
Abstract
Background:
Respiratory infections are increasingly difficult to treat due to the emergence of multidrug-resistant bacteria. Rediscovery and implementation of inhaled bacteriophage (phage) therapy as a standalone or supplement to antibiotic therapy is becoming recognized as a promising solution to combating respiratory infections caused by these superbugs. To ensure maximum benefit of the treatment, phages must remain stable during formulation as a liquid or powder and delivery using a nebulizer or dry powder inhaler.
Methods:
Pseudomonas-targeting PEV phages were used as model phages to assess the feasibility of aerosolizing biologically viable liquid formulations using commercial nebulizers in the presence and absence of inhaled antibiotics. The advantages of powder formulations were exploited by spray drying to produce inhalable powders containing PEV phages with and without the antibiotic ciprofloxacin.
Results:
The produced phage PEV20 and PEV20–ciprofloxacin powders remained stable over long-term storage and exhibited significant bacterial killing activities in a mouse lung infection model.
Conclusion:
These studies demonstrated that inhaled phage (-antibiotic) therapy has the potential to tackle respiratory infections caused by superbugs.
Introduction
Antimicrobial resistance (AMR) is a critical threat to human health. Despite international efforts, AMR is still on the rise and the innovation to date has been unable to keep up with emerging resistance. A global rise in multidrug-resistant (MDR) and even pandrug-resistant infections has led to a significant increase in morbidity and mortality with financial burdens. Respiratory infections caused by Pseudomonas aeruginosa are particularly problematic due to its high intrinsic and acquired antibiotic resistance.(1,2) This Gram-negative pathogen is associated with chronic recurrent respiratory infections in cystic fibrosis, bronchiectasis, chronic obstructive pulmonary disease, and pneumonia with high morbidity and mortality.(2–4) The World Health Organization has identified P. aeruginosa as one of the priority pathogens to steer attention toward developing novel antimicrobials.(5)
Over the past decade, inhaled bacteriophage therapy (or phage therapy) has regained attention in the West as a promising treatment option for combating respiratory infections caused by MDR bacteria.(2) Phages are natural predators of bacteria and are highly effective against MDR and pandrug-resistant “superbugs,” including their biofilm forms. Phage therapy exploits obligatory lytic life cycle of phage, in which it adsorbs onto the bacterial cell surface, injects its DNA or RNA, self-replicate, and then causes bacteriolysis to release progenies for subsequent round of infection. Unlike antibiotics, phages cause minimal disruption of the normal microbiota and no major safety concerns have been identified in human use,(2) making them a promising antimicrobial agent in the era of AMR.
This article is based mainly on the content of an invited podium presentation on the topic “Bacteriophages to tackle respiratory infections caused by superbugs” by the first author at the 2021 ISAM Congress. For extensive reviews on inhaled phage therapy, the readers are referred to our recent review(2) or by Bodier-Montagutelli et al.(6)
Phages Against P. aeruginosa
In the literature, examples of Pseudomonas-targeting phages and phage cocktails (i.e., mixture of two or more phages), including phage PELP20, PELI40, PELI41, PELI42, PELI43, PTM1, and PTM2, were assessed for inhaled therapy in vitro(7,8) as well as in preclinical(9–11) and clinical studies.(12) In our study, a collection of anti-pseudomonal phages, including PEV1, PEV2, PEV10, PEV20, PEV31, PEV40, and PEV61, were investigated for potential use in inhalable phage therapy. They belong to both podovirus (PEV2 and PEV31) and myovirus (PEV1, PEV10, PEV20, PEV61, and PEV40). These phages were isolated from a sewage treatment plant in Olympia (WA) by the Evergreen Phage Lab (Kutter Lab). The 7 phages were screened against 90 clinical and MDR P. aeruginosa isolates collected in Australia.(13) Individually, PEV1, PEV20, and PEV61 were each found to be effective against around 60% of the hosts, and over 70% when used in combination.
Liquid Formulation and Delivery of PEV Phages
Liquid formulations provide a quick and simple way of delivering PEV phages for inhalation. Studies have shown that aerosol delivery of phages in liquid formulation can significantly reduce the bacterial burden in the lungs of small animals infected with P. aeruginosa.(9,11) However, nebulizer devices are usually relatively bulky, require regular disinfection, and a power source for atomization. Liquid formulations may also require refrigeration and cold chain storage for transport. Perhaps, most significantly, phages are under mechanical stress during nebulization, which may induce structural damage to the phages, resulting in titer loss. In fact, reduced infectivity of phage caused by nebulization has been reported previously.(14,15) When a mycobacteriophage D29 was nebulized using a vibrating mesh nebulizer (Aerogen Solo), air-jet nebulizer (Pari LC Sprint) and soft mist inhaler (Respimat), the respective titer reductions were 0.4, 3.7, and 0.6 log10 plaque-forming unit (PFU)/mL,(15) demonstrating that selection of a suitable nebulizer system is crucial to delivery of phages in liquid.
