
Abstract
Interleukin (IL)-15 serves as a survival factor for a broad array of cells. Renal cells express both IL-15 and its receptor (IL-15R); however, the role of IL-15 in the kidney is yet to be determined. We examined IL-15 and IL-15R levels in sepsis-related renal injury, ischemia-reperfusion injury (IRI), and cisplatin-induced nephrotoxicity. To test the anti-apoptotic effect of IL-15, Bcl-2/Bax mRNA levels were assessed in kidneys of IL-15Rα−/− mice and in IL-15-stimulated renal epithelial cells (RECs). In addition, RECs were exposed to cisplatin and apoptosis was evaluated by TUNEL staining, caspase-3 activity, and cell cycle analysis. Intrarenal IL-15 levels decreased 24 h after initiation of all three examined pathologies by 5.8-fold (sepsis), 11-fold (IRI), and 23-fold (cisplatin-induced nephrotoxicity). Further experiments revealed that while addition of rIL-15 (1 ng/mL) to wild-type (WT) RECs increased Bcl-2/Bax ratio by 2-fold, kidneys of IL-15Rα−/− mice exhibited 4-fold lower Bcl-2/Bax ratio compared to WT mice. Accordingly, IL-15 lowered the apoptotic rate in cisplatin-treated cultured REC, and IL-15Rα−/− renal cells exhibited a higher rate of cisplatin-induced apoptosis. Furthermore, IL-15 levels negatively correlated with BUN of cisplatin-treated mice (R = −0.69, P = 0.003), suggesting that a decline in renal-derived IL-15 is detrimental to renal cell survival and kidney function during pathological stress.
Introduction
I
IL-15 utilizes a heterotrimeric receptor complex that consists of two of the three IL-2 receptor subunits (IL-2Rβγc) and an IL-15-specific α-chain. IL-15Rα exhibits an extremely high affinity for IL-15 (K d = 10−11) (Anderson and others 1995; Lorenzen and others 2006). Although IL-15 and IL-15R are constitutively expressed in renal cells (Weiler and others 1998; Tejman-Yarden and others 2005), their role in the kidney has not yet been determined. Recent reports show that IL-15 participates in a unique transpresentation signaling mechanism in which cells that express the IL-15Rα chain can bind to extracellular IL-15 and transpresent it to neighboring IL-15Rβγc or IL-15Rαβγc-expressing cells (Sandau and others 2004; Budagian and others 2006; Bulfone-Paus and others 2006). This mechanism necessarily requires a coordinated synchronized expression of both IL-15 and IL-15R in order to take place (Burkett and others 2004).
In experimental models of acute and toxic renal injury, apoptosis appears to play a dominant role in renal epithelial cell (REC) death that leads to loss of renal function (Arany 2008). Various injuries, including sepsis, ischemia-reperfusion injury (IRI), cisplatin toxicity, mechanical obstruction, and polycystic disease, share uniform apoptotic characteristics such as nuclear condensation, caspase activation, DNA fragmentation, guanosine triphosphate (GTP) depletion, mitochondrial dysfunction, and formation of reactive oxygen species (Ramesh and Reeves 2002). Since local IL-15 behaves as an anti-apoptotic and survival cytokine for multiple cell types (Fehniger and Caligiuri 2001), it may play a pivotal role in renal injury; however, thus far the experimental evidence is insufficient. Shinozaki and others (2002) demonstrated decreased intrarenal IL-15 levels in a model of nephrotoxic serum nephritis (NSN), and, by using IL-15−/− mice, IL-15 was shown to serve as a survival factor for kidney epithelial cells (Shinozaki and others 2002). In contrast, elevation of IL-15 mRNA levels was observed in renal transplant biopsies during acute cellular rejection (Strehlau and others 1997; Cookson and others 2003).
We have demonstrated in previous studies that IL-15 is produced by RECs in a steady-state manner and that it is up-regulated by IFN-γ (Tejman-Yarden and others 2005). Shortly, 7-min IL-15 stimulation of RECs induced tyrosine phosphorylation of the main downstream IL-15-signaling molecules Jak-1, Jak-3, STAT-3, and STAT-5 (Tejman-Yarden and others 2005). These data demonstrate the presence of a functional IL-15 receptor in the kidney. In the present study, we examined the involvement of IL-15 in three in vivo models of renal pathologies: sepsis-induced renal damage, IRI, and cisplatin-induced nephrotoxicity. We demonstrate, here, that kidney dysfunction correlates with reduced intrarenal IL-15 levels, and show that IL-15 exhibits anti-apoptotic properties in the kidney.
