
Abstract
Background:
Mycobacterium vaccae vaccine, a composition of Mycobacterium proteins, has been known to have bidirectional immunomodulatory functions. Recent studies have shown that M. vaccae has a therapeutic potential for treating asthma. However, little is known regarding the effect of M. vaccae aerosol inhalation during allergen sensitization or challenge on asthma. The purpose of this study was to explore the effect and the underlying mechanism of M. vaccae aerosol inhalation during allergen sensitization or challenge on airway inflammation in an asthma mouse model.
Methods:
Asthma mouse models were established. Mice received aerosol inhalation with M. vaccae once daily during allergen sensitization or challenge for 5 days successively. Airway responsiveness, bronchoalveolar lavage fluid (BALF) cell count, histology, and cytokine concentrations (IL-4, IFN-γ, IL-10, and IL-17) were measured. The relative mRNA expression of ASC, caspase-1, TNF-α, and IL-1β was also determined. Expression of pulmonary NLRP3 and nuclear factor kappa B (NF-κB) protein was measured using immunohistochemistry and Western blot.
Results:
M. vaccae aerosol inhalation suppressed airway hyperresponsiveness and inflammation, reduced levels of IL-4, upregulated expression of IFN-γ and IL-10 in BALF, inhibited mRNA expression of pulmonary ASC, caspase-1, TNF-α, and IL-1β, and also inhibited expression of pulmonary NLRP3 and NF-κB protein during allergen sensitization or challenge.
Conclusion:
M. vaccae aerosol inhalation can suppress airway hyperresponsiveness and inflammation during allergen sensitization or challenge, and may be a promising approach for asthma therapy.
Introduction
Asthma is a chronic respiratory disease characterized by airway hyperresponsiveness and airway inflammation, which primarily manifests as shortness of breath, chest tightness, and cough.(1) The process of asthma is associated with many factors, such as heredity, environment, allergen, and immune dysfunction. In addition, the incidence of asthma has increased considerably in recent decades to affect more than 300 million people worldwide.(2) Currently, inhaled corticosteroids are used as a main treatment to reduce airway inflammation in people with asthma; however, many problems still remain, including significant side effects and drug resistance.(3) Thus, it is urgent to look for new therapies for asthma.
Mycobacterium vaccae vaccine is mainly composed of Mycobacterium proteins and can be used as an adjuvant treatment for tuberculosis.(4) The M. vaccae has been known to have bidirectional immunomodulatory functions. It has been reported that M. vaccae is able to increase T helper (Th) cell activity and enhance B cell proliferation and antibody secretion.(5) Our previous study showed that M. vaccae could prevent the formation of asthmatic model in mice.(6,7) However, little is known regarding the effect of M. vaccae aerosol inhalation during allergen sensitization or challenge on asthma. The major aim of this study was to explore the effect and the underlying mechanism of M. vaccae aerosol inhalation during allergen sensitization or challenge on airway inflammation in an asthma mouse model.
Materials and Methods
Ethics approval
The protocols used in this study were approved by the Guangxi Medical University Ethics Committee (Approval No. 202005007).
Animals
Balb/c mice (male, 4–6 weeks, about 20 g) were purchased from Changsha Tianqin Biotechnology Co., Ltd. (License No. SCXK [Xiang] 2019-0014, Changsha, China). The mice were housed under specific pathogen-free laboratory conditions at suitable temperature (23°C–25°C) and humidity (40%–50%) with food and water ad libitum.
Asthma mouse model
Balb/c mice were randomly assigned into four groups (n = 6 mice/group): normal control (NC) group, asthma model with M. vaccae (Anhui Longkema Biological Pharmaceutical Co., Anhui, China) aerosol inhalation during allergen sensitization group (M.v./Sen), asthma model with M. vaccae aerosol inhalation during challenge group (M.v./Cha), and asthma model group (OVA). Mice in the M.v./Sen group and M.v./Cha group received aerosol inhalation with 22.5 μg M. vaccae dissolved in 10 mL saline once a day during allergen sensitization or challenge for 5 days successively, and mice in the NC group or OVA group received aerosol inhalation with saline. Four groups received aerosol inhalation using an ultrasonic nebulizer (WH-2000; Yuehua Medical Instrument Factory, Guangdong, China; particle size: 1.0–5.0 μm) in a closed chamber (20 × 30 × 20 cm, length, width, height, respectively), 30 minutes per time, once per day. The asthma model was based on our previous study.(8)
Briefly, each mouse was injected intraperitoneally with an OVA (Grade V; Sigma, St. Louis, MO) mixture (comprising 25 μg OVA, 1 mg aluminum hydroxide adjuvant, and 0.2 mL phosphate-buffered saline [PBS]) on days 0, 7, and 14. From days 21 to 28, they were challenged with 2% OVA (400 mg OVA dissolved in 20 mL PBS) using an ultrasonic nebulizer as described above, 30 minutes per time, once per day (Fig. 1).