We conducted the first study to investigate the effect of nebulization on structural stability of PEV44, which is a myovirus-type phage active against P. aeruginosa.(16) The purpose was to obtain visual evidence on structural change of nebulized phages, and to establish a relationship between these structural changes and the titer reduction. Three different types of nebulizers, including air-jet (Sidestream), vibrating-mesh (Aeroneb Go), and static-mesh (Omron), were used to aerosolize phages, followed by assessment of phage structure and viability. Based on images acquired by transmission electron microscopy (TEM), the phage in the nebulized samples was classified as (i) being intact and (ii) having changed features such as contracted tail, empty capsid (i.e., nucleic acid already ejected) or the tail detached from the capsid (Fig. 1A). Most notably, regardless of the type of nebulizations, the proportion of intact phages was reduced. However, the greatest reduction in the proportion of intact phages occurred using air-jet nebulization as compared with vibrating-mesh and static-mesh nebulizers (Fig. 1B). The reduced proportion of intact phages was reflected by a corresponding drop in the titer (Fig. 1C).

TEM images of PEV44 showing intact phages from the stock suspension (left), phages with contracted tails with full and empty capsids (middle), and phages with empty capsid and uncontracted tail (end)
To further understand the dependence of the structural integrity of phages on air-jet nebulization, we studied phages with varying tail lengths using PEV2 (podovirus), PEV40 (myovirus), and D29 (siphovirus)(17) and established a positive correlation between the titer loss and tail length (Fig. 2). PEV2 has a 65 nm-wide capsid with a short, stubby, noncontractile tail of 10 nm. PEV40 has a 65 nm-wide capsid with a long, straight contractile tail of 140 nm. D29 has a 60 nm-wide capsid with a long, flexible, noncontractile tail of 180 nm. For the short-tailed podovirus PEV2, air-jet nebulization had a negligible impact on its viability and morphological integrity. On the other hand, the proportion of intact phages dropped significantly for PEV40 (from 50% to 27%) and D29 (from 15% to 2%) after nebulization. Tail damage as observed with TEM is likely an important phage deactivation mechanism as it was partly correlated with the titer reduction upon air-jet nebulization. The repeated shear stress and baffle impaction during air-jet nebulization may have caused breakage of phage at the collar position, which is the weakest region of a myovirus with a nonflexible tail(18) such as PEV40.

Cryo-TEM images of PEV2 podovirus, PEV40 myovirus, and D29 siphovirus
On the other hand, D29 with a long flexible tail may have partially tolerated such mechanical stress by absorbing the impact energy through tail motions (e.g., vibrations, stretching, and bending), but led to snapping of the tail into smaller pieces during the process.(17) Thus, it is important to consider the tail length when using an air-jet nebulizer for phage aerosol delivery. In addition to the tail length, different morphotypes of phages, such as tail rigidity, the ability for the tail to become contracted, the presence of appendages on the tail, and/or antennae on the capsid, may also play a role in determining the robustness of phages against the stress of jet nebulization. Further studies involving phages of different class and morphotypes will need to be explored to validate this hypothesis. For vibrating mesh nebulizers, the size of the phages (75–240 nm) is unlikely to have a major impact on the structural integrity as the aperture diameter of vibrating mesh is in the micron range (>10 times the size of phage). Being nano-sized, the phages distribute within the nebulized droplets, which deposit in the lungs after inhalation (i.e., deposition determined by the aerosol not the phage).
Powder Formulation and Delivery of PEV Phages
Formulation of biologics (such as phages) as inhalable dry powders are more demanding due to technological hurdles both the aerosol and phage have to overcome.(19) Yet the powder delivery devices have the advantages of being small, portable, and not requiring electrical power for operation, leading to better patient compliance. Most importantly, cold chain storage is not a necessity. Powder formulations may also have a longer shelf-life than liquids and likely lead to a cheaper cost for the final product.
It is a dual challenge to produce phage powders for inhalation as it requires the powder to be inhalable and the phage to remain biologically stable in the powder. As phages are composed mainly of proteins besides nucleic acids, it makes sense to start with the same strategy that has been used to stabilize protein therapeutics in the literature.(20) Disaccharides are a major stabilizing agent used for biologics, including proteins, nucleic acids, and liposomes in dry state.(19,21) In fact, they were previously shown to be effective in preserving phage in powders.(22–24) Using trehalose, leucine, and optionally a third excipient (surfactant or casein sodium salt) as excipients, phages KS4-M, KS14, and cocktails of phages ΦKZ/D3 and ΦKZ/D3/KS4-M were spray dried with titer loss of 0.4–0.8 log10 PFU, and the resulting spray-dried phage powders were highly dispersible. We produced inhalable and stable phage powders by cospray drying with GRAS (generally regarded as safe)-type excipients, including lactose, trehalose, mannitol, glycine, leucine, polyethylene glycol 3000, and Pluronic F68.(13)
Of these, only lactose, mannitol, and glycine are approved for inhalation delivery, as the number of commercial inhalation products is limited (compared with orally administered drugs). Consequently, GRAS-type excipients are commonly utilized in developing formulations for pulmonary use as regulatory agencies tend to favor these over other excipients.(25) For example, although
By adopting Taguchi experimental design, we have identified that a simple formulation containing sugar (lactose or trehalose) and leucine was sufficient for producing inhalable and stable phage powders with less than 1 log10 titer reduction during production. The stability of phage was increased with the amount of lactose(13) or trehalose(28) in the powder (Fig. 3). But a high sugar content tends to make the powder sticky and difficult to disperse since spray-dried lactose or trehalose is amorphous.