Materials and Methods
Mice
C57BL/6 mice (Harlan, Jerusalem, Israel) and IL-15Rα−/− mice (JAX Laboratories, Bar Harbor, ME) were housed in the animal laboratory of the Soroka Medical Center (Beer-Sheva, Israel). IL-15Rα−/− are of C57BL/6 background. The generation and characterization of IL-15Rα−/− mice has been previously described (Giri and others 1994). Ten- to twelve-week-old female mice were used in the sepsis and cisplatin models. In the IRI model, C57BL/6 male mice were used since female mice are reported to be more resistant to ischemic acute renal failure (ARF) than the males (Wei and others 2005). Animal use conformed to the guidelines established by the Animal Care Committee of our institution.
Sepsis model
Escherichia coli were grown in Luria-Bertani broth (Conda Laboratories, Madrid, Spain) and harvested during log phase. Bacteria containing 30% glycerol were stored at −70°C. Sepsis was induced in C57BL/6 mice by intraperitoneal (i.p.) inoculation of a sublethal dose of E. coli (1.5 × 107 colony-forming unit) as previously described (Mazar and others 2005; Rogachev and others 2006). Immediately after inoculation, or at 3.5, 6, and 24 h after inoculation, animals were sacrificed and left kidneys were harvested for total RNA extraction and for protein determination.
Ischemia-reperfusion injury model
C57BL/6 mice were anesthetized with sodium pentobarbital (i.p., 50 mg/kg) and maintained on a micropuncture heating table at 38°C throughout the experiment. An abdominal incision was made above the left kidney to expose the renal vessels. The renal artery was occluded with a vascular clamp for 30 min, and then removed to allow the organ to reperfuse. Immediately after reperfusion, or 8 and 24 h after reperfusion, the animals were sacrificed and both kidneys were harvested for total RNA extraction. For SHAM operation, the same procedure was carried out with the exclusion of clamping the renal artery.
Cisplatin nephrotoxicity model
C57BL/6 mice were injected i.p. with either saline or cisplatin (15 mg/kg, Pharmachemie BV-Teva Group, Haarlem, Holland). Immediately after injection, or at 24, 48, and 72 h after injection, animals were sacrificed and their left kidneys were harvested for total RNA extraction.
Preparation and stimulation of human and mouse RECs
Primary human RECs were obtained from normal cortex tissue of kidneys from patients with hypernephroma, as described previously (Weiler and others 1998). Primary mouse RECs were obtained from cortex tissue of C57BL/6 and IL-15Rα−/− mouse kidneys. Cells were grown in M199 medium containing 10% heat-inactivated fetal calf serum, 2 mM
Quantitative analysis of apoptosis
Stimulating mouse RECs was discontinued by trypsinization, and apoptosis was evaluated by the three following methods; for assessment of apoptosis by cell cycle assay, cells were centrifuged at 800g for 5 min and washed twice with 1 mL ice cold phosphate-buffered saline (PBS; Biological Industries). Cells were then fixed in cold ethanol and stored at −20°C. Sixteen hours later, cell pellet was washed twice with 1 mL PBS and incubated for 40 min with RNase and 0.1% Triton X-100 in PBS. Samples were incubated in the dark with 50 µg/mL propidium iodide (PI; Sigma, Rehovot, Israel) for 15 min. Analysis was performed by flow cytometer (FACSCalibur; Beckman Coulter, Inc., Fullerton, CA) equipped with an argon laser at 488 nm wavelength. A minimum of 10,000 cells were analyzed from each sample. Apoptotic cells were detected on a PI histogram as a sub-G1 hypodiploid area. Dead cells and debris were excluded. Data analysis was performed with Multicycle for Windows Software, Phoenix Flow System, Inc. Caspase-3 has been identified as a key mediator of apoptosis of mammalian cells. The presence of active caspase-3 was determined by using the fluorescein-labeled inhibitor FAM-DEVD-FMK, following the protocol of CaspaTag™ Caspase-3 In Situ Detection Kit (CHEMICON, Temecula, CA). TUNEL assay was performed using the Fluorescein FragEL™ DNA fragmentation detection kit (Calbiochem, Cambridge, MA) according to the manufacturer’s recommendations.
