNγ complexes increased IFNγ protein expression in BALF. The enhanced effects of IFNγ protein were similar to the increased expression of IFNγ mRNA after aerosol PEI/pcDNA-IFNγ complexes to lungs. Protein expressions of IFNγ were 40.47±6.49, 46.20±10.13, and 142.88±45.11 following no aerosol or application with 2 mg of empty plasmid or PEI/pcDNA-IFNγ, respectively. IFNγ protein levels were significantly increased compared with those of the nonaerosolized or empty plasmid groups (p<0.05 for both; n=5 for each treatment group; Fig. 2B). These results indicated that aerosolized delivery of PEI/pcDNA-IFNγ enhanced the mRNA level and protein concentration of IFNγ in mouse lung tissues.
The effect of PEI/pcDNA-IFNγ on inflammatory cytokine expression in lung tissues
Once the transfection of IFNγ via aerosol delivery of PEI/pcDNA-IFNγ had been confirmed by its enhanced mRNA expression in the pcDNA-IFN group, we examined the changes in the mRNA levels of the inflammatory cytokines IL-1, IL-6, IL-10, TNF, and GM-CSF that were the result of this transfection. The mRNA levels of IL-1, IL-6, and GM-CSF were significantly decreased 24 hr after treatment with PEI/pcDNA-IFNγ compared with that of the PEI/pcDNA3.1 group. In the PEI/pcDNA3.1 and PEI/pcDNA-IFNγ treatment groups, mRNA expressions of IL-1 were 1.00±0.73 and 0.03±0.03, respectively (p=0.018); those of IL-6 were 1.00±0.78 and 0.06±0.10 (p=0.028), those of IL-10 were 1.00±1.28 and 0.70±0.40 (p=0.635), those of TNF were 1.00±0.23 and 0.68±0.25 (p=0.067), and those of GM-CSF were 1.00±0.66 and 0.08±0.07 (p=0.015; Fig. 3). These results showed that PEI/pcDNA-IFNγ could reduce mRNA expression of IL-1, IL-6, and GM-CSF, but not TNF-α and IL-10 within a day.

The levels of cytokine mRNA expression in mouse lung tissues in the PEI/pcDNA-IFNγ and the PEI/pcDNA3.1 groups. Total mRNA was extracted from lung tissues in mice treated with aerosolized PEI/pcDNA-IFNγ, and the mRNA levels of IL-1, IL-6, IL-10, TNF-α, and GM-CSF were determined by qRT-PCR. The results showed significant changes in IL-1
The time course of cytokine mRNA expression in lung tissues after aerosol PEI/pcDNA-IFNγ treatment
We investigated the changes in cytokine mRNA levels over time in the mouse lung after PEI/IFNγ aerosol delivery. The mRNA levels of IFNγ in the lungs reached a maximum 1 day after aerosol exposure to PEI/IFNγ complexes (p<0.05, n=5). The change in mRNA on the third day was less than on the first day, but remained at a level higher than that of the control group (p<0.05, n=5). The enhanced gene expression of IFNγ returned gradually to that of the controls in the following 4 days (Fig. 4A). Simultaneously, the expression levels of the inflammatory cytokines were attenuated. The lowest levels of IL-1, IL-6, IL-10, TNF, and GM-CSF were found from 1 to 5 days after treatment (2.9%, 6.1%, 3.2%, 39%, and 8.1%, respectively, of the levels in the nonaerosolized group) and then returned to almost normal levels (Fig. 4B–F). In particular, the mRNA levels of IL-1, IL-6, and GM-CSF were decreased significantly, and correlated with increased mRNA expression of IFNγ after aerosolized administration of PEI/pcDNA-IFNγ (p<0.05, n=5). However, the mRNA levels of IL-10 and TNF were not decreased significantly after aerosol PEI/pcDNA-IFNγ application within a week (p>0.05, n=5). This result indicates that increased IFNγ expression induced by aerosol PEI/pcDNA-IFNγ concomitantly reduced some of the inflammatory cytokines tested.
