
Abstract
Chronic alcohol consumption has been shown to decrease the activity of natural killer (NK) cell cytolytic function and the production of various cytokines from the spleen. We have recently shown that naltrexone, an opiate receptor antagonist, when administered for a period of 2 weeks suppresses µ-opiate receptor binding but increases ∂-opiate receptor activity in rat splenocytes. However, the effects of long-term naltrexone treatment on alcohol-induced alteration of NK cell cytolytic activity and cytokines production in splenocytes have not been determined. Male rats were pair-fed an isocaloric liquid diet or fed an ethanol-containing liquid diet for a period of 3 weeks. These rats were additionally treated after a week with a subcutaneous implant of either a naltrexone pellet or placebo pellet for 2 weeks. Splenocytes were isolated and used for determination of various cytokines interleukin (IL)-2, IL-4, and IL-6, and interferon-γ (IFN-γ) using enzyme-linked immunosorbent assay (ELISA), and the basal and IL-2-, IL-12-, or IL-18-induced NK cytolytic activity was measured using a standard 4-h 51Cr release assay against YAC-1 lymphoma target cells. Ethanol consumption resulted in a reduction of the production of IL-2, IL-4, and IL-6 as well as the basal and cytokine-activated NK cell cytolytic activity and IFN-γ production in splenocytes. Naltrexone administration increased the production of IL-2, IL-4, and IL-6 and the basal and cytokine-activated NK cell cytolytic activity and IFN-γ production in the splenocytes of pair-fed and alcohol-fed rats. These results indicated that naltrexone treatment increases NK cell cytolytic activity and cytokine production in the spleen in vivo. Furthermore, these results identify the potential of the use of naltrexone in the treatment of immune deficiency in alcoholic and non-alcoholic patients.
Introduction
E
NK cell cytolytic activity has been shown to be activated by interferon-γ (IFN-γ), which has a number of opioid-like effects (Santoli and Koprowski 1979; Reiter 1993). Various other cytokines are also known to increase NK cell cytolytic activity and lymphocyte proliferation. Of these cytokines, interleukin (IL)-2, IL-12, and IL-18 stimulate NK cell cytolytic activity (Otraldo and others 1983; Wysocka and others 1995; Trinchieri and others 1996; Atkins and others 1997; Tomura and others 1998; Thornton and others 2001; Trinchieri 1998). Other cytokines like IL-4 and IL-6 are known to regulate NK cell proliferation and differentiation (Jansen and others 1990; Hayakawa and others 1991; Bonavida and others 1993; Trinchieri and others 1993). Cytokines IFN-γ, IL-2, IL-4, IL-6, IL-12, and IL-18 have been shown to also affect the function of other immune cell populations in splenocytes (Van der Meide PH, Schellekens H. 1996; Dunn 1991). We have previously shown that chronic alcohol administration suppresses NK cell cytolytic activity as well as IFN-γ production from splenocytes in rats (Boyadjieva and others 2001, 2002). Alcohol consumption is also known to decrease concanavalin A (ConA)-induced production of various cytokines, including IL-2, IL-6, and IL-4 from splenocytes (Wang and others 1993). Whether naltrexone administration prevents alcohol-altered production of NK cell cytolytic activity and cytokine production in splenocytes is not known. In order to better understand naltrexone actions on the immune system, we determined this µ-opiate receptor blocker on NK cell cytolytic activity and the production of NK cell regulatory cytokines in the spleen of male rats in vivo.