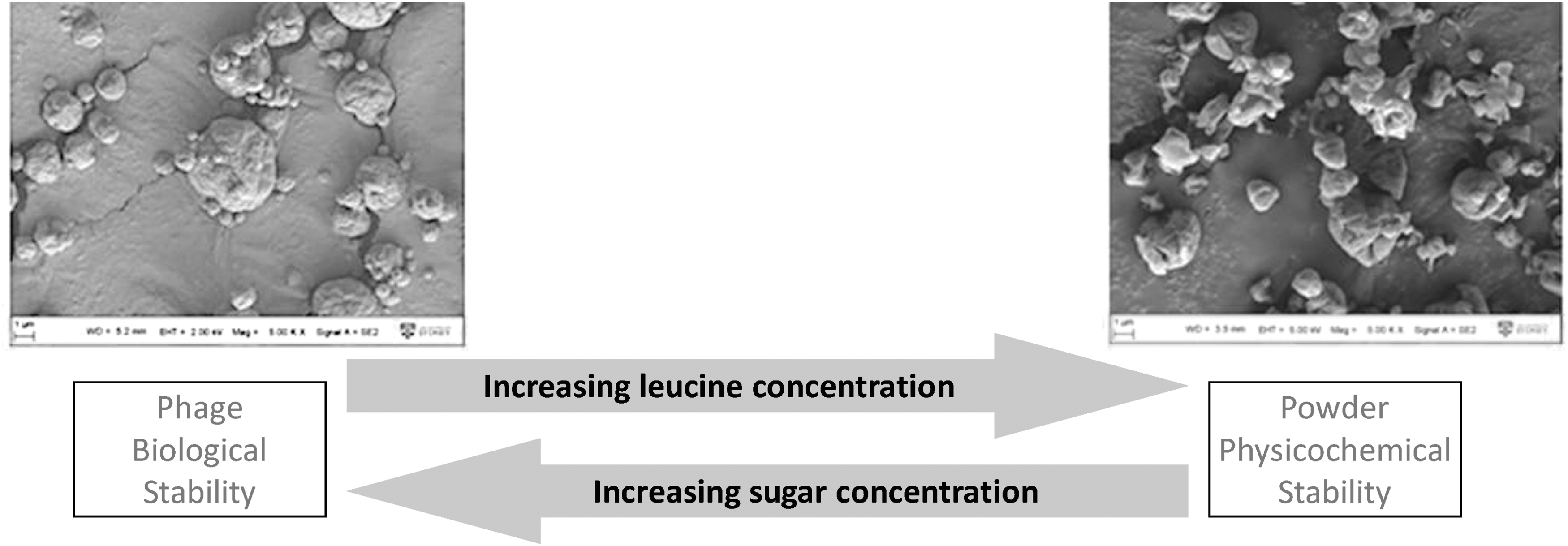
Increasing
In terms of biostability, the phage powders containing more lactose exhibited superior phage protection. For example, in the presence of 90% lactose, PEV61 titer was completely preserved, while ∼1.2 log10 titer loss was observed in those with less lactose. Moreover, biological stability was phage dependent with PEV20 being more prone to inactivation, highlighting the importance of storage stability assessment for each of the phages for clinical development. Overall, both the aerosol performance and phage infectivity of these powders containing disaccharide and leucine were well preserved over storage. In fact, our most recent data show 48-month stability of these powders (unpublished).
In addition to lactose, trehalose (in combination with leucine) has been widely utilized in producing spray-dried phage powders that are well dispersible and stable over storage at room temperature.(30,31) More recently, atmospheric spray freeze drying has been utilized in processing phage D29 into a dry powder formulation by using trehalose and mannitol as stabilizing excipients.(32) Although the powder was not intended for aerosol delivery, the study demonstrated the feasibility of stabilizing phages in solid state using atmospheric spray freeze drying, which would be useful in producing inhalable powder formulations of phages that are heat sensitive.
Mechanistically, disaccharides were proposed to stabilize therapeutic proteins by formation of (i) hydrogen bonds with the proteins, and/or (ii) a glassy matrix to reduce molecular mobility of proteins, and these two processes are not necessarily exclusive in nature.(33,34) To explore the glass stabilization mechanism, we produced spray-dried powder formulations of PEV20 phage with varying contents of lactose and leucine to control their glass transition temperature (Tg) and stored them at different temperatures, Ts (5°C and 25°C), and humidity levels (15% and 33% RH).(35) This resulted in a series of powder systems having various values of (Tg–Ts). According to the glass stabilization hypothesis, it is not the Tg nor the Ts alone that determines the stability of a protein, but rather the temperature gap between them (i.e., Tg–Ts).
As a general rule of thumb, it has been postulated that the protein will remain stable when (Tg–Ts) is great than about 50°C due to reduced local mobility of the molecules.(36) Indeed, our data showed a clear trend that phages are stabilized at storage temperature 50°C below the glass transition temperature (Fig. 4). Phage PEV20 remained stable when the (Tg–Ts) value was above ∼45°C, but became unstable when the value fell below 40°C, supporting the vitrification hypothesis that phage stabilization in solid state is through immobilization inside a rigid glassy sugar matrix.