Quantitative mRNA analysis
Total RNA was extracted from human RECs or mouse renal tissue using RNeasy Mini Kit (Qiagen, Hilden, Germany). cDNA was prepared as previously described (Basok and others 2001). Quantitative real-time polymerase chain reaction (QPCR) assays were carried out for β-actin, IL-15, IL-15R subunits α, β, and γc chains, Bcl-2, and Bax with the use of specific primers (Table 1). cDNA templates (7 µL) were diluted 9-fold and added primers (0.2 mM) and Thermo-Start master mix (ABgene, Surrey, UK). Reactions were carried out in a Rotor-Gene Real-Time PCR apparatus (Corbett-Research, Northlake, Australia).
L
Abbreviation: IL, interleukin.
Western blot and ligand-binding analysis
Kidney tissue was homogenized in RIPA buffer (150 mM NaCl, 50 mM Tris–HCl, pH 7.4, 1% NP-40, 0.25% Na− deoxycholate, 1 mM ethylene glycol tetraacetate, proteinase inhibitor cocktail, and phosphatase inhibitor, Sigma). Twenty-five micrograms of protein were separated by SDS-PAGE and transferred to polyvinylidine difluoride membranes (Bio-Rad, Hercules, CA). The membranes were blocked for 1 h with TBST buffer (20 mM Tris–HCl, pH 7.5, 150 mM NaCl, 0.05% Tween-20) containing dry skim milk (5% w/v) or human serum albumin (4%) and then probed with primary antibodies: anti-IL-15Rβ (M-20; Santa Cruz Biotechnology, San Diego, CA), IL-15Rγ (M-20; Santa Cruz Biotechnology), anti-Bcl-2 (Oncogene, San Diego, CA), anti-Bax (Biosource International, Camarillo, CA), or 1 ng/mL rhIL-15 (R&D Systems, Minneapolis, MN) for 1 h at room temperature. Primary antibody reactions were detected using biotin-conjugated anti-human IL-15 (R&D Systems) and HRP–streptavidin (Zymed Laboratories, Inc., San Francisco, CA) or with other appropriate peroxidase-conjugated antibodies. Following analysis, membranes were stripped and reprobed with a specific primary anti-actin–mIgM antibody (Oncogene, San Diego, CA), followed by incubation with a secondary antibody. Visualization of specific proteins was conducted using Western Lightning™ chemiluminescence Reagent Plus (Renaissance; PerkinElmer, Boston, MA).
Statistical analysis
Results are expressed as means ± S.E. To compare levels between groups, one-way analysis of variance was used. P values of <0.05 were considered significant.
Results
IL-15 and IL-15R expression in common acute kidney injury
To investigate the importance of intrarenal IL-15 production and activity, we examined IL-15 and IL-15R expression in two renal injury models in vivo: sepsis-induced renal damage and renal IRI. Expression of intrarenal IL-15 and the three IL-15R subunits was evaluated by QPCR and by Western blot analysis at different time points after renal insult. As seen in Figure 1A, both IL-15 and IL-15Rα mRNA levels were elevated following E. coli inoculation, exhibiting a peak at 3.5 h. At this indicated time point, IL-15 mRNA levels were 2-fold higher than at time 0. The temporal increase was followed by a synchronized decline in IL-15 and IL-15Rα expression levels that persisted until IL-15 and IL-15Rα transcripts were reduced to one-fifth and two-third of their levels at time 0, respectively (Fig. 1A). As expected, IL-15R protein levels increased to a peak at 6 h (Fig. 1B). As seen in Figure 2, consistent with the sepsis model described earlier, IRI exhibited a significant decrease in intrarenal IL-15 mRNA levels, which were 11-fold less than SHAM-operated animals, as detected in ligated kidneys 24 h after reperfusion. Accordingly, ligated kidneys expressed 9-fold lower IL-15Rα mRNA levels compared to SHAM-operated animals 24 h after reperfusion (Fig. 2). In contrast, non-ligated kidneys from the same mice exhibited elevated IL-15 and IL-15R mRNA levels. Importantly, in the two examined pathologies, afflicted kidneys showed a significant reduction in IL-15 levels after 24 h.