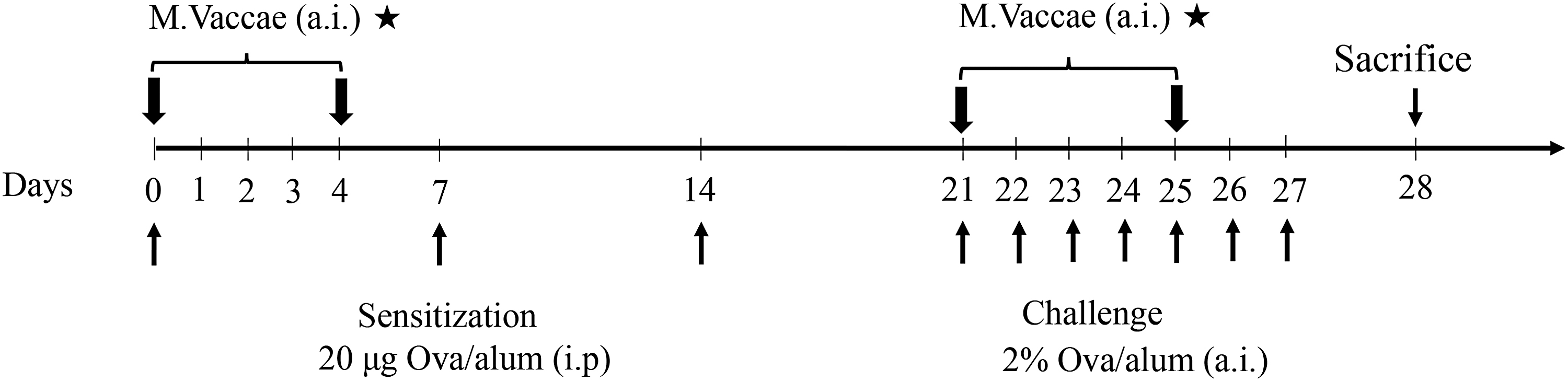
Time line of experimental protocol. Mice were aerosol inhalation with Mycobacterium vaccae during allergen sensitization or challenge. a.i., aerosol inhalation; i.p., intraperitoneally.
Evaluation of airway responsiveness
Airway responsiveness was determined via a noninvasive lung function machine (Fine-Pointe™ NAM system TBL4500; Buxco, Wilmington, NC) after the final challenge. The airway responsiveness was measured by specific airway resistance, and the detection method was referenced to our previous study.(9) Briefly, after adaptation of testing chamber for 5 minutes, mice were exposed to aerosols of PBS and methacholine (Mch; Sigma-Aldrich) at concentrations of 6.25, 12.5, and 25 mg/mL for 3 minutes separately.
Sample collection
The mice were anesthetized with 1% sodium pentobarbital (60 mg/kg intraperitoneal). The lungs were lavaged three times with 0.5 mL PBS, and bronchoalveolar lavage fluid (BALF) was obtained. The BALF supernatant was frozen at −80°C for cytokine analysis. The left lungs of mice were fixed with 4% paraformaldehyde for histology. The right upper lobes were immersed in RNAlater reagent (Solarbio, Beijing, China) for real-time reverse transcription polymerase chain reaction (RT-PCR) analysis. The right lower lobes were frozen at −80°C for Western blotting analysis.