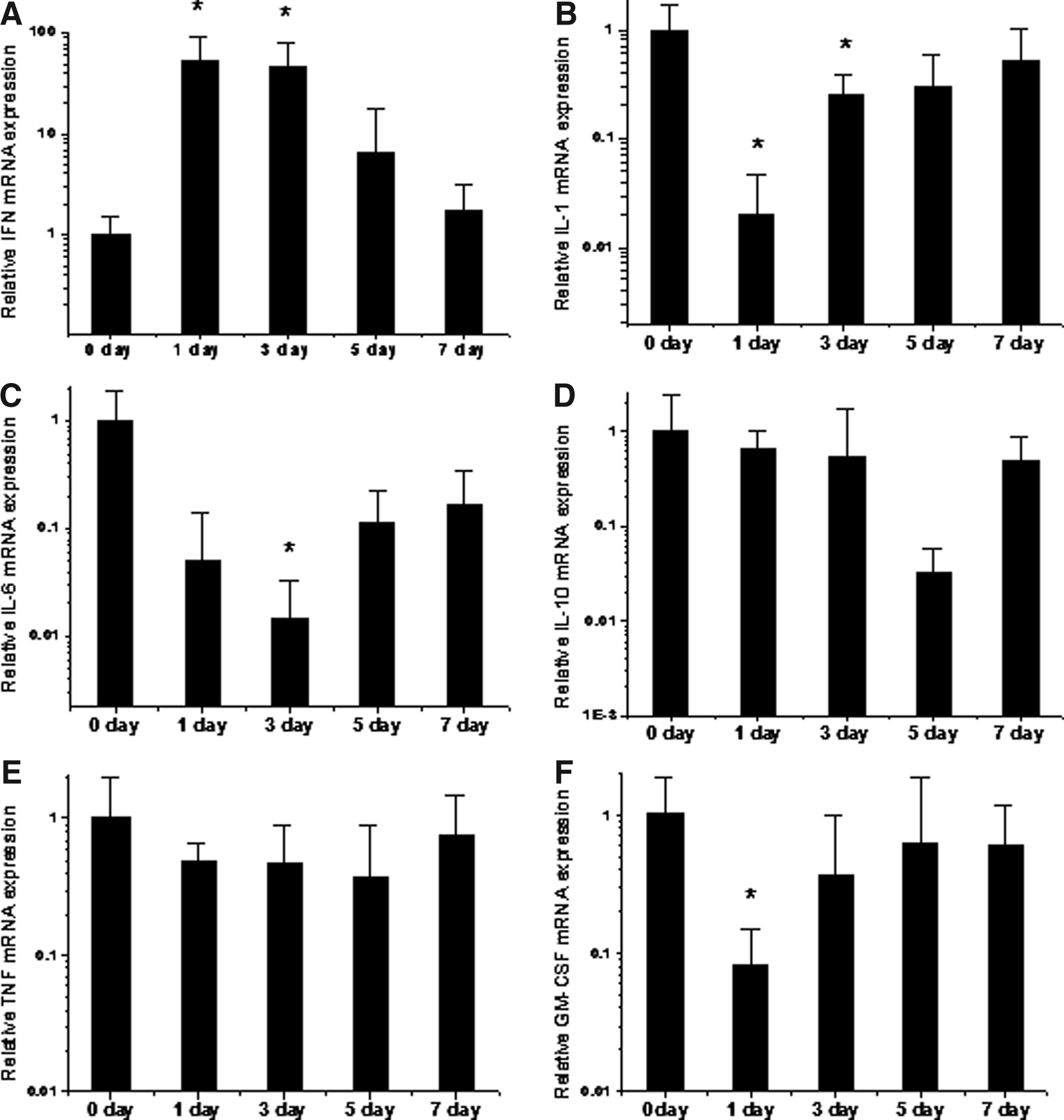
The time-dependent response of cytokine mRNA expressions in mouse lung tissues after a single dose of aerosol PEI/pcDNA-IFNγ administration. Total mRNA was extracted from lung tissues in mice treated with a single dose of aerosolized PEI/pcDNA-IFNγ at the indicated times, and the mRNA levels of IFNγ, IL-1, IL-6, IL-10, TNF-α, and GM-CSF were determined by qRT-PCR. The results (bar graphs) show the mRNA levels of IFN
Discussion
In contrast with the traditional means of delivery, direct inhalation gene therapy to the lung delivers relatively high doses with less adverse effects.(13) To date, there have been no reports regarding aerosol delivery of polymer-gene complexes that resulted in increased expression of IFNγ in lung tissues. The present study reports changes in inflammatory cytokines in murine local lung tissue after administration of inhalant PEI/pcDNA-IFNγ.