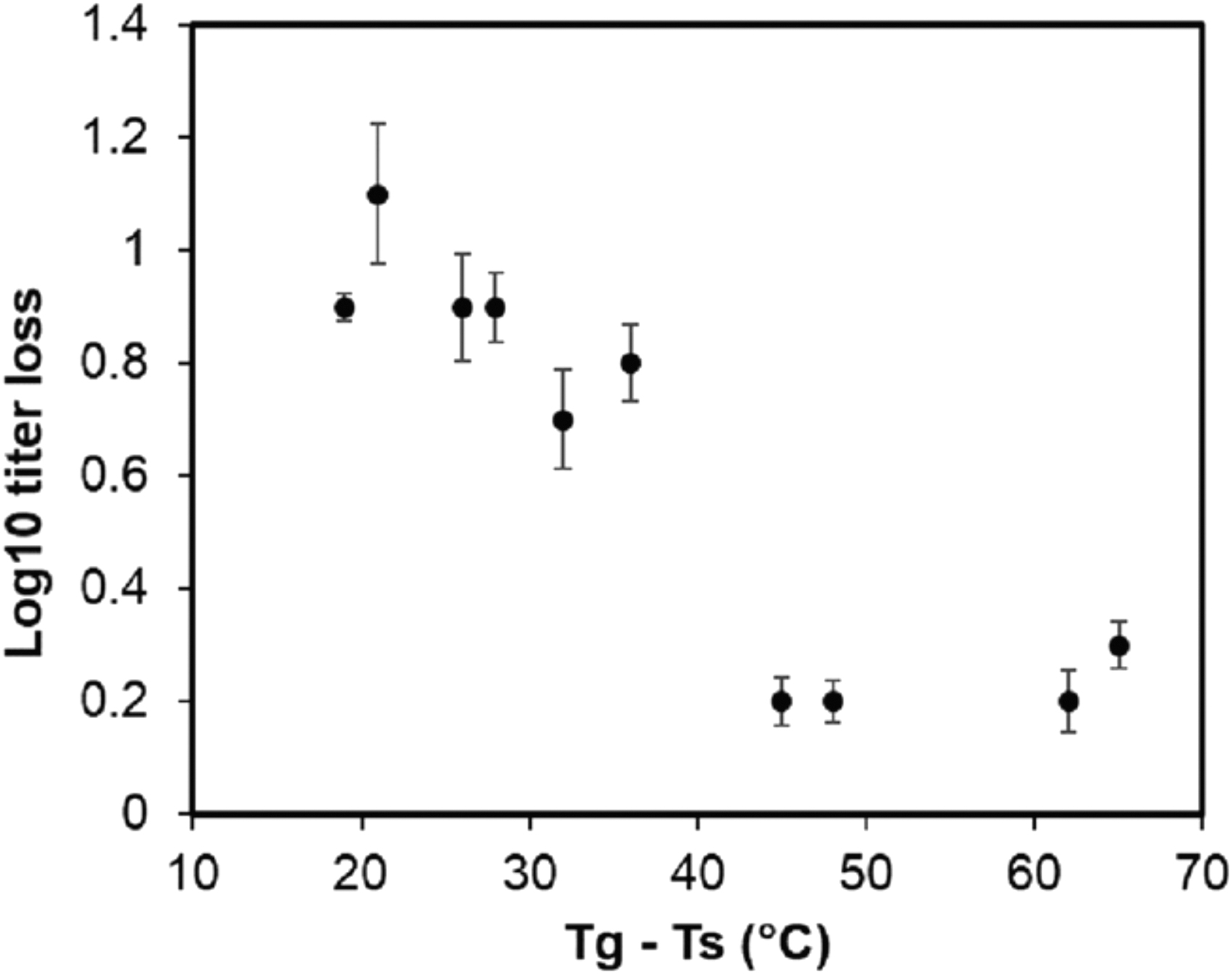
Scatterplot of difference between glass transition (Tg) and storage (Ts) temperatures and log10 titer reduction of phage PEV20. Reprinted from Ref.(35)
Proof-of-Principle In Vivo Efficacy and Safety Testing
We have conducted the first in vivo efficacy study of phage powder formulation using P. aeruginosa lung infection model in mice. A neutropenic mouse lung infection model was adopted to study the in vivo bactericidal activities of spray-dried PEV20 powder delivered directly to the lungs using intratracheal insufflation.(37) The mice were inoculated with a clinical MDR P. aeruginosa isolate (FADDI-PA001), followed by treatment with 2 mg of the PEV20 phage powder containing 2 × 107 PFU/mg at 2 hours postinoculation. At 24 hours postinoculation, as expected, the bacterial load in the lungs of the untreated mice was increased by several log10 (Fig. 5A). In contrast, the bacterial load in the lungs of the phage-treated mice did not increase, which resulted in a 5.3 log10 (p < 0.0005) difference between the two animal groups.