BALF cell count and cell classification
The BALF was centrifuged at 1000 rpm for 10 minutes to separate cells and the supernatant, and the cells were resuspended in PBS. The BALF cells were counted on a cell counting plate. Cell classification was determined using Wright–Giemsa staining (Solarbio), and a minimum of 200 cells were counted under optical microscopy.
Histology
The left lungs were fixed with 4% paraformaldehyde, embedded in paraffin, and sectioned into 3-μm-thick slices. Hematoxylin and eosin (HE) staining was used to evaluate inflammatory cell infiltration, and periodic acid–Schiff (PAS) staining was used to determine goblet cell hyperplasia and mucus secretion. The severity of peribronchial inflammatory cell infiltration was scored semiquantitatively as follows: score 0 means no inflammatory cell; score 1 means a few inflammatory cells; score 2 means a ring of inflammatory cells 1 cell layer deep; score 3 means a ring of inflammatory cells 2–4 cells deep; and score 4 means a ring of inflammatory cells >4 cells deep. The goblet cell (PAS-positive epithelial cell) hyperplasia in airways (internal diameter: 100–300 μm) was scored semiquantitatively as follows: score 0 means no goblet cells; score 1 means <25% of the goblet cells; score 2 means 25%–50% of the goblet cells; score 3 means 50%–75% of the goblet cells; and score 4 means >75% of the goblet cells,(10) the percentage of PAS-positive cells was calculated from the number of PAS-positive epithelial cells per bronchus divided by the total number of epithelial cells in each bronchus.
Measurement of cytokines in BALF
The concentrations of cytokines (IL-4, IFN-γ, IL-10, and IL-17) in BALF were measured with the corresponding ELISA kit (Cusabio Biotech, China). The absorbance was measured by a microplate reader (Varioskan LUX; Thermo Fisher) at 450 nm.
Real-time PCR of lung tissues
cDNA amplification was performed with SYBR-Green Premix (Takara, Otsu, Japan). Real-time PCR reactions were performed with a 7500 Real-Time PCR System (Applied Biosystems). Levels of mRNA expression were analyzed using the 2−ΔΔCT method with β-actin as the internal control. All the primer sequences shown in Table 1 were designed and synthesized by TsingKe Biological Technology Company (Beijing, China).
Primer Sequences in Reverse Transcription-quantitative Polymerase Chain Reaction (RT-qPCR)
Immunohistochemistry
The paraffin sections (3-μm thick) were dewaxed, rehydrated, blocked with 0.3% H2O2, antigen repaired, and 10% goat serum closed. Subsequently, sections were incubated with the rabbit anti-NALP3/CIAS1 antibody (1:200 dilution; Bioss, Beijing, China) and rabbit anti-nuclear factor kappa B (NF-κB) p65 (1:400 dilution; Cell Signaling Technology, Inc., MA) at 4°C for 24 hours. And then, sections were incubated with a secondary antibody (1:100 dilution; Solarbio), developed color with diaminobenzidine, counterstained with hematoxylin, and differentiated with 1% hydrochloric acid alcohol. Finally, sections were dehydrated and sealed using neutral resin. The result was observed under an optical microscope (Olympus C-5050, Japan) and analyzed by ImageJ software.
Western blot
The total proteins were extracted. Proteins (50 μg) were separated using a 10% sodium dodecyl sulfate–polyacrylamide gel, and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore). The membranes were blocked with a blocking solution for 15 minutes, and incubated with the rabbit anti-NALP3/CIAS1 antibody (1:1000 dilution) and rabbit anti-NF-κB p65 (1:1000 dilution) at 4°C for 24 hours. Then the membranes were incubated with the anti-rabbit IgG (H+L) (1:10,000 dilution; Cell Signaling Technology, Inc.) for 60 minutes, and the images were detected and analyzed via the LI-COR Odyssey system (LI-COR Biosciences).
Statistical analysis
One-way ANOVA with post hoc least significant difference (LSD) analysis was used to assess differences between groups and data are presented as mean ± standard deviation. The semiquantitative data of the histopathological parameters were analyzed using the Kruskal−Wallis nonparametric test. Data were analyzed using SPSS version 22.0 software, and p < 0.05 was considered to indicate statistical significance. #p < 0.05, ##p < 0.01, and ###p < 0.001 versus the NC group; *p < 0.05, **p < 0.01, and ***p < 0.001 versus the asthma group.