To improve gene transfection efficiency, in the present study plasmids were complexed with PEI, which is one of the most effective polymer carriers to deliver gene material into tissue.(14) In particular, PEI has been shown to protect the gene of interest against shear forces during inhalation.(15) Moreover, we used an optimized PEI-DNA ratio and aerosol DNA dose in the experiments, based on previous reports.(16,17) Our results showed that, compared with the control groups, administration of the PEI/pcDNA-IFNγ complex in an aerosol resulted in significantly increased IFNγ mRNA and protein expression in the lung tissue after 24 hr. This is satisfactory transfection efficiency. The results shown in Fig. 3 indicate that PEI/pcDNA-IFNγ could suppress the expressions of other inflammatory cytokines in comparison with the PEI/pcDNA3.1(–) group to different degrees. This is further proof that bioactive IFNγ was expressed. This result agreed with other investigations that IFNγ could inhibit expression of cytokines such as IL-1, IL-6, IL-10,(18–20) and GM-CSF(21) in macrophages, airway smooth muscle cells, and T cells. Furthermore, the IL-1, IL-6, IL-10, TNF, and GM-CSF cytokines showed a valley of change along with a peak expression of IFN after aerosol treatment with a single dose of PEI/pcDNA-IFNγ, within a week (Fig. 4). A possible explanation could be that after aerosol delivery of the gene into cells, IFNγ may be released slowly under the influence of PEI, which could result in increased target gene expression. Subsequently, enhanced IFNγ expression could reduce the production of other cytokines. On the other hand, exogenous DNA is unstable in mammalian cells and is degraded to acid-soluble fragments after a few days.(22)
The change in cytokines was significant in these experiments. However, the effect of IFN on inflammatory cytokine production is complex. For example, inflammatory cytokine production regulated by IFNγ is species-specific.(23) This study is limited in that the effect of PEI/pcDNA-IFNγ on cytokines was tested only in mouse lung tissue and did not include other species. In addition, although it is believed that PEI improves the efficiency of delivery of a DNA vector without organ toxicity or inflammation in mice,(11) other work still suggests pulmonary cytokine responses associate with PEI-DNA aerosol gene therapy.(24) Previous research showed the enhanced levels of TNF-α were higher than those of IL-1 in BALF induced by aerosol PEI/DNA delivery to lungs at the 24-hr time point,(24) which may be an underlying mechanism to explain the noticeable down-regulation of IL-1, IL-6, and GM-CSF, but not TNF-α and IL-10, after PEI/pcDNA-IFNγ aerosol application. In this and other studies, the effect of PEI on the cytokines regulated via enhanced IFN expression was not examined. We will study these aspects further in the near future.
It has been reported that some pulmonary diseases are associated with the high expression of proinflammatory cytokines. Blockade of their production has a potential role in the alleviation of lung disease.(25) Another report showed that inhibition of IL-6 would be beneficial to alleviate inflammation.(26) Inhibition of GM-CSF may attenuate the severity of COPD and asthma development.(27) Furthermore, increasing IFNγ and blocking proinflammatory cytokines might be considered effective treatment for idiopathic pulmonary fibrosis on the basis of the latest researches.(28,29) Our experimental results show that aerosol IFNγ gene therapy has the potential to regulate the immune response by suppressing expression of cytokines produced in mouse lung. The expression of cytokines has been correlated with respiratory disorders such as asthma.(30) As a result, we think that application of exogenous IFNγ via aerosol gene transfection may be beneficial in the treatment of lung disease by reducing the expression of inflammatory cytokines.
In summary, in the present study aerosol PEI/IFNγ therapy in vivo caused a remarkable increase in IFNγ expression and a decrease in the levels of the inflammatory cytokines IL-1, IL-6, GM-CSF, and so on. This study showed for the first time that induced IFNγ gene expression may modify the production of inflammatory cytokines, thereby attenuating immune responses in mouse lung, and may be useful in the study of immune reactions in human lung diseases.
Footnotes
Acknowledgments
This work was supported by the Natural Sciences Foundation of Liaoning Province (no. 2001101031).
Author Disclosure Statement
The authors declare that no conflicts of interest exist.