
Abstract
Macrophages appear to have a fundamental role in the pathogenesis of osteosarcoma. These highly diverse plastic cells are subdivided into classical or inflammatory macrophages known as M1 and alternative macrophages, which decrease inflammation and are reparative, called M2. Although primary and metastatic osteosarcomas are infiltrated with M2 macrophages, targeting the M1 macrophages with the immune adjuvant muramyl tripeptide phosphatidyl ethanolamine (MTP-PE) has been the greatest recent therapeutic advance in osteosarcoma. This discrepancy between the presence of M2 and activation of M1 macrophages is intriguing and is likely explained either by the plasticity of M1 and M2 macrophages or nonclassical patrolling monocytes (PMos). To date, MTP-PE has been approved in combination with chemotherapy for nonmetastatic osteosarcoma, but its use in metastatic tumors has not been investigated. In this review, we focus on macrophages, monocytes, and osteoclasts, their role in osteosarcoma, and the potential for targeting these cells in this disease.
Introduction
O
The current standard treatment of localized high-grade osteosarcoma is illustrated in Figure 1. There have been few substantive improvements in osteosarcoma outcomes in recent decades perhaps best exemplified by the disappointing results of The European and American Osteosarcoma Study Group-1 (EURAMOS-1) trial. This phase III study of newly diagnosed high-grade localized osteosarcoma examined maintenance therapy with pegylated interferon (IFN) alpha-2b in patients with a good histologic response (<10% viable tumor cells) to induction MAP (cisplatin, doxorubicin, and methotrexate) chemotherapy. 2 There was no statistical benefit to postoperative MAP plus IFN alpha-2b compared with postoperative MAP alone. Patients with a poor histological response to preoperative MAP chemotherapy (≥10% viable tumor cells) were randomized to postoperative MAP or MAP with ifosfamide and etoposide (MAPIE). 3 This experimental treatment arm led to increased toxicity without improvement in event-free survival.

A standard treatment design for localized high-grade osteosarcoma. Doxorubicin (A), cisplatin (P), high-dose methotrexate (M). *Week 12 chemotherapy administered when adequate healing postoperatively has occurred.
The greatest recent improvement in treatment has been with the introduction of immunotherapy. The Pediatric Oncology Group and Children's Cancer Group initiated Intergroup Study 0133, a phase III randomized trial of patients with newly diagnosed clinically localized osteosarcoma. Interim results were published in 2005 with final results published in 2008. A 2 × 2 factorial design was used in which (1) MAP was compared with MAP and ifosfamide: arms A and B, respectively, Figure 2; (2) the benefit of adding muramyl tripeptide phosphatidyl ethanolamine (MTP-PE) to chemotherapy was also binomially evaluated: arms 1 and 2, Figure 2. MTP-PE (mifamurtide) is a lipophilic synthetic analog of the Bacillus Calmette-Guérin cell wall component muramyl dipeptide. MTP-PE activates monocytes and macrophages, rendering them tumoricidal in canine osteosarcoma and rodent xenograft models.4–6 The addition of MTP to chemotherapy improved 6-year EFS from 61% with chemotherapy alone to 67% when combined with chemotherapy, hazard ratio 0.80, p = 0.08: not statistically significant. In the final 2008 analysis, the 6-year overall survival was 70% with chemotherapy alone and 78% when combined with MTP, hazard ratio 0.71 (95% confidence interval, 0.52–0.96), p = 0.03. MTP-PE is approved for use in Europe, but the future of its use in the United States remains uncertain. Interrogating the alveolar macrophage chimera may provide some supplementary explanations for the benefit of MTP-PE observed in the INT-0133 trial. In 2015, it was found that nonclassical patrolling monocytes control tumor metastasis to the lung and reduced lung metastasis in multiple mouse tumor models that did not include an osteosarcoma model. 7 This may be relevant given the pulmonary tropism of metastatic osteosarcoma. There also have been discrepant results between the established in vitro inhibitory osteosarcoma effect of MTP-PE combined with IFN-γ, an M1-mediated effect, compared with the tendency for tumor-associated macrophages (TAMs) to have similarity to an M2 phenotype [10]. M1 and M2 are different macrophage polarization states that are viewed as a spectrum of continuum of macrophage activation rather than discrete subsets. In a further consideration, in a series of patients with localized primary osteosarcoma, the frequency of CD163(+) (cluster of differentiation 163) M2-type TAMs in osteosarcoma was found to be correlated with that of TIM-3(+) T cells, the most exhausted T cell subset. 8 The conclusion of this research was that CD163(+) M2-type macrophages support the suppression of tumor-infiltrating T cells in osteosarcoma. The immunology of osteosarcoma is an important area of future research with clarification of existing data potentially informative of future therapeutic opportunities. This review details the biology of monocytes, macrophages, and osteoclasts, themselves tissue-resident macrophages, in the context of osteosarcoma.

INT-0133 trial study design.