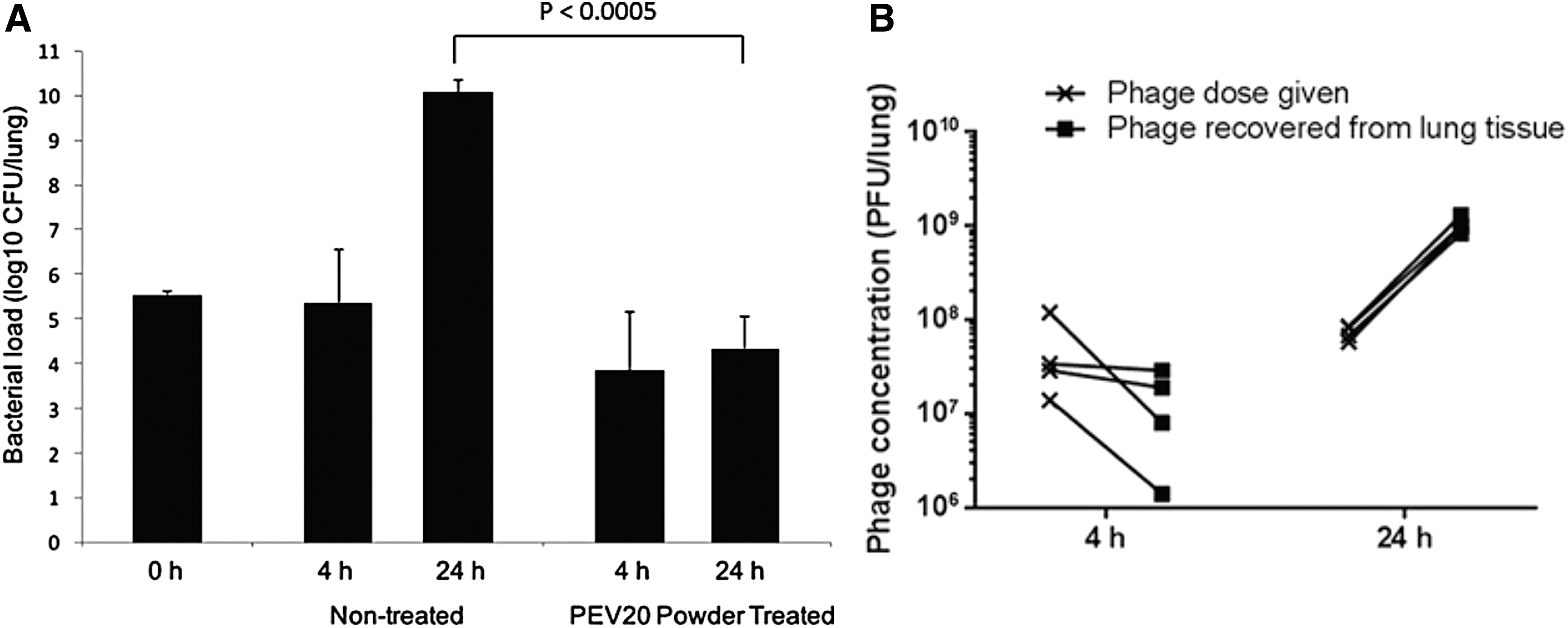
Bacterial loads
Moreover, phages replicated in the lungs of the treated group leading to a 1 log10 increase at 24 hours (Fig. 5B). The histopathological results of the lungs show no major local toxicity of the phage powder. Severe lung damage caused by intratracheal inoculation of bacteria in neutropenic mice was alleviated by treatment with PEV20 powder, suggesting that phage can relieve the lung damage even in the presence of limited host immune system. These results supported the safety and efficacy of pulmonary delivery of phage powder formulation for the treatment of acute lung infection. Despite the promising results, this study showed that inhaled phages can arrest the replication, but are not sufficient to eradicate the bacteria in the lungs. We suspect that multiplicity of infection (MOI) is unlikely to be the reason for incomplete bacterial killing as we have administered the phage powder at an MOI of 100, which is deemed high.
Moreover, in our recent work (unpublished), even MOI of >100 led to incomplete bacterial clearance in the lungs. Two possible reasons for insufficient bacterial eradication could be, first, the bacteria may have developed resistance to phages in the lungs during the time course. Second, the neutropenic mice used in this study lacked immunity. It has been postulated that the immune system is essential for phage therapy.(38) In an acute pneumonia mouse model caused by MDR P. aeruginosa, the host immune response was shown to be essential to control phage-susceptible and emergent phage-resistant bacteria.(39) Efficacious phage therapy requires innate immune components such as neutrophils to work in synergy with therapeutic phage to promote complete clearance of bacteria from the infection site.
Therapeutic dose of phage is poorly established in phage therapy. Due to the self-replicating nature of phages with unique pharmacokinetic and pharmacodynamic profiles in the presence of its host bacteria, the inhaled dose required to observe therapeutic efficacy in humans is unknown. There are two ongoing phase 1/2 clinical trials to assess the safety and tolerability of inhaled phages in subjects with lung infections [NCT04596319 and NCT04684641], and we anticipate that information on potential therapeutic dose would be available soon as the field further progresses.
For clinical use, it is feasible to increase the concentration of liquid phage formulations to 4 × 1011 PFU/mL or higher, which can then be diluted in saline to achieve the desired concentration. The delivered dose can also be increased by lengthening the inhalation time. Similarly, phage powders can be produced at a higher concentration by incorporating a high titer of phage lysate in the feed suspension, followed by spray drying. For example, by using phage lysate at 4 × 1011 PFU/mL, the phage concentration in the powder would be around 109 PFU/mg. If the filled weight of a capsule is 30 mg, then there would be 3 × 1010 PFU of phages available for inhaled delivery. Moreover, patients can always administer more than one capsule, if needed, to achieve the desired lung dose.