Results
Airway responsiveness
Compared with the NC group, the airway responsiveness in the OVA group was significantly increased when mice were exposed to aerosols of Mch at concentrations of 12.5 and 25 mg/mL (p < 0.001). Compared with the OVA group, the airway responsiveness of the M.v./Sen group and M.v./Cha group was significantly decreased when mice were exposed to aerosols of Mch at concentrations of 12.5 and 25 mg/mL (p < 0.05). However, airway responsiveness did not differ significantly when mice were exposed to aerosols of Mch at concentrations of 6.25 mg/mL (Fig. 2).

Results of airway responsiveness to methacholine. Data are expressed as mean ± SD. Cha, challenge; M.v., Mycobacterium vaccae; M.v./Cha, asthma model with M. vaccae aerosol inhalation during challenge group; M.v./Sen, asthma model with M. vaccae aerosol inhalation during allergen sensitization group; Mch, methacholine; NC, normal control group; OVA, asthma model group; SD, standard deviation; Sen, sensitization; sRaw, specific airway resistance. ###p < 0.001 versus the NC group; *p < 0.05, **p < 0.01, and ***p < 0.001 versus the asthma group.
Cell counts and classification in BALF
Compared with the NC group, cell counts and classification analysis showed that the number of total inflammatory cells and the proportion of eosinophils in the OVA group were significantly increased (p < 0.001). Compared with the OVA group, the M.v./Sen group and M.v./Cha group showed lower cells and eosinophil ratio (p < 0.001) (Fig. 3A, B).

Total cell count and cell classification count in BALF cell.
Histology
HE staining analysis showed that there was significant inflammatory cell infiltration surrounding airways in the OVA group compared with the NC group (p < 0.001), and there were significantly lower inflammatory cells surrounding airways in the M.v./Sen group and M.v./Cha group compared with the OVA group (p < 0.05) (Fig. 4A, C). PAS staining analysis showed that there were significant goblet cell hyperplasia and mucus hyperproduction surrounding airways in the OVA group compared with the NC group (p < 0.001), and there were significantly lower goblet cell hyperplasia and mucus hyperproduction surrounding airways in the M.v./Sen group and M.v./Cha group compared with the OVA group (p < 0.05) (Fig. 4B, D).

Histopathology of the lung tissue (magnification 400 × ).
Cytokine levels in BALF
The cytokine concentrations of IL-4 in the NC, M.v./Sen, M.v./Cha, and OVA groups were (4.37 ± 0.80 pg/mL), (15.79 ± 3.34 pg/mL), (13.54 ± 3.06 pg/mL), and (33.09 ± 3.35 pg/mL). The cytokine concentrations of IFN-γ in the NC, M.v./Sen, M.v./Cha, and OVA groups were (334.98 ± 32.46 pg/mL), (214.71 ± 17.75 pg/mL), (252.23 ± 18.23 pg/mL), and (184.13 ± 19.86 pg/mL). The cytokine concentrations of IL-10 in the NC, M.v./Sen, M.v./Cha, and OVA groups were (77.88 ± 4.84 pg/mL), (68.05 ± 6.72 pg/mL), (83.73 ± 6.29 pg/mL), and (56.14 ± 3.74 pg/mL). The cytokine concentrations of IL-17 in the NC, M.v./Sen, M.v./Cha, and OVA groups were (241.13 ± 26.96 pg/mL), (493.86 ± 78.61 pg/mL), (345.45 ± 27.82 pg/mL), and (547.46 ± 88.68 pg/mL). Compared with the NC group, the expression levels of IL-4 and IL-17 in the OVA group were significantly increased (p < 0.001), and the expression levels of IFN-γ and IL-10 in the OVA group were significantly decreased (p < 0.001). However, compared with the OVA group, the expression levels of IL-4 in the M.v./Sen group and M.v./Cha group were significantly decreased (p < 0.001), the expression levels of IL-17 in the M.v./Cha group were significantly decreased (p < 0.001), and the expression levels of IFN-γ and IL-10 in the M.v./Sen group and M.v./Cha group were significantly increased (Fig. 5).