Materials and Methods
Animals and feeding design
Male Fischer-344 rats of 150–175 g body weight were maintained on a 12-h light/dark cycle (lights on 0700 h and lights off 1,900 h). Animals were either fed an ethanol-containing liquid diet (8.7% v/v) or pair-fed an isocaloric liquid diet (Bio-Serv, Frenchtown, NJ), and were given 30 graduated ball-barrel cylinders containing the freshly prepared diet daily, 1 h before lights off at 19:00 h. Animals were also given free choice of water. In this study, we have not included ad lib chow-fed animals to identify any nutritional influence of liquid diet feeding, since we have previously identify no significant effect of liquid diet feeding procedure on immune functions in Fischer rats (Boyadjieva and others 2001, 2002; Dokur and others 2001). After the first week of adjustment to the diet, both pair-fed and ethanol-fed rats were subcutaneously implanted with a naltrexone pellet or with a placebo pellet (50 mg/pellet, 21-day release; Innovative Research of America, Sarasota, FL). The naltrexone treatment as well as alcohol and pair-feeding continued for additional 2 weeks. As we have shown previously (Boyadjieva and others 2004), the diet consumption of the ethanol-fed animals, both treated and untreated, was similar (∼70–95 mL/day). The blood alcohol concentration at the last day of treatment in rats treated with ethanol and naltrexone was not different (0.12 ± 0.01 g/dL; n = 7) from that of rats treated with ethanol and placebo (0.13 ± 0.01 g/dL; n = 8). At the end of the experiments, animals were weighed and decapitated. Spleen tissues were immediately removed, weighed, and used for isolation of lymphocytes and for studies on NK cell activity and cytokine production. Animal surgery and care were performed in accordance with institutional guidelines and complied with NIH policy. The animal protocol was approved by the Rutgers Animal Care and Facilities Committee.
NK cell cytolytic activity
NK cell activity was determined using a standard 4-h 51Cr release assay with YAC-1 lymphoma cells as targets. At the end of the experiments, spleens from treated rats were taken, and splenocytes were isolated as described by us previously (Boyadjieva and others 2001). Erythrocytes were removed by a 5-s hypotonic shock with sterile distilled water. Cells were cultured for NK cell activity assay for 12 h in the presence or absence of rIL-2 (500 U/mL), rIL-12 (10 ng/mL), or rIL-18 (50 ng/mL; all from Sigma, St Louis, MO). The percentages of cytolytic activity at effector:target ratios of 200:1, 100:1, 50:1, and 25:1 were converted to lytic units per 106 effector cells according to Pross and others (1981). Each assay was conducted in quadruplicate and the cytolytic activity of the individual effector:target ratios used in the conversion. NK activity is expressed in lytic units.
Determination of cytokine levels (IL-2, IL-4, IL-6, and IFN-γ) in supernates of cultured splenocytes
Splenocytes of alcohol-, naltrexone-, or control treated animal were used for determination of IFN-γ, IL-2, IL-4, and IL-6 production. For IFN-γ determination, splenocytes cells were cultured in 24 wells (500,000 cell/well) in RPMI 1640 medium containing glucosamine in the presence or absence of rIL-2 (500 U/mL), rIL-12 (10 ng/mL), or rIL-18 (50 ng/mL, St Louis, MO) for a period of 24 h. For determination of IL-2, IL-4, and IL-6 production from splenocytes, cells were cultured in 24 wells (500,000 cell/well) in RPMI 1640 medium, containing glucosamine and concanavaline A (Con A; 5 µg/mL) for a period of 24 h. The supernatant was removed and used for the determination of cytokines. The rat IL-2, IL-4, IL-6, and IFN-γ ELISA kits (Ouantikine M, Murine; R&D Systems, Inc, Minneapolis, MN) were used for the quantitative determination of rat cytokines concentrations in picograms per milliliter in cell culture media.
Statistics
The mean and standard error of mean (± SEM) of the data were determined and are presented in the text and figures. Data were analyzed using ANOVA. The differences between groups were determined using the Student Newman-Keuls test. A value of P < 0.05 was considered a significant difference.
Results
Effects of naltrexone administration on NK cell cytolytic activity in splenocytes of alcohol-fed and control-fed rats
We have previously shown that alcohol administration for 2 or 4 weeks decreases the NK cell cytolytic activity (Boyadjieva and others 2001). In agreement with these findings, data shown in Figure 1A indicate that ethanol intake for 3 weeks reduced the NK cell activity as compared to pair-fed animals (ethanol-treated rats had 71.5% lower value than controls). Additionally, we show that naltrexone administration for 2 weeks during the period of ethanol intake antagonized the suppressive effects of alcohol on NK cell cytolytic activity (naltrexone rats had 405% higher values than placebo rats). Naltrexone also increased NK cell cytolytic function in pair-fed control rat's splenocytes (naltrexone rats had 157% higher values than placebo rats). Determination of ethanol effect on the cytokine-stimulated NK cell cytolytic activity revealed that ethanol markedly inhibits IL-2-, IL-12-, or IL-18-activated NK cell cytolytic function (ethanol-treated rats had values 14.4%, 14.3%, and 12.4% of placebo rats following IL-2, IL-12, and IL-18 treatment, respectively; Fig. 1B–1D). Naltrexone administration was able to increase IL-2-, IL-12-, and IL-18-activated NK cell cytolytic function in both alcohol-fed (naltrexone rats had 731%, 509%, and 398% higher values than placebo rats following IL-2, IL-12, and IL-18 treatment, respectively) and pair-fed animals (naltrexone rats had 216%, 134%, and 129% higher values than placebo rats following IL-2, IL-12, and IL-18 treatment, respectively).