Phage and Antibiotic Combination
Phage used in combination with antibiotics can synergistically kill bacteria and minimize emergence of resistant isolates while favorably reversing bacteria to become antibiotic susceptible.(40) We screened the in vitro synergistic efficacy of phage PEV20 with approved antibiotics for inhalation, including colistin, aztreonam, tobramycin, ciprofloxacin, and amikacin, at a concentration below their minimal inhibitory concentration (MIC) against three different MDR P. aeruginosa hosts [FADDI-PA001, JIP865, and 20,844 n/m(s)] by monitoring the bacterial growth over 24 hours.(41) The three isolates exhibited varying susceptibility to PEV20 with FADDI-PA001 being the most susceptible, followed by JIP865 and then 20,844 n/m(s). Among the tested phage and antibiotic combinations, PEV20 exhibited the most synergistic effect with ciprofloxacin. Synergistic killing against FADD1-PA001 over 24 hours was achieved using PEV20 at a low MOI (phage:bacteria ratio) of 0.1 together with ciprofloxacin or amikacin at just a quarter of their respective MIC (Fig. 6).
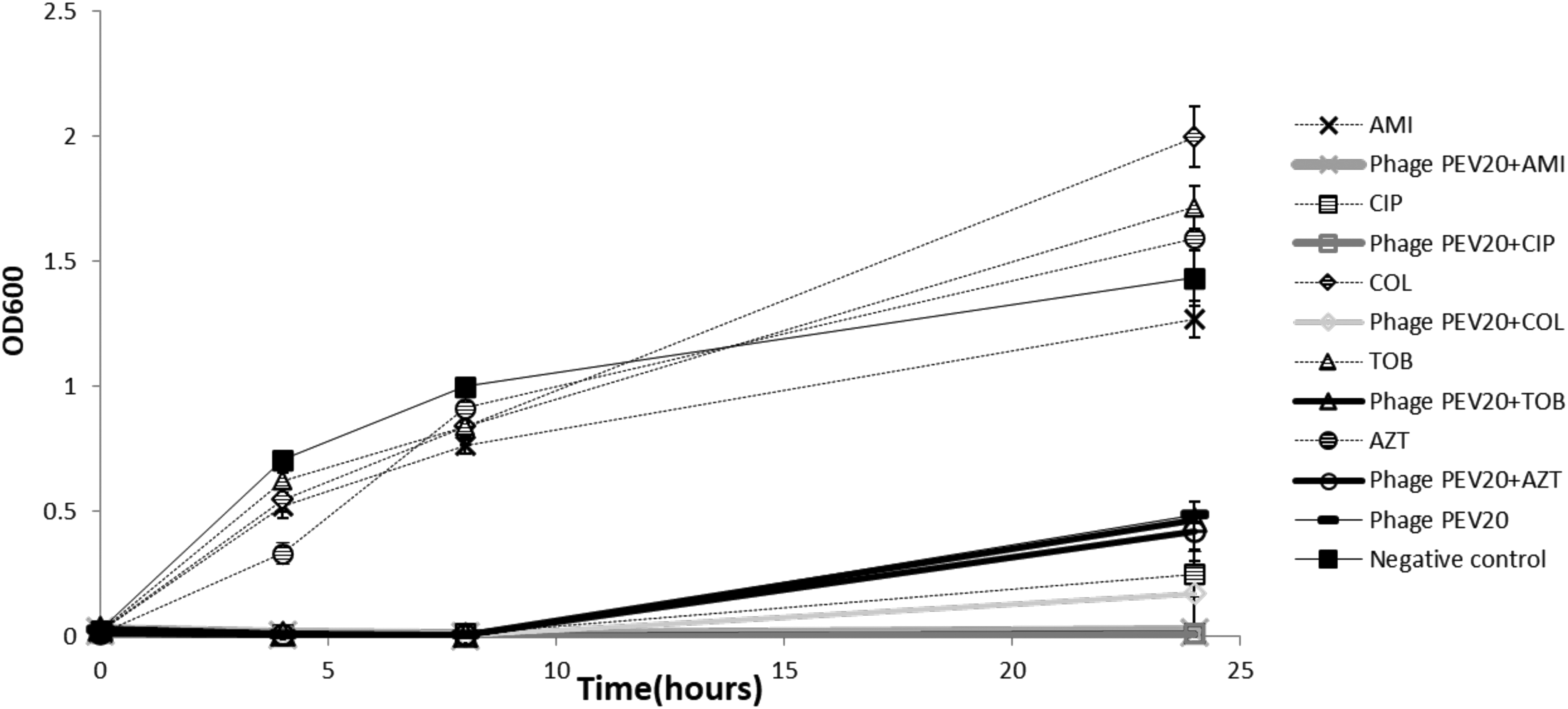
Antibacterial activity of phage PEV20 (MOI of 0.1) against Pseudomonas aeruginosa FADDI-PA001 in the presence of 1/4 × MIC of CIP, AMI, COL, TOB, and AZT. Reprinted from Ref.(41) AMI, amikacin; AZT, aztreonam; CIP, ciprofloxacin; COL, colistin; MIC, minimal inhibitory concentration; MOI, multiplicity of infection; TOB, tobramycin. Copyright 2018 with permission from Elsevier.