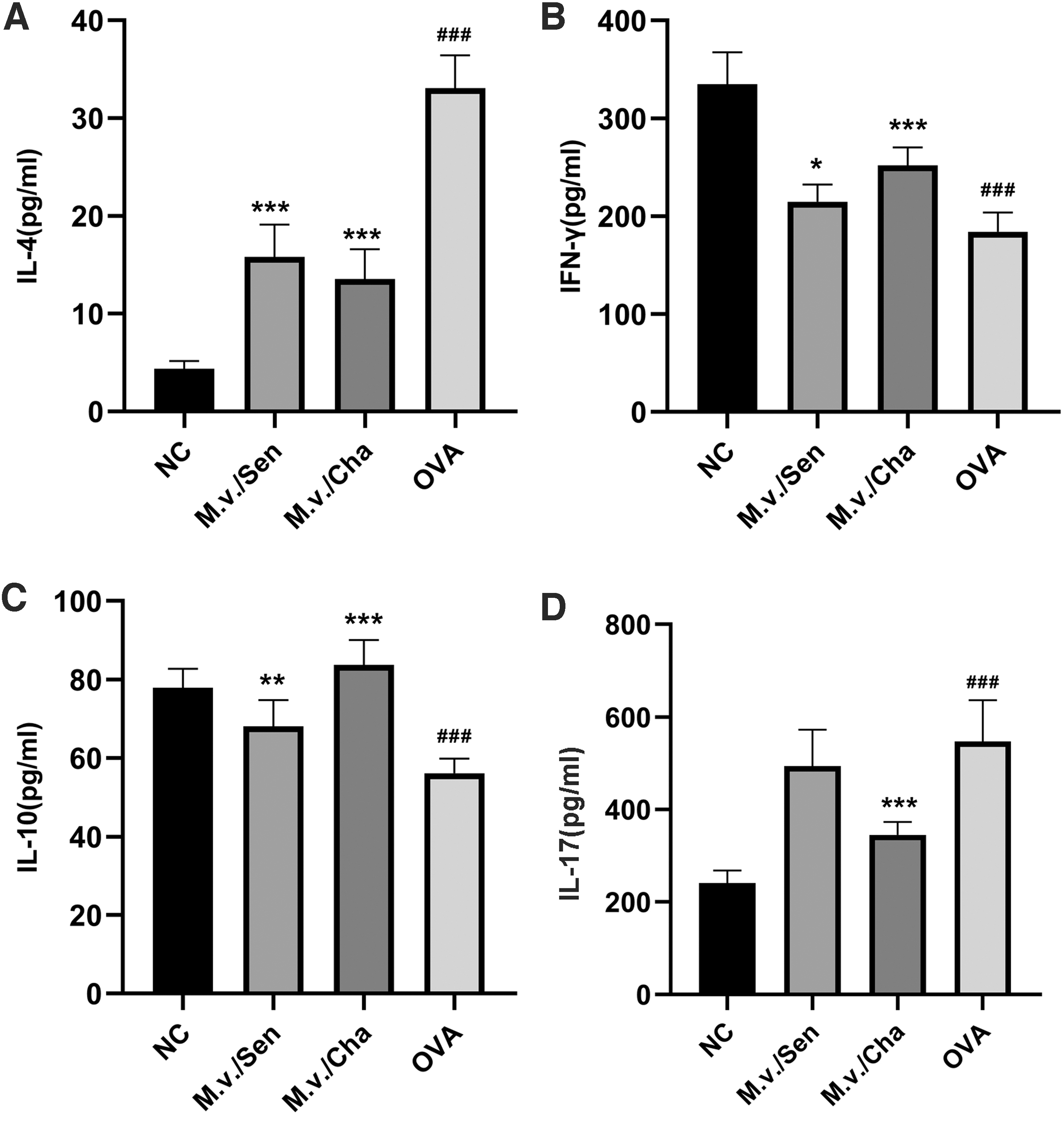
Detection of cytokine concentrations in BALF cell.