Monocytes and Macrophages
The mononuclear phagocytic system comprises monocytes, macrophages, and dendritic cells. Although monocytes are considered precursor cells of this system, it is notable that dendritic cells may also directly originate from dendritic cell precursors. 9 Monocytes migrate from the circulation and extravasate through the vascular endothelium to differentiate into either macrophages or dendritic cells. Macrophages can be distinguished from dendritic cells by phenotypic immunohistochemical differences and functional distinctions, in that dendritic cells can leave tissues, whereas macrophages are fixed. Cell surface markers such as F4/80 are differentially expressed by macrophages compared with dendritic cells and F4/80 is a marker of some, but not all, macrophages in mice. It is, however, of limited use in humans in whom it is predominantly expressed in eosinophils. 10 Other genes differentially expressed by macrophages compared with dendritic cells are CD11b, CD18, CD68, and Fc receptor. It is instructive to know that CD68 is expressed on all macrophages and is useful for immunohistochemistry on human tissues, CD11c is expressed on numerous monocyte-derived cells, including macrophages. CD163 is expressed on most tissue macrophages, including human, and LY6C is enriched in monocytic myeloid lineages. LY6C when used with LY6G is a useful method to determine the relative abundance of granulocytes and monocytes or macrophages. A caution pertains, however, in that defining macrophages and myeloid dendritic cell populations based on cell marker expression is not simple as both cell types arise from common myeloid precursors and lack specificity of surface marker expression.
In adult mice, the mononuclear phagocytic system is derived from three sources with ontologic origins that often persist into adulthood as illustrated in Figure 3. 11 (1) The yolk sac produces progenitor cells, which colonize all tissues and have progeny, which remain throughout life as F4/80 bright resident macrophages. Regulation of these lineages is mainly controlled by CSF1R and its ligands CSF1 and interleukin (IL)-34. (2) Langerhans cells are derived from fetal liver potentially from progenitor cells derived from the fetal yolk sac. (3) A lineage of bone marrow origin gives rise to circulating monocytes as well as their progeny F4/80low macrophages and dendritic cells. In this specific, Ly6c+ monocytes give rise to classical Steinman dendritic cells, which are continuously replenished and regulated by FLT3. There are other F4/80low macrophages, which arise from Ly6c+ monocytes, and in lungs, they can coexist with those of yolk sac origin to give a chimeric pulmonary population. Patrolling pulmonary monocytes are Ly6c− in mice. Macrophages are terminal cells of the mononuclear phagocytic system and in mice are phenotypically CD68+, CD11b+, CSF1R+, and F4/80+. Receptor expression profiling of human monocytes can distinguish a classical population, which has a (CD14++CD16−) profile, and a nonclassical population, which has a (CD14+CD16++) profile. These are synonymous with the respective murine monocyte subset, Ly6C+CX3CR1loCCR2+CD62L+, which has a short half-life and occurs in inflamed tissues, and the Ly6CloCX3CR1hiCCR2−CD62L− subset, which has a longer half-life and occurs in noninflammed tissue.12–14
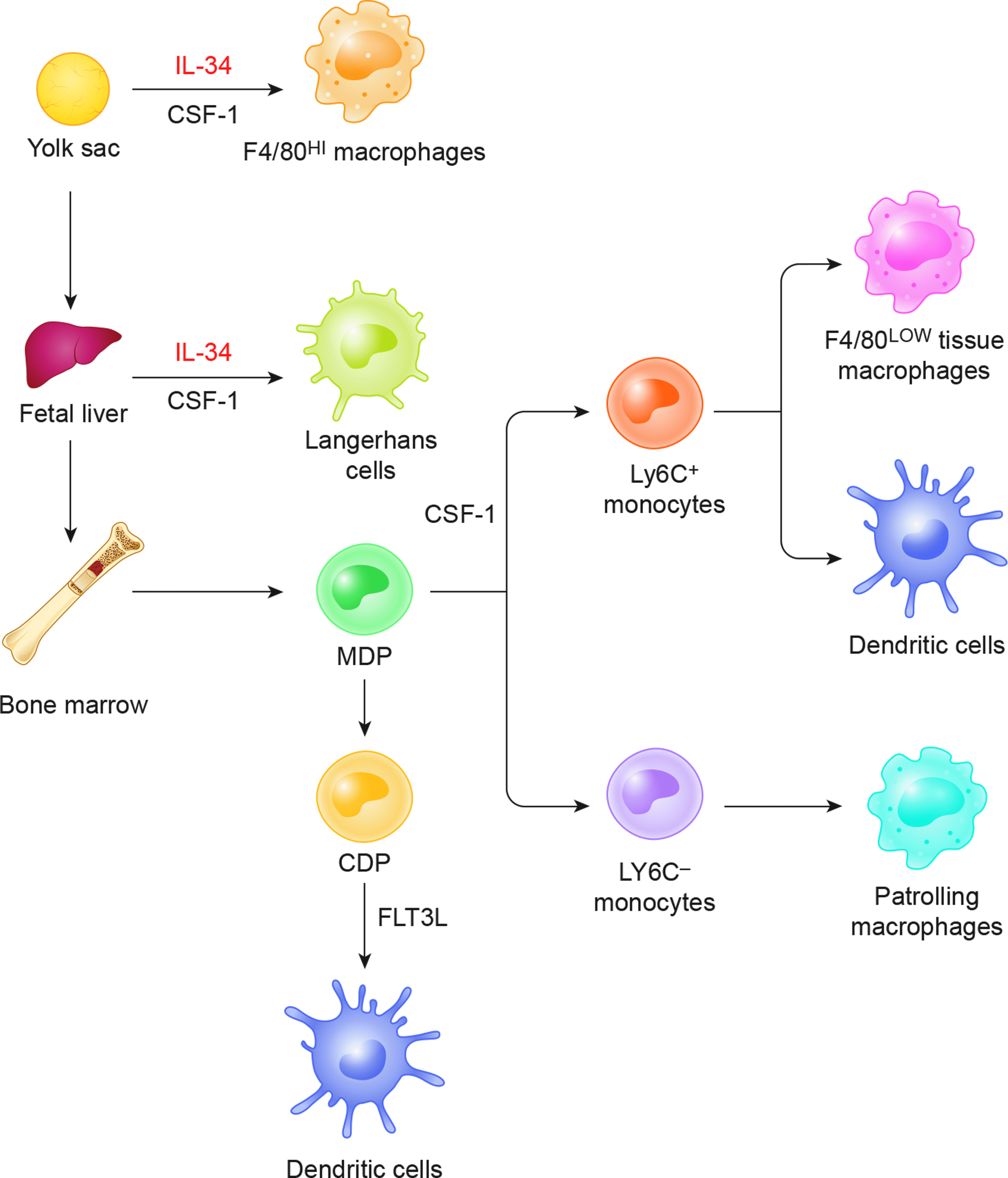
Murine macrophage lineages.