The same bacterial killing was achieved for JIP865, but with ciprofloxacin at an increased concentration (1/2 × MIC) and a much higher MOI of 100. The synergistic bacterial killing of FADD1-PA001 and JIP865 by PEV20–ciprofloxacin combination was preserved after nebulization using both air-jet and vibrating mesh nebulizers. For the bacterial strain 20,844 n/m(s), none of the PEV20–antibiotic combinations exhibited synergistic killing under the test conditions of 1/2 × MIC of antibiotic and a higher MOI of 1000. It is expected that the PEV phage and antibiotic combination may not be effective against every bacterial strain. Thus, the lack of efficacy against 20,844 n/m(s) does not mean phage therapy will not work, but rather the importance of matching the bacterial host to the suitable phage, and the latter can always be isolated from the nature or by engineering.(42)
A cocktail of phages (i.e., mixture of two or more phages) can also be utilized to cover a wide range of bacterial infections. In fact, phage products, both commercial and those under development, often have multiple phages in the formulation to expand the target host spectrum. However, microbiological assessment of phage susceptibility to the target pathogen before and during the treatment as well as their interaction with antibiotics would be highly beneficial in maximizing the therapeutic efficacy. Depending on this in vitro assessment, phage cocktail can be further optimized by replacing phages that are ineffective with those that induce therapeutic effect with synergistic bacterial killing.
While the above results were obtained from planktonic bacterial cells, P. aeruginosa after colonization in the lungs commonly produces biofilm, which imposes an impenetrable barrier for antibiotics, making eradication of bacteria difficult.(43) In fact, biofilms can tolerate up to 1000 times higher concentrations of antibiotics than planktonic cells. The PEV20–ciprofloxacin combinations were also found to be effective in removing P. aeruginosa biofilms isolated from cystic fibrosis and wound patients.(44) PEV20 (108 PFU/mL) together with ciprofloxacin at concentrations of MIC or 2 × MIC showed distinctly better bactericidal effects than PEV20 alone or ciprofloxacin alone at 3 × MIC (Fig. 7). Furthermore, PEV20–ciprofloxacin combination protected lung epithelial (BEAS-2B) and fibroblast (HFF-1) cells from P. aeruginosa and promoted cell growth. The addition of phage lowered the ciprofloxacin concentration required to remove biofilms, indicating the potential for implementing lower dosage regimen to help avoid the side effects often associated with high doses of antibiotics.
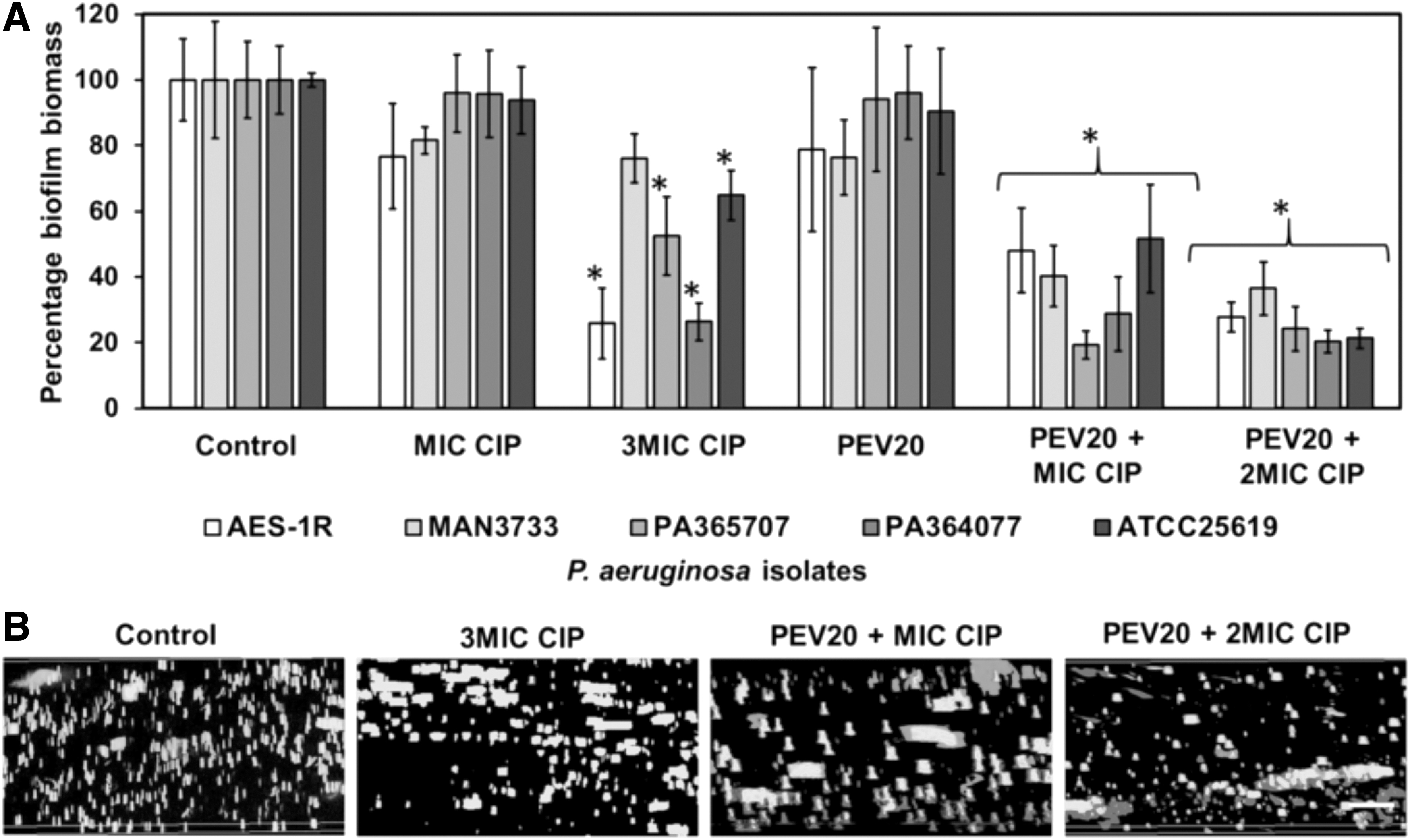
Percentage biofilm biomass after 24 hours treatment with CIP alone (MIC or 3 × MIC), phage PEV20 along (108 PFU/mL), or CIP (MIC or 2 × MIC) combined with PEV20 (108 PFU/mL). Crystal Violet assay
Powder formulations of the PEV20–ciprofloxacin combination were produced by cospray drying with lactose and/or leucine as excipients.(45) The rationale of the formulations containing 1/3 and 2/3 of ciprofloxacin and PEV20 at (108–109 PFU/mL) was based on a 32.5 mg clinical dose of ciprofloxacin.(45) In our previous study, PEV20 at 108 and 109 PFU/mL in combination with ciprofloxacin at 1/4 × MIC and 1/2 × MIC, respectively, exhibited synergistic killing on each of the two clinical stains (FADDI-PA001 and JIP865). As such, a single dose of ciprofloxacin was proportionally reduced to ∼8 and 16 mg in the combination powder formulations. These powders were produced under operating conditions commonly used for spray drying of heat-labile biologics: inlet temperature 60°C, aspiration 35 m3/h, liquid feed rate 1.5 mL/min, and atomizing airflow 742 L/h.