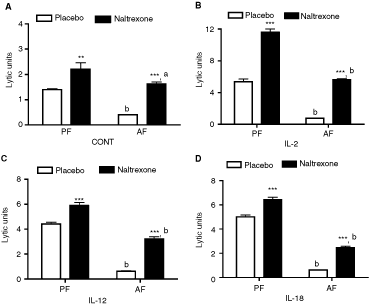
Effects of chronic ethanol consumption with or without naltrexone on (
Effects of naltrexone administration on IFN-γ production from splenocytes of alcohol-fed and control-fed rats
We have previously shown that alcohol administration for 2 weeks decreases the production of IFN-γ from splenocytes (Dokur and others 2001). In agreement with the previous findings, we show here ethanol feeding reduces the splenocytes production of IFN-γ (ethanol-treated rats had 14% of control values; Fig. 2A). In addition, ethanol feeding reduces the ability of splenocytes to produce IFN-γ in response to IL-2-, IL-12-, and IL-18 (ethanol-treated rats had 28.4%, 21.1%, and 23.4% of control values following IL-2, IL-12, and IL-18 treatment, respectively; Fig. 2B–2D). Naltrexone administration for 2 weeks during the period of ethanol intake antagonized the suppressive effects of alcohol on IFN-γ in splenocytes (naltrexone rats had 537% higher values than placebo rats). Naltrexone increased IFN-γ levels in pair-fed control rat's splenocytes (naltrexone rats had 177% higher values than placebo rats). Naltrexone administration was also able to increase IL-2-, IL-12-, and IL-18-activated production of IFN-γ in both alcohol-fed (naltrexone rats had 403%, 775%, and 449% higher values than placebo rats following IL-2, IL-12, and IL-18 treatment, respectively) and pair-fed animals (naltrexone rats had 123%, 158%, and 144% higher values than placebo rats following IL-2, IL-12, and IL-18 treatment, respectively).

Effects of chronic ethanol consumption with or without naltrexone on (
Effects of naltrexone administration on IL-2, IL-4, and IL-6 production from splenocytes of alcohol-fed and control-fed rats
Alcohol consumption is known to decrease the splenic production of IL-2, IL-6, and IL-4 in response to immune challenge in mice (Wang and others 1993). We also show here chronic ethanol consumption reduced the ConA-induced production of IL-2, IL-4, and IL-6 from the spleen of rats (ethanol-treated rats had 44.0%, 61.0%, and 40.7% of control IL-2, IL-4, and IL-6 values, respectively; Fig. 3A–3C). Furthermore, naltrexone treatment was able to increase the production of these cytokines in splenocytes of both alcohol-fed rats (changes in IL-2, IL-6, and IL-4 levels were 1,143%, 356%, and 645% of controls, respectively) and pair-fed animals (changes in IL-2, IL-6, and IL-4 levels were 666%, 261%, and 500% of controls, respectively).

Effect of chronic ethanol consumption with or without naltrexone on (
Discussion
The data presented here demonstrated that ethanol consumption suppressed basal and IL-2-, IL-12-, and IL-18-induced NK cell cytolytic activity and INF-γ production from splenocytes as well as reduced the ability of splenocytes to produce IL-2, IL-4, and IL-6 in response to ConA. The studies also provided evidence that chronic administration of naltrexone stimulates the production of cytokines and NK cell cytolytic activity in splenocytes. Additionally, the data demonstrate that naltrexone is able to antagonize ethanol's suppressive effects on NK cell cytolytic activity and splenocytes’ cytokine production.
NK cells are populations of lymphocytes that can be activated to mediate significant levels of cytotoxic activity and to produce high levels of certain cytokines and chemokines (Perussia 1996). In addition to NK cells, T cells and macrophages are known to produce many cytokines upon stimulation by mitogen in the spleen (Van der Meide and Schellekens 1996). The biological consequences of the production of specific cytokines are related to the