Expression of pulmonary mRNA
Compared with the NC group, the relative mRNA expressions of ASC in the M.v./Sen, M.v./Cha, and OVA groups were (2.80 ± 0.26) times, (1.68 ± 0.23) times, and (3.36 ± 0.44) times. The relative mRNA expressions of caspase-1 in the M.v./Sen, M.v./Cha, and OVA groups were (1.87 ± 0.35) times, (1.99 ± 0.42) times, and (2.77 ± 0.29) times. The relative mRNA expressions of TNF-α in the M.v./Sen, M.v./Cha, and OVA groups were (1.54 ± 0.27) times, (1.36 ± 0.22) times, and (1.94 ± 0.41) times. The relative mRNA expressions of IL-1β in the M.v./Sen, M.v./Cha, and OVA groups were (1.26 ± 0.13) times, (1.25 ± 0.11) times, and (1.60 ± 0.19) times. Compared with the NC group, the relative mRNA expression of ASC, caspase-1, TNF-α, and IL-1β in the OVA group was significantly increased (p < 0.01). However, compared with the OVA group, the relative mRNA expression levels of ASC, caspase-1, TNF-α, and IL-1β were significantly decreased in the M.v./Sen group and M.v./Cha group (p < 0.05) (Fig. 6).

Detection of relative pulmonary mRNA expression levels.
Expression of pulmonary NLRP3 and NF-κB protein
Immunohistochemistry analysis showed that the protein staining levels of NLRP3 and NF-κB were significantly higher in the OVA group than in the NC group. Compared with the OVA group, the M.v./Sen group and M.v./Cha group showed significantly lower expression of NLRP3 and NF-κB (Fig. 7A–C). Western blot analysis showed consistent results (p < 0.01) (Fig. 7D, E).
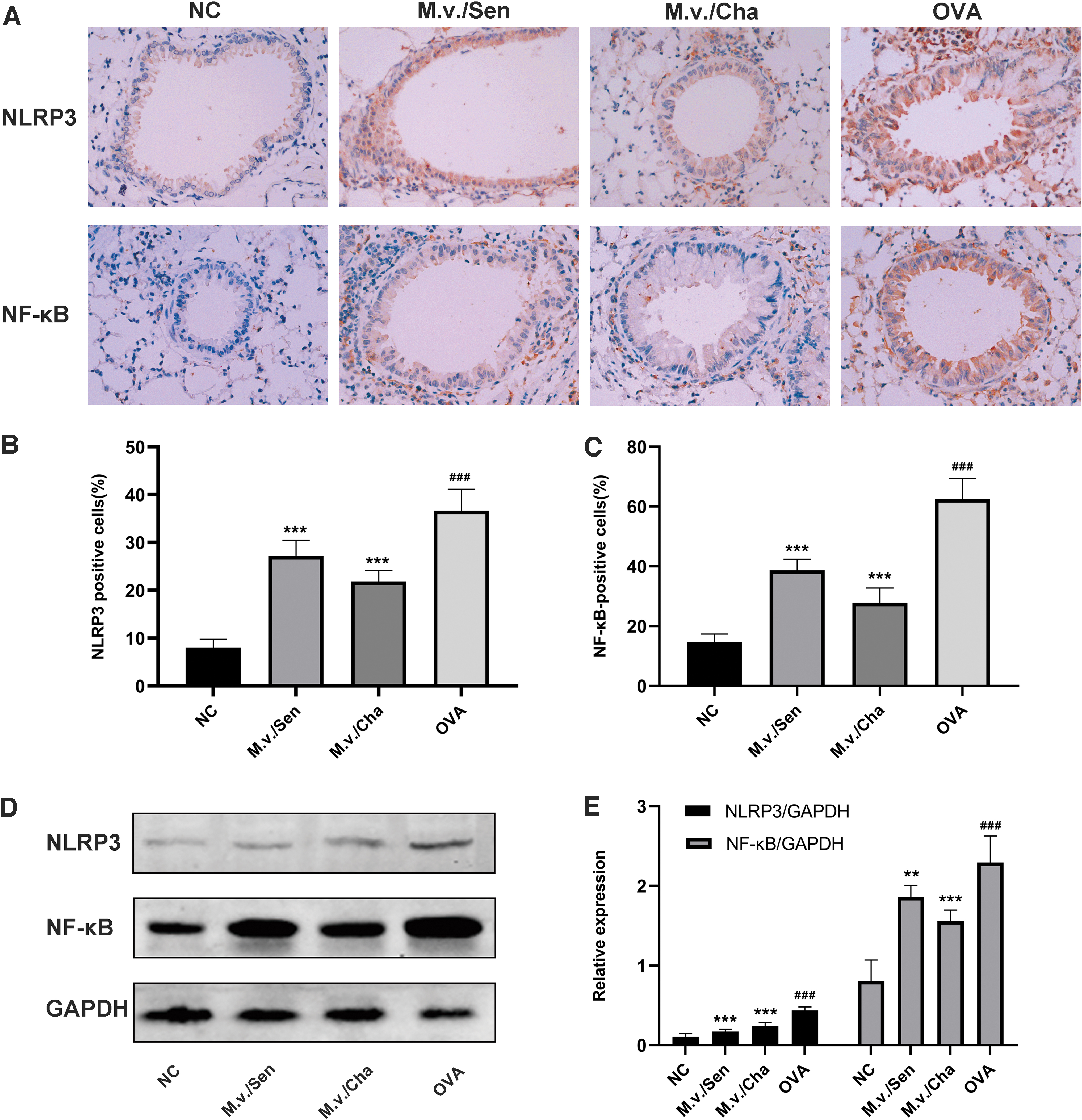
Expressions of inflammation-related proteins in lung tissues analyzed by immunohistochemical staining (magnification 400 × ) and Western blotting.
Discussion
Our study found that M. vaccae aerosol inhalation during allergen sensitization or challenge could alleviate manifestations, attenuate airway inflammation, and reduce mucus secretion in the asthma mouse model through inhibiting the NF-κB and NLRP3 signaling pathways. These results suggest the potential of M. vaccae as a new drug against asthma.
Asthma is a chronic heterogeneous disease and hard to be cured.(11) Shirakawa et al.(12) reported that there was a strong inverse association between delayed hypersensitivity to Mycobacterium tuberculosis and atopy. Previous literatures have confirmed that Bacillus Calmette–Guerin (BCG) and other mycobacterial preparations have protective effects against allergic asthma.