The powders were highly dispersible with a FPF of 60%–64% and infectivity of the phage preserved in the powders after aerosolization by a low-resistance Osmohaler at 100 L/min. Dispersion using a high-resistance Osmohaler at 60 L/min resulted in a FPF of 71%–73%. By coformulating phages and antibiotic together as a single inhalable formulation, the burden on patients will be reduced by lowering the frequency of administration. Moreover, it will increase the likelihood of lung deposition of both antimicrobial agents, thereby exerting synergistic antibacterial killing while minimizing the emergence of resistant mutants.
The synergistic bactericidal action of the powder formulation was confirmed in vivo in the P. aeruginosa mice lung infection model.(46) The mice were inoculated intratracheally with ∼106 CFU (colony-forming units) of clinical strain P. aeruginosa FADD1-PA001, followed by treatment 2 hours later by insufflation of PEV20–ciprofloxacin powder. The combination powder significantly reduced the bacterial load in mouse lungs by almost 6 log10 (Fig. 8), accompanied by reduced inflammation in the lungs associating with bacterial infection. Strong lung monocyte/macrophage and CD8+ T cell response associated with P. aeruginosa infection in the control group was significantly lowered in mice treated with PEV20–ciprofloxacin combination powder. In contrast, PEV20 (106 PFU) or ciprofloxacin (0.33 mg) alone treatment did not reduce bacterial growth.

Bacterial killing activity of PEV20 only (106 PFU), ciprofloxacin only (0.33 mg), and PEV20–ciprofloxacin (106 PFU and 0.33 mg, respectively) combination powders in mouse lung infection model at 24 hours post-treatment. Reprinted from Ref.(46) Copyright 2021 with permission from Elsevier.
Complete bacterial eradication was not observed, which reinforced the potential importance of host immune system as these mice were neutropenic. Bacterial resistance might have been developed to PEV20 phages and/or ciprofloxacin, but unfortunately it was not monitored in this pilot study. It is possible that given a longer period of time, a complete bacterial eradication could occur as phage replication continues beyond 24 hours and reaches the concentration required to kill the bacteria synergistically with the antibiotics.
Conclusion
There are several main take-home messages based on our data and experience. First, we recommend mesh nebulizers over air-jet nebulizers for generating phage aerosols, as less structural damage to the phage is likely to occur. Second, it is achievable to produce inhalable powders with phages remaining biologically intact during long-term storage. Third, in a murine acute lung infection model with MDR P. aeruginosa, we have shown phage powders are safe and efficacious. Finally, inclusion of antibiotics in inhaled phage therapy may be necessary to maximize the treatment outcome. For future direction leading to clinical translation, it is necessary to explore further the PK/PD and dose regimen for respiratory infection, whenever possible, using non-neutropenic animal model.
Footnotes
Acknowledgment
H.-K.C. thanks the organizers of the 2021 ISAM Congress for the opportunity to present the invited talk and submit this article.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
The authors acknowledge the financial supports of the Australian Research Council (Discovery Project DP150103953), National Health and Medical Research Council (Project Grant APP1140617), and National Institute of Allergy and Infectious Diseases of the National Institutes of Health under award number R21AI121627 and R33AI121627. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases or the National Institutes of Health.
Reviewed by:
Nathalie Heuze-Vourc'h
Laurent Vecellio