Macrophage Polarization
Monocytes, macrophages, and dendritic cells along with mast cells and neutrophils are professional phagocytic cells. 15 These cells express multiple cell surface receptors that detect signals not normally found in healthy tissues, a feature which distinguishes them from nonprofessional phagocytic cells. For example, professional phagocytes express Toll-like receptor, which is a member of the pattern recognition receptors that detect nonself or damage. Professional phagocytic cells also express scavenger receptors that bind necrotic and apoptotic cell debris and opsonize pathogens. Macrophages are phagocytic cells of metazoan phylogeny that are functionally diverse. They may transit from a resting state to an activated state and finally to a state of hyperactivation when stimulated. Different stimuli activate monocyte–macrophages to become functionally distinct subpopulations, which are polarized to classically activated (M1) or alternatively activated (M2) phenotypes. Peripheral blood monocytes differentiate toward a classical M1 macrophage activation state in response to pro-Th1 stimuli, including IFN-γ+granulocyte–macrophage colony-stimulating factor (GM-CSF) or lipopolysaccharide, such as occurs when eradicating microbes. In contrast, when peripheral blood monocytes are exposed to pro-Th2 stimuli such as IL-4, IL-13, or macrophage colony-stimulating factor (M-CSF), monocytes become anti-inflammatory M2 macrophages.16,17 This is illustrated in Figure 4. Some of the molecules involved in the polarization of M1, M2, and TAMs are documented in Table 1. 18 In a general perspective, TAMs suppress antitumor immunity and may originate from myeloid-derived suppressor cells. 19
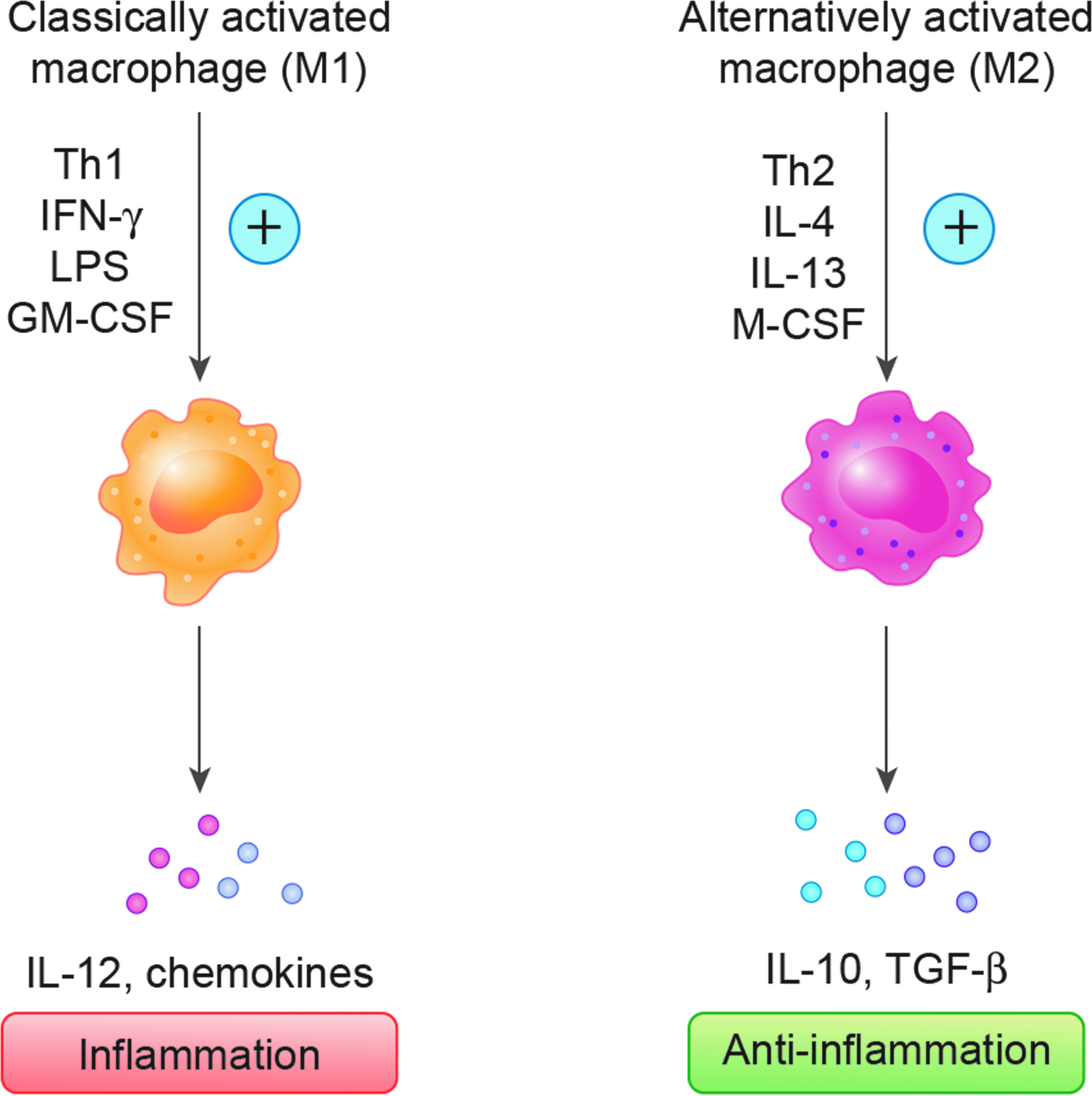
Activator and effector molecules in classically activated and alternatively activated macrophages.