(13,14) Our previous results also showed that Mycobacterium phlei aerosol inhalation could improve asthma symptoms and suppress airway hyperresponsiveness in adults(15) and children(16) with moderate persistent asthma. However, BCG could potentially have adverse reactions such as local skin redness, ulcer, and even anaphylactic shock. Compared with BCG and other mycobacterial strain, the M. vaccae vaccine used in this study is mainly composed of Mycobacterium proteins rather than mycobacterial strain, and could lower the possibility of potential side effects. Interestingly, our previous results demonstrated that M. phlei aerosol inhalation could significantly increase IL-10 and IFN-γ in bronchoalveolar lavage, moreover, a 5-day course of M. phlei was superior to a 3- and 4-day course of M. phlei in reducing airway inflammation.(8) We refer to the previous treatment courses and used a 5-day course of M. vaccae for the treatment of asthma mouse model during allergen sensitization or challenge.
Although the etiopathogenesis of asthma has not yet been clearly identified, the imbalance of Th1/Th2 and Th17/regulatory T cells (Tregs) affects inflammatory responses in asthma.(17) Zuany-Amorim et al.(14) reported that the expression level of allergen-specific CD4+CD45RB(Lo) Tregs was increased with SRP299 (killed suspensions of M. vaccae) treatment. Smit et al. reported that M. vaccae treatment downregulated the Th2 cytokines IL-4 and IL-5 in the BALF, however, they did not observe an upregulation of the Th1 cytokine IFN-γ in the BALF.(18) In another study, treatment with M. vaccae observed an upregulation of Th1 response.(19) In the present study, we detected the cytokine levels of IL-4 and IFN-γ in the BALF to assess whether there was a Th1/Th2 imbalance, and we investigated the cytokine levels of IL-10 and IL-17 in the BALF to assess whether there was a Th17/Tregs imbalance. We found that a 5-day course of M. vaccae treatment during allergen sensitization or challenge markedly elevated the cytokine levels of IFN-γ and IL-10. Moreover, M. vaccae treatment during allergen sensitization or challenge evidently reduced the number of eosinophils and inhibited IL-4 level in BALF. The discrepancies in results may be explained partly by the differences in study design, such as the mycobacterial strain and its preparation, mode of administration, the dosage, and time of administration of M. vaccae.