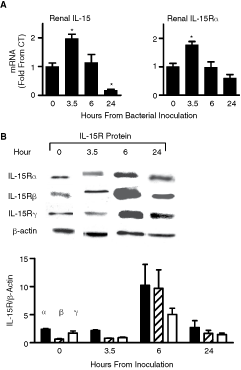
Interleukin (IL)-15 and IL-15R expression in sepsis. Renal injury during bacterial sepsis. Mice were injected with saline (CT) (n = 4) or inoculated with a sublethal dose of Escherichia coli (n = 4). Kidneys were harvested at indicated times after inoculation. (
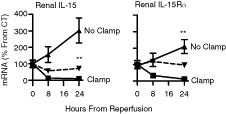
Interleukin (IL)-15 and IL-15R expression in ischemia-reperfusion injury (IRI). Renal damage during IRI. Ischemia of left renal artery was maintained for 30 min in all but SHAM-operated mice (n = 4). Left (clamp) and right (no clamp) kidneys were harvested at indicated times and QPCR was performed with primers specific for IL-15 and IL-15R. Dashed line, SHAM-operated mice (n = 4). CT, mRNA levels at 0 h. Representative experiment out of three is shown. Results: mean ± SE. **P < 0.01 between clamped and non-clamped kidneys.
Effects of IL-15 on Bcl-2 and Bax expression in the kidney
Local renal tissue apoptosis is a consistent finding for the kidney pathologies examined in the present study. Since IL-15 induces Bcl-2 in various T-cell lines and in NK cells (Cooper and others 2002; Berard and others 2003; Oh and others 2008), we examined steady-state Bcl-2 and Bax expression in kidneys of wild-type (WT) and IL-15Rα−/− mice. As shown in Figure 3A, IL-15Rα−/− mice expressed 2-fold greater Bax levels and 2-fold less Bcl-2 levels compared to WT mice. To investigate whether exogenous IL-15 can affect Bcl-2/Bax ratio, cultures of primary human RECs were added rIL-15 (1 ng/mL). As shown in Figure 3B, this resulted in the reversal of the molecular ratio and a significant 4.6-fold increase in the Bcl-2/Bax ratio.

Effects of interleukin (IL)-15 on Bcl-2 and Bax expression in the kidney. (
Anti-apoptotic effect of IL-15 on primary renal cells
It is possible that IL-15 plays a role in setting a higher apoptotic threshold for renal cells by elevating the Bcl-2/Bax ratio. To examine whether IL-15 has a protective effect on kidney cells during a toxic challenge, primary cultures of RECs isolated from WT or IL-15Rα−/− mice were exposed to cisplatin. Apoptosis was evaluated by three standard apoptosis assays that examine early and late apoptosis. In accordance with the decreased Bcl-2/Bax ratio found in vivo in kidneys of IL-15Rα−/− mice, IL-15Rα−/− primary renal cells were more sensitive to cisplatin-induced apoptosis compared to WT cells (Fig. 4A). While the addition of the toxin cisplatin to cultures produced a 2.3-fold increase in the apoptotic rate in normal cells, it produced a 3.8-fold increase in IL-15Rα−/− cells, as determined by cell cycle analysis (Fig. 4A). Furthermore, as shown in Figure 4B, addition of exogenous rIL-15 to normal renal cells reversed cisplatin-induced apoptosis. RECs that were added rIL-15 exhibited a decrease of 33.9%, 31.6%, and 89.4% in the apoptotic rate compared to control cells, as determined by cell cycle analysis, as well as the presence of active caspase-3, and by terminal deoxynucleotidyl transferase (TdT)-mediated nick end-labeling (TUNEL) staining, respectively.
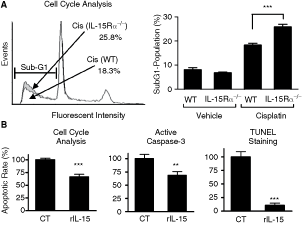
Anti-apoptotic effect of interleukin (IL)-15 on renal cells. (
Association between IL-15 and renal function during cisplatin-induced nephrotoxicity
To further investigate the role of intrarenal IL-15 in maintaining kidney integrity and function, we examined IL-15 and IL-15R expression in an in vivo model of cisplatin-induced nephrotoxicity (Fig. 5). A dramatic decrease of IL-15 mRNA (23-fold) and IL-15Rα mRNA (12-fold) levels was observed 24 h after toxin injection. Notably, 48 and 72 h after cisplatin injection IL-15 and IL-15Rα levels remained lower than basal levels (Fig. 5A). Renal dysfunction was assessed by determining BUN levels by which a positive correlation between renal dysfunction and low IL-15 levels was observed (Fig. 5B). These findings suggest that the protective role of IL-15, when expressed at higher levels, is a common feature of various kidney insults. In support of the functionality of IL-15 during these changes, a striking coordinated expression between IL-15 and its critical IL-15R was observed in all three models examined in this study.