↑, high expression or production; ↓, low expression or production; +, present, −, absent; CCL, chemokine ligand; IFN, interferon; IL, interleukin; M-CSF, macrophage colony-stimulating factor; MHC, major histocompatibility complex; TAMs, tumor-associated macrophages; TGF, transforming growth factor; TNF, tumor necrosis factor.
In vitro models of human osteosarcoma demonstrated that osteosarcoma growth is inhibited by M1-like macrophages when stimulated by MTP-PE, but conditional on being combined with IFN-γ. 20 There is diversity in lineage of origin distribution and variation in transcription profiles of macrophages suggestive of many unique nonoverlapping subclasses in each division. 21 The main historical subclasses of macrophages also have distinct metabolic differences with M1 macrophages metabolizing arginine to nitric oxide, whereas M2 macrophages metabolize arginine to ornithine. Classical monocytes express chemokine receptor type 2 (CCR2) and are recruited to sites of infection by tissues expressing chemokines in particular chemokine ligand 2 (CCL2). The egress of circulating monocytes from the microvasculature to inflamed tissues is dependent on CX3CR1 (CX3 chemokine receptor 1) and CCR2. 22 CCR2 is not present in nonclassical monocytes, but they do express CX3CR1. The ligand for CX3CR1 is fractalkine (CX3CL1), which occurs in a membrane-bound configuration, which supports monocyte adhesion to endothelium, as well as occurring in a soluble form. The binary M1/M2 subclassification of macrophages is not comprehensive and oversimplifies the spectrum of microenvironment circumstance-specific biology of these cells. In addition, there are three other types of macrophages: (1) tumor-associated macrophages, (2) TCR+ macrophages, and (3) CD169+ macrophages. 18 The profile of M1 macrophages is CD68+/CD80+ and that of M2 macrophages is CD68+/CD163+. On a functional perspective, regulatory macrophages, M2 macrophages, TAMs, and myeloid-derived stem cells demonstrate immunosuppressive activity. 23
Greater detail on molecules that are involved in polarization of M1, M2, and TAMs is provided in Table 1. However, a pure categorical approach to descriptors of macrophage activation/polarization is considered less than satisfactory and a standard approach has been proposed based on three determinants: (1) the source of the macrophages, (2) a consensus collection of markers to describe macrophage activation, and (3) definition of the activators. 24 There are also supplementary genetic considerations that determine macrophage activation state.
The activation of macrophages is associated with transcriptional reprogramming, and with this knowledge, the binary M1 M2 model is being supplanted by a spectrum model of human macrophage activation. 25 In one study of 299 macrophage transcriptomes, a spectrum of macrophage activation states was identified with network analysis identifying central transcriptional regulation universally found in all activated macrophages, complemented by regulators related to stimulus-specific programs. TAMs are not a true subset of macrophages as they are not present in steady-state environment, but are identified in several tumors. TAMs are like M2 macrophages; however, evidence is conflicting as to their subset identity, in particular whether they are a discrete M2 myeloid population, and reconciling that they may also exhibit features of M1 macrophages.26–28 This is perhaps a consequence of phenotypic macrophage plasticity. Tumors through elaboration of chemoattractants can recruit circulating monocytes with tumor microenvironments influencing macrophage polarization states. It has been considered that Ly6Chigh monocytes are direct precursors of TAMs as tumor-infiltrating monocytes are predominantly Ly6C+CX3CR1lo. 29 Neither M1 nor M2 like phenotypic polarization has a clear association with survival in osteosarcoma; however, the different polarization states may have different roles in primary and metastatic osteosarcomas with a context-specific clinical effect that merits further research. 30
Patrolling Monocytes and Selectivity of Recipient Metastatic Sites
One subtype of monocytes that may be of potential specific relevance to osteosarcoma are patrolling monocytes (PMos). These cells are CX3CR1highLy6C− in mice and CX3CR1highCD14dimCD16+ in humans. In general, classical monocytes promote tumorigenesis and metastasis; however, nonclassical patrolling monocytes are enriched in the pulmonary microvasculature where they survey the endothelium and have been shown to decrease breast and melanoma lung metastasis in mouse tumor models. 7 The vascular endothelium is important as chemokine stromal cell-derived factor 1 (SDF-1), a type of chemokine, is expressed on the surface of vascular endothelial cells. Its receptor is CXCR4, the mRNA of which is expressed in osteosarcoma and is correlated with metastasis at diagnosis.31,32 PMos scavenge tumor material from the lung microvasculature, prevent cancer cell extravasation, and promote both the recruitment and activation of natural killer cells. Given the proclivity for osteosarcoma to metastasis to the lung and the generic role of PMos in surveying the pulmonary vasculature, examining if PMo function is impaired or subverted in recurrent osteosarcoma merits future research.