NF-κB as a critical proinflammatory transcription factor regulates various inflammatory genes, including TNF-α and IL-1β,(20) and plays an important role in the inflammatory development of asthma.(21) TNF-α is related to asthma airway pathology, and anti-TNF-α therapy could improve airway hyperresponsiveness in asthma patients.(22) IL-1β is involved in a variety of inflammatory diseases, including asthma. The secretion of IL-1β could be regulated by the NF-κB pathway and NLRP3 inflammasome.(23–25) A previous study has demonstrated that M. vaccae may enhance the antitumor efficiency of γδ T cells by stimulating circulating type 1 myeloid dendritic cells to produce IL-12, IL-1β, and TNF-α.(26) However, to my knowledge, the potential regulatory effect of M. vaccae on TNF-α and IL-1β in asthma has not been clarified. In this study, the levels of pulmonary NF-κB, TNF-α, and IL-1β were markedly upregulated in mice with asthma compared with normal mice, which was consistent with prior reports in the literature.(21,24) Our results suggested that M. vaccae lowered the levels of pulmonary NF-κB, TNF-α, and IL-1β in the asthma model mice, which might be an important mechanism to reduce the inflammatory response of asthma.
Researches have shown that NLRP3 plays a critical role in the regulation of inflammation and immune response.(27,28) NLRP3 interacts with the apoptosis-associated speck-like protein ASC, which contains a caspase recruitment domain, recruited and activated procaspase-1, caspase-1, and is a member of the NLRP3 inflammasome complex.(29) The NLRP3 inflammasome complex as an upstream activator of NF-κB signaling is closely associated with the NF-κB pathway.(30–32) In this research, we found that M. vaccae aerosol inhalation during allergen sensitization or challenge lowered pulmonary NLRP3, ASC, and caspase-1 levels in asthmatic mice, and the decreased levels of IL-1β may be related to the decreased levels of NLRP3, ASC, and caspase-1.
In our research, we use nebulizer therapy of M. vaccae vaccine on the asthma model mice, which is a mucosal immunotherapy method with few side effects. Vaccines administered via mucosal surfaces could elicit more cell-mediated immune responses in humans.(33) Moreover, M. vaccae aerosol inhalation is easy to perform due to its noninvasive way. Recently, Jiang et al. studied the preventive effects of M. vaccae on the asthmatic mice model, mice received aerosol inhalation with M. vaccae within 1 month before establishing asthma model, and found that M. vaccae prevents asthma via neural mechanisms.(6) Our study protocol differs in some respects. We investigated whether M. vaccae is either able to suppress the induction of asthma or able to suppress already established asthma in mice, and we found M. vaccae could protect mice against asthma through inhibiting the NF-κB and NLRP3 signaling pathway during sensitization and antigen challenge. Nevertheless, questions remain about whether there are dose-dependent effects and time-dependent effects of M. vaccae or not, and further studies are needed to the confirm dosage and duration of treatment. In addition, the timing of M. vaccae administration is also critical, whether the effect of M. vaccae is related to allergen exposure is worth further study. Lastly, the biologic processes of sensitization and the response to antigen exposure after sensitization are different, and this is an interesting question deserving further investigation.
Conclusion
In conclusion, our study demonstrates that M. vaccae aerosol inhalation during allergen sensitization or challenge could regulate the Th1/Th2 and Th17/Treg balance by increasing the Th1 and Treg expressions; also, M. vaccae aerosol inhalation could significantly reduce the levels of NF-κB, TNF-α, and IL-1β, and inhibit the levels of NLRP3, ASC, and caspase-1. These results indicate that M. vaccae could protect mice against asthma through inhibiting the NF-κB and NLRP3 signaling pathways, and further studies are needed to confirm the anti-inflammatory mechanism.
Footnotes
Author Disclosure Statement
The authors declare there are no competing financial interests.
Funding Information
Supported by grants from the National Natural Science Foundation of China (No. 81470230) and the Guangxi Natural Science Foundation (No. 2020GXNSFDA238003).
Reviewed by:
Adam Wanner
Theresa Sweeney