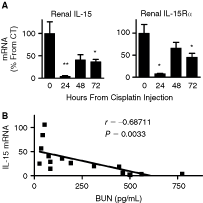
Association between interleukin (IL)-15 and renal function during cisplatin-induced nephrotoxicity. Renal damage during cisplatin-induced toxicity. Mice were injected with either saline (CT) (n = 5) or 15 mg/kg cisplatin (n = 5). Kidneys were harvested at indicated times and QPCR was performed. (
Discussion
In previous studies we have shown that IL-15 is produced by renal cells and that the three IL-15R subunits (α, β, γ) are expressed constitutively in cortex and medulla of murine kidneys and in primary cultures of human RECs (Weiler and others 1998; Tejman-Yarden and others 2005). In the present study, the synchronous kinetics of intrarenal IL-15 and intrarenal IL-15Rα expression was evaluated in three animal models that represent important human renal pathologies: sepsis-induced renal damage, renal IRI, and cisplatin-induced nephrotoxicity.
Most cytokines are not constitutively expressed in the kidney, but are rather up-regulated during renal disease (Yoshioka and others 1993; Hribova and others 2007). Indeed, the role of inflammation has been documented in acute renal injury and involves rapid cytokine-mediated activation of neutrophils, mononuclear cells, and endothelial cells (Jain and others 2000; Jo and others 2002). The induction of different cytokines and the increased expression of adhesive molecules by endothelial cells and phagocyte cells ultimately converge to mediate tissue injury (Willinger and others 1992). It appears, therefore, that the interplay between the kidney-infiltrating leukocytes and resident RECs can control the amplitude of local inflammation. Interestingly, while the expression of IL-15 is constitutive in normal RECs, our study indicates that it is significantly reduced in the afflicted kidneys after a 24-h period. Our findings are supported by the recent study of Shinozaki and others that showed decreased IL-15 expression in mouse kidneys after induction of NSN (Shinozaki and others 2002).
In the sepsis model, a temporal increase in IL-15 and IL-15Rα mRNA levels was observed several hours after E. coli inoculation. All three subunits of IL-15R protein reached a peak at 6 h, in agreement with increased levels of IL-15Rα transcripts. In our previous work, stimulation of cultured RECs with rIFN-γ induced an increase in mRNA and protein levels of IL-15 and its specific IL-15Rα chain, but did not affect the expression of IL-15Rβ and IL-15Rγ subunits (Tejman-Yarden and others 2005). Since the promoter of IL-15 and IL-15Rα genes contains an IFN-γ regulatory factor response element (IRF-E), it is possible that the inducible effect exerted on both IL-15 and IL-15Rα is caused by a rise of IFN-γ in the inflammatory environment (Mariner and others 2002). The increase in IL-15Rβ and IL-15Rγ protein levels that we found in vivo, therefore, may be partly attributed to infiltrating IL-15Rβγ-bearing leukocytes.
Consistent across all three models, changes in intrarenal IL-15 were closely linked to changes in IL-15Rα, the expression of which appears to pose a limiting factor for IL-15 activity. Recent studies have shown that IL-15 is presented in trans to neighboring target cells that express high affinity (IL-15Rαβγc) or intermediate affinity (IL-15Rβγc) complexes (Bulfone-Paus and others 2006). Substantial in vitro and in vivo experimental evidence indicates that membrane- associated IL-15 expressed on dendritic cells and monocytes can support the growth and proliferation of adjacent NK and CD8+ T cells (Sandau and others 2004), which express the partial IL-15R heterodimer (IL-2Rβγc). The striking coordinated expression of IL-15 and IL-15Rα observed in all three renal pathologies suggests that transpresentation of IL-15 may represent a mechanism for a localized cytokine function in renal tissue.
Notably, at 24 h after induction of injury, the decrease in IL-15 and IL-15Rα coincided with the appearance of apoptotic damage, a process that typifies the three examined pathologies (Arany 2008). Indeed, in the nephrotoxicity model, we found a significant correlation between reduced kidney function and reduced IL-15 levels. Our findings therefore support a protective role for IL-15 during kidney insult. According to these data, it is possible that during stress or pathological states, both IL-15 and IL-15Rα are expressed by kidney cells, which then bind IL-15 to IL-15Rα on cell membranes and transpresent the survival cytokine to adjacent renal cells, thereby promoting cell viability during lethal conditions.