Undoubtedly, osteosarcoma has a predilection for metastasizing to the lungs. Tropism for a tumor metastasizing preferentially to a particular site can be partly explained by the interaction between chemokines and chemokine receptors. For example, CXC chemokine receptor 7 (CXCR7) mediates progression of osteosarcoma in the lung where CXCR7 ligand is expressed. 33 Chemokines are a family of homologous cytokines (designated CXCL number) that stimulate the movement of leukocytes and regulate their migration from the blood to the tissues. CXCR7 is a G protein-coupled receptor and is also a scavenger receptor for chemokine 12 (CXCL12). The main function of CXCR7 is recruitment of effector T cells. CXCR7 occurs in both primary and metastatic osteosarcomas with predominant expression in the tumor-associated microvasculature. Experimentally, CXCR7 expression is correlated with distant metastasis. 34 The interaction of fractalkine (CX3CL1) and CX3CR1 in osteosarcoma is also a fascinating area of future research. When monocytes exit the bone marrow, they adopt two different cell fates, one population of which is the cells that exhibit high CX3CR1 expression and low expression of LY6C with a patrolling function. The effect of chemokines and chemokine receptors in osteosarcoma should be investigated with separation of their distinct roles in primary and metastatic disease.
Antitumor Immunity, Cancer, and Osteosarcoma
Phenotypic demonstration that antitumor immunity exists in specific cancers is evidenced by histologic dense lymphocytic infiltrates in specific tumor subtypes such as medullary breast cancer or mismatch repair-deficient colorectal cancer. Indeed, this has been suggested as a surrogate marker of tumor neoantigen load—a vicarious index of the mutational tumor burden and likely susceptibility to checkpoint inhibitor treatment.35,36 Macrophages promote tumorigenesis in carcinomas (cancers of epithelial origin), but in osteosarcoma (mesenchymal origin), tumor-infiltrating macrophages are associated with metastatic suppression, providing a rationale for treatment with macrophage-activating agents. In one study of differential gene expression in a series of prechemotherapy high-grade primary osteosarcomas biopsied, 118/132 differentially expressed genes were upregulated. The techniques used were mRNA profiling and immunohistochemistry. Patients with these upregulated gene profiles were less likely to later develop metastasis. Approximately 50% of these genes had an immunological function especially related to macrophages. These macrophage-related genes were expressed by tumor-infiltrating macrophages and not by the osteoblastic-osteosarcoma lineage. TAMs were M1 (CD14/HLA-DR alpha) and M2-type TAMs (CD14/CD163). TAMs in the primary malignancies were associated with significantly better overall survival.
There have been phase 1 trials of small-molecule inhibitors of macrophage receptors in other malignancies. In pancreatic cancer, a phase 1b trial established that there is additive or synergistic efficacy by combining a macrophage inhibitor targeting CCR2 PF-04136309 with FOLFIRINOX chemotherapy. 37 The combination of FOLFIRINOX and the CCR2-targeted agent was tolerable and safe. In synovial giant cell tumors, an epistatic antitumor benefit arises from a selective macrophage colony-stimulating factor 1 receptor (CSF1R) inhibitor PLX3397. 38 This neoplasm has a t(1:2) translocation, which links the CSF1 gene to the COL6A3 gene. Targeted inhibition of the CSF1:CSF1R interaction is therefore beneficial in this tumor characterized by proliferation of synoviocytes, which attract other cells, including hemosiderin-laden macrophages and histiocytes. The neoplastic clone only constitutes a small minority of the cells in this tumor.
The clinical consequences of macrophages in osteosarcoma may differ between primary and metastatic sites. In addition, the general findings pertaining to carcinomas may be irrelevant to osteosarcoma. Therefore, spatial, tumor-specific, and immune subclass-specific challenges exist in application of the literature. One discovery is, however, of general interest to the field of innate immunity and cancer. 39 The nuclear factor κB pathway is controlled by IKKβ with NF-κB, a ubiquitously expressed transcription factor. It has been found that inhibition of IκB kinase (IKK)β, by deletion in the myeloid linage, causes TAMs to switch to an M1 phenotype, which is cytotoxic to malignant cells. 39 This was associated with increased IL-12 expression, inducible nitric oxide synthase (NOS2), and major histocompatibility complex class II by macrophages. This was mediated by IKKβ role in inhibiting (Stat)1 signaling in macrophages.