Recent evidence suggests that REC apoptosis can be triggered either by the death receptor-signaling pathway, or by the intrinsic pathway, which is mediated by members of the Bcl-2 protein family (Shimizu and Yamanaka 1993; Takeda and others 1997). Our results indicate that IL-15 and IL-15Rα regulate the expression of Bcl-2 and Bax in RECs. We show that kidneys of IL-15Rα−/− mice express lower Bcl-2 and higher Bax mRNA and protein levels, compared to WT mice. Indeed, WT cells that were introduced with rIL-15 produced an elevated Bcl-2/Bax ratio, contrary to that observed in IL-15Rα−/− mice. These findings are supported by the study of Wu and others (2002), which demonstrated a failure to up-regulate Bcl-2 expression by memory CD8+ T cells deficient in IL-15Rα, compared with naïve CD8+ cells. Also, rIL-15 both up-regulated the level of Bcl-2 and reduced lethality of activated WT and mutant CD8+ T cells in vitro (Wu and others 2002). Similarly, in several works, IL-15 was shown to increase the expression of anti-apoptotic Bcl-2 and Bcl-xL in CD8+ T and fibroblast cells (Kurowska and others 2002; Berard and others 2003). Furthermore, it has been recently demonstrated that dendritic cells co-incubated with IL-15 express high levels of Bcl-2 and display an increased resistance to apoptosis (Tourkova and others 2002). IL-15 was shown to be critical for the maintenance of NK cells by up-regulating Bcl-2 (Minagawa and others 2002). Several studies reported that Bcl-2 gene promoter contains a STAT-5 consensus element, and that STAT-5 is able to induce Bcl-2 expression (Fang and others 2008). We previously showed that engagement of IL-15 with IL-15R induces Jak-1/Jak-3 and STAT-3/STAT-5 tyrosine phosphorylation in kidney cells (Tejman-Yarden and others 2005). Thus, the Bcl-2 down-regulation we found in renal cells of IL-15Rα−/− mice may be the result of reduced Jak/STAT-5 activation. In our present study, in concordance with the decreased Bcl-2/Bax ratio observed in kidneys of IL-15Rα−/− mice, IL-15Rα−/− primary renal cells were significantly more sensitive to cisplatin-induced apoptosis. Furthermore, according to in vitro assays using WT REC, IL-15 decreased the rate of cisplatin-induced apoptosis.
The protective effects of IL-15 were reproducible in three independent apoptosis assays, which are indicative of both early and late apoptosis. For example, pretreatment of RECs with rIL-15 prior to cisplatin addition exhibited a substantial decrease in the activated form of caspase-3, compared to untreated cells. This is in line with previous works (Bouchard and others 2004; Obermeier and others 2006) that have reported the reduced presence of active caspase-3 in neutrophils and intestinal epithelial cells following IL-15 induction. In the present study, addition of IL-15 to cultured RECs was similarly found to reduce sub-G1 population and to yield a significant decrease in DNA fragmentation. These observations are in accordance with the previous study of Shinozaki and others (2002) and suggest that IL-15 is critical for the survival of REC during pathological conditions.
In conclusion, both IL-15 and its receptor are likely to have an important role in both normal and pathological conditions of the kidney. However, the molecular mechanisms that modulate intrarenal IL-15/IL-15R levels in kidney injuries are unclear. While our findings indicate that the survival functions of IL-15 in the kidney are mediated by members of the Bcl-2 family and involve caspase-3 regulation, additional factors are most probably involved. The protection against apoptosis by IL-15 is particularly important in renal cells since this cell population is continuously exposed to toxic inducers of apoptosis and to extreme and variable pH conditions. There is no direct evidence to show a protective role for IL-15 on human renal cells. However, a significant homology exists between mouse and human IL-15 mRNA (73% identity) (Grabstein and others 1994), and in addition, Fehniger et al. confirmed that IL-15 expression is similarly controlled in mice and humans at the levels of transcription, translation, posttranslational processing, intercellular trafficking, and translocation (Fehniger and Caligiuri 2001). Thus, these observations suggest that IL-15 has a similar effect on human kidneys as is seen in our mouse model.
A better understanding of the role and regulation of renal IL-15 may provide new insights into the pathogenesis of renal failure and offer a basis for new therapeutic strategies.
Footnotes
Acknowledgments
This work was supported by the Dr. Montague Robin Fleisher Kidney Transplant Unit Fund. We would like to thank Valeria Frishman for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.