Osteosarcoma and Immunity
The host immune system can recognize tumor-associated antigens. Conventional high-grade osteosarcomas have a moderate to heavy leukocyte infiltrate. The principal component of the inflammatory response in osteosarcoma comprises CD14+/CD68+ TAMs as well as CD3 T lymphocytes. 40 DC-SIGN/CD11c+ dendritic cells are also frequently observed when there is a significant tumor-associated macrophage and T lymphocytic infiltrate. M2 macrophages produce transforming growth factor (TGF)-β, vascular endothelial growth factor (VEGF), and contribute to angiogenesis. In osteosarcoma, patients with variant genotype +252 A>G of the TGF-β gene have a poor event-free survival of 20% at 100 months. 41 This variant leads to greater levels of cytokine production. The tumor necrosis family of cytokines, which are noncovalently linked trimers, have a recurring theme of importance in osteosarcoma tumorigenesis. Human osteosarcoma cells, which secrete osteoclast-inducing factors, express tumor necrosis factor (TNF)-α-converting enzyme messenger RNA. 42 In addition, RANKL, which is important in osteosarcoma, is a member of the TNF family. Further considering osteosarcoma in the specific, a xenograft mouse model of human osteosarcoma recruited TAMs that were polarized to the M2 subtype. Experimental removal by macrophage-eliminating liposomes decreased osteosarcoma growth. 43 In human experiments involving peripheral blood and tumor-infiltrating T cells in osteosarcoma, CD163+M2 TAMs supported the suppression of tumor-infiltrating T cells in osteosarcoma and thereby increased osteosarcoma cell evasion of the immune system.8,41
In the AOST0221 phase II trial involving 42 evaluable patients specific to first pulmonary recurrence of osteosarcoma, and administered inhaled GM-CSF, there was no improvement in outcome with a 12-month event-free survival of 20%.44,45 As previously mentioned, peripheral blood monocytes differentiate toward inflammatory (M1) macrophages in response to GM-CSF and Th1. In the AOST0221 trial, the biological endpoint of Fas/Fas ligand expression and presence of dendritic cells were also negative. Some studies found that a high M2/M1 macrophage ratio favors osteosarcoma metastasis and high levels of M1 macrophages decrease osteosarcoma primary tumor burden.46,47 Osteosarcoma cells produce IL-34 and recruit M2 macrophages. In a retrospective series of prechemotherapy osteosarcoma biopsies, the osteosarcoma metastasis-negative group was characterized by a high infiltrate of iNOS+ M1-polarized macrophages and upregulated osteoprotegerin. The osteosarcoma metastatic tumors were demonstrated by increased CD146 cells. CD163 correlated positively with CD146 cells. In this study, TAMs were correlated with a better overall survival. 48
Macrophage recruitment to tumor implantation sites is polarized to the M2 subtype. In a xenografted human osteosarcoma mouse model, deletion of tumor-associated M2 macrophages decreased tumor growth. 43 In another experimental study, CD163(+) M2-type tumor-associated macrophages that produce IL-10 in osteosarcoma were found to support the suppression of tumor-infiltrating T cells. 8 In metastatic osteosarcoma, programmed death ligand 1 is expressed, which binds to the apoptosis-inducing programmed cell death protein-1 (PD-1) receptor expressed on activated T cells. 49 T cell immunoglobulin and mucin domain-containing molecule 3 (TIM-3) occurs in osteosarcoma. 50 Tumor-infiltrating T cells in osteosarcoma contain higher levels of TIM-3+PD-1− and TIM-3+PD-1+ cells than their peripheral blood counterparts. Both PD-1 and TIM-3 are markers of T cell exhaustion (a state of T cell dysfunction, characterized by sustained inhibitory receptor expression, poor effector function, and a transcriptional state distinct from memory and effector T cells) in osteosarcoma. 51 PD-1 and TIM-3 negatively regulate the production of IFN-γ in T cells and can promote apoptosis.52,53 The frequency of CD163(+) macrophages is directly correlated with the frequency of suppressed TIM-3(+) PD-1(+) T cells, the most exhausted T cell subset in osteosarcoma. Therefore, M2 TAMs are correlated with intratumoral T cell-specific immunosuppression and tumor evasion of the immune system. In a potential therapeutic inference, depletion of M2-type (CD163) macrophages improves T cell production of proinflammatory cytokines and T cell proliferation.
In addition, CD4+T cells and CD8+T cells occur in osteosarcoma. 50 For CD4+T cells to be activated, they require major histocompatibility complex II molecules, which are downregulated in M2-type macrophages. Ultimately, reversal of T cell exhaustion is a recognized method to restore antitumor immunity in other malignancies, and anti-PD-1 therapeutics may enter the treatment paradigm in osteosarcoma contingent on the outcome of future clinical trials: examples include a phase II and phase I study of Nivolumab with or without ipilimumab in treating young patients with recurrent or refractory solid tumors or sarcoma (I.D. ADVL1412, NCI-2014-01222, NCT02304458). The apparently detrimental effect of macrophages in metastatic osteosarcoma does not conflict with the better outcome in patients with primary high-grade osteosarcoma described by Buddingh et al. 30 The context and microenvironment differences in primary and metastatic disease cannot be overstated.37,38
Heterotypic RANKL Signaling and Osteoclasts (Multinucleated Tissue-Resident Macrophages)
Cells of the mononuclear phagocytic system are of diverse types and include tissue-resident cells such as Langerhans cells (skin), veiled cells, and interdigitating dendritic cells (lymph nodes), Kupffer cells (liver), alveolar macrophages (lungs), type A cells (synovium), mesangial cells (kidney), and microglial cells (brain). Osteoclasts are specialized tissue-resident macrophages that occupy bone. They are multinucleated cells of monocytic lineage and aggregation, Bone comprises bone cells and bone matrix. Bone cells are osteocytes, osteoblasts, or osteoclasts. Osteosarcoma is a tumor of osteoblastic lineage; however, it has emerged that osteoclasts (cells of monocytic lineage) have a crucial role in the pathogenesis of osteosarcoma. Osteoclasts express the RANK (receptor activator of NF-κB) receptor, whereas osteoblasts produce its ligand RANKL. Functionally, osteoblasts synthesize the organic components of bone matrix comprising type 1 collagen, proteoglycans, and several glycoproteins. Osteoblast activity is stimulated by parathyroid hormone (PTH). Bone matrix is later calcified. Osteoclasts participate in bone resorption and bone remodeling and solubilize calcium from bone.
Bone remodeling and bone metabolism provide important insights into the development of osteosarcoma as exemplified by the increased prevalence of osteosarcoma in patients with Paget's disease of bone and fibrous dysplasia. Molecularly, bone remodeling is mediated by PTH induction of cAMP (adenosine 3′, 5′-monophosphate), which activates protein kinase A to produce RANKL (receptor activator of NF-κB ligand). RANKL regulates osteoblast lineage cells to differentiate into bone-resorbing osteoclasts and also negatively regulates the tumor suppressor gene PTEN. The molecular mechanism of bone remodeling is illustrated in Figure 5. It is noteworthy that protein kinase A has a regulatory subunit called Prkar1α. In genetically engineered mouse models of osteosarcoma, loss of one Prkar1α allele increases the rate of development of protein kinase A hyperactive osteosarcoma through overexpression of RANKL. 54

Molecular mechanism of bone remodeling and osteoclast lineage differentiation.
Mutations in the tumor suppressor genes Rb or tumor protein p53 (TP53) predispose to osteosarcoma and alteration in these genes is found in sporadic cases or in osteosarcomas arising in the context of a familial cancer predisposition syndrome. In conditional gene deletion mouse models, specific deletion of one Prkar1α allele in the osteoblastic lineage when combined with p53Δ/ΔOB mice produces more rapid development of osteosarcoma than biallelic loss of Rb and p53Δ/ΔOB with loss of Prkar1 α causing hyperactivation of protein kinase A. This asserts Prkar1 α as a tumor suppressor gene in osteosarcoma. In genetically engineered mouse models with conditional or whole body deletion of p53, Rb, Rankl, and Prkar1α in cells of osteoblastic lineage, heterotypic RANKL-RANK interactions dominated in the development of osteosarcoma. 55
A different mouse model termed MOTO (murine osteocalcin SV40 T/t antigen osteosarcoma) uses SV40 T/t antigen to inactivate Rb/p53, whereas the afore-described model uses the Col1α1 promoter to express Cre recombinase in committed osteoblasts, thereby creating compound deletions in the p53, Rb, and Prkar1α genes. 54 MOTO-RankΔ/ΔOC mice, which have osteoclast-specific Rank deletion, demonstrate delayed tumor initiation and decreased number of pulmonary metastasis. However, when Rank deletion is restricted to osteoblasts, a minimal effect on osteosarcoma development is observed. Therefore, osteoclast-specific Rank deletion inhibits osteosarcoma genesis, but osteoblast-specific Rank deletion is not sufficient to block osteosarcoma genesis. Figure 6 demonstrated some of these points graphically.
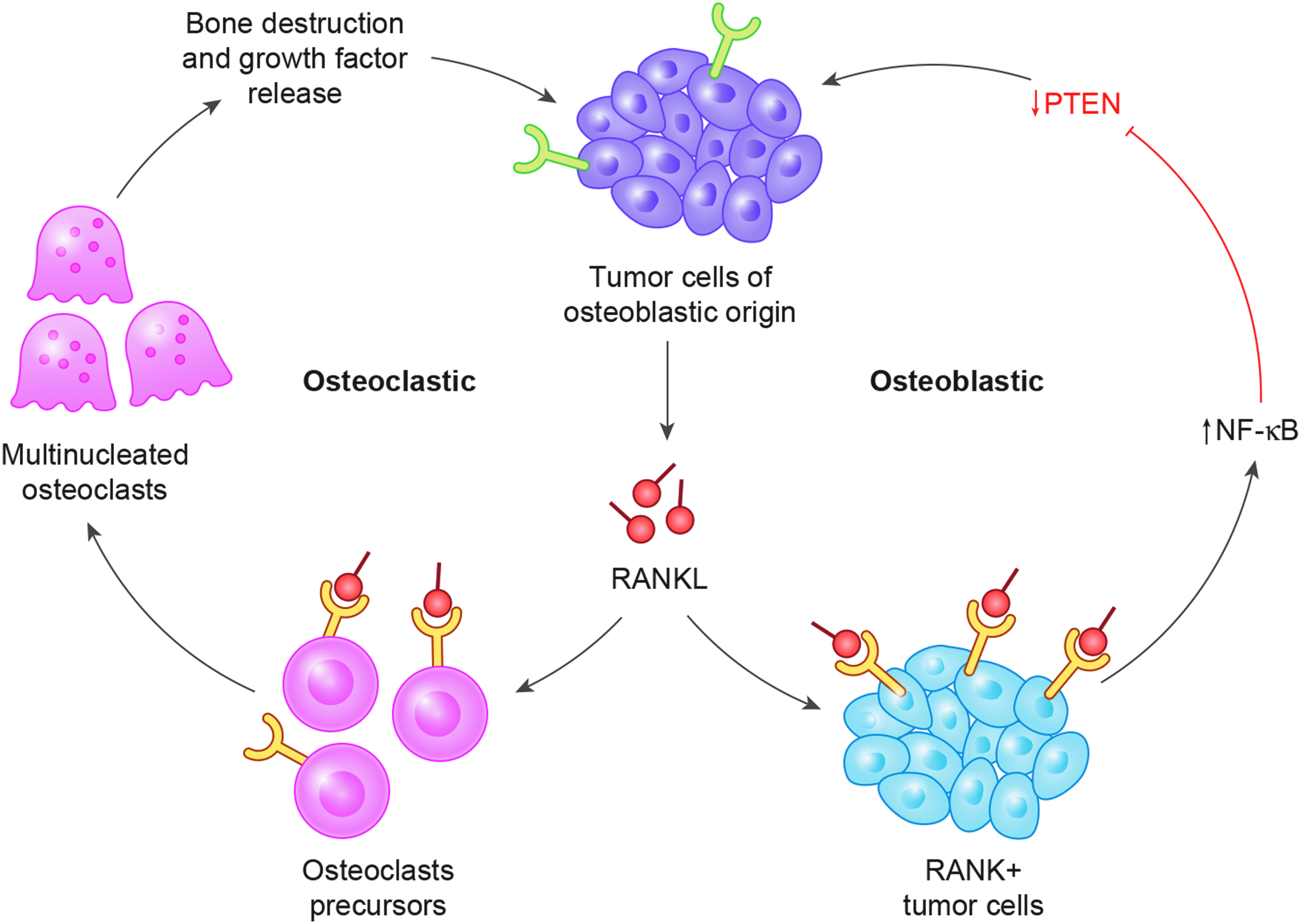
Cellular and molecular biology of local osteosarcoma progression with RANKL from tumor cells causing concurrent osteoblast and osteoclast-mediated effects.
In genetically engineered mice predisposed to develop osteosarcoma, pharmacological inhibition of RANKL with RANK-Fc prevented osteosarcoma development and lung metastasis. Furthermore, pre-emptive administration of RANK-Fc completely prevented development of this sarcoma. This provides a strong rationale for devising clinical trials involving denosumab (a fully human monoclonal antibody RANKL inhibitor) to treat osteosarcoma. A National Cancer Institute-sponsored phase II clinical trial (AOST1321, NCI-2015-00543, NCT02470091) of denosumab in patients with recurrent or refractor osteosarcoma is ongoing.
NF-κB negatively regulates PTEN (tumor suppressor gene phosphatase and tensin) in murine embryonic fibroblasts. 56 In the converse scenario in murine osteosarcoma models, osteoclastic deletion of Rank was associated with upregulation of PTEN. 55 Interestingly, NF-κB mediates dendritic cell transcription profiles. 9 RANKL itself inhibits PTEN. PTEN is a tumor suppressor gene that downregulates the PI3K/AKT pathway. There is increased expression of Pten in primary osteosarcoma tumor cell lines derived from Rankl-null mice compared with MOTO-Rankl+/+ tumor cell lines. In an overview of malignant disease, the PTEN pathway in T regulatory cells (Tregs) is an important driver of the suppressive tumor microenvironment. 57 In a generic perspective on tumor biology, PTEN stabilizes Tregs within tumors to prevent them reprogramming into inflammatory effector cells. 58
Osteosarcoma, Macrophages, and Developmental Pathways
Molecular aberration in osteosarcoma can involve abnormalities of cellular proliferation, immortalization, and disruptions in differentiation. Macrophages also have developmental roles and mediate their developmental activities through WNTs, VEGF, and proteases. Wnt has a differentiation role in the transition of mesenchymal cell to acquisition of osteoblastic differentiation. This is relevant as in murine osteoblasts, Wnt signaling downregulates expression of RANK ligand and inhibits osteoclastogenesis in vitro. 59 Canonical Wnt signaling within osteoblasts also controls osteoclast differentiation with increased stability of β-catenin (the end effector molecule of the Wnt signaling pathway), resulting in increased bone mass, whereas deletion leads to osteopenia. This is caused by β-catenin with TCF proteins regulating osteoblast expression of osteoprotegerin, which negatively regulates osteoclast differentiation. 60 Osteoprotegerin, a bone differentiation marker, is a decoy receptor molecule for RANKL. There have been reports that Wnt signaling is antitumorigenic in osteosarcoma, and dickkopf-1 (DKK-1), a negative regulator of Wnt signaling, can play a protumorigenic role in osteosarcoma. Serum levels of DKK-1 are elevated in patients with osteosarcoma due to its overproduction by tumor cells. In a human xenograft mouse model of osteosarcoma, a monoclonal antibody against DDK-1, BHQ880, inhibited osteosarcoma metastasis. 61 In correlative evaluations, there was increased nuclear staining of β-catenin in the orthotopically implanted patient-derived osteosarcoma xenografts.
Osteosarcoma Therapeutics and Monocytes/Macrophages
The recurring theme of macrophages as important in osteosarcoma biology suggests future avenues of therapeutic possibilities. As detailed nonclassical monocytes express CX3CR1, the ligand for this receptor is fractalkine (CX3CL1). Fractalkine synthetic analogs may have a future therapeutic role in abrogating osteosarcoma pulmonary metastases pending the outcome of future research. Denosumab has a likely future role in osteosarcoma by a heterotropic effect on osteoblasts through inhibiting the RANKL: RANK interaction on osteoclasts, themselves cells of monocytic lineage. MTP-PE has a proven benefit in osteosarcoma concurrent with chemotherapy, but the differential effect on M1 and M2 subclass macrophages needs characterization. Last, exhausted T cell subsets in metastatic osteosarcoma arising in the context of tumor-infiltrating macrophages may be reactivated with anti PD-1 checkpoint inhibitors such as nivolumab or Pembrolizumab. With the prospect of restoring tumor immune surveillance, the recognition of differing context-dependent roles of macrophages between different tumor types, primary and metastatic, and subclass-specific differences in the tumor microenvironment mean that scientific exactitude is required to realize the therapeutic possibilities.
Conclusion
The innate immune system is important in osteosarcoma and understanding its role offers potential new therapeutic strategies. Agents such as PD-1 checkpoint inhibitors can reactivate exhausted T cells in the adaptive immune system, which interacts with cells of innate immunity. The spatial distribution of osteosarcoma (primary, metastatic to lung, metastatic to non-pulmonary sites) and the heterogeneous immune infiltrate in the resected primary will all determine immune-directed therapeutic benefits. Macrophage polarization state, itself a spectrum phenomenon, needs categorical descriptors such as M1 and M2 to assist future therapeutic algorithm approaches. Finally, osteoclasts with heterotropic effects on the osteoblast lineage are amenable to molecular targeting. The 30-year plateau in survival improvement of osteosarcoma may be at an inflexion point of treatment improvement in this exciting new era.
Footnotes
Author Disclosure Statement
No competing financial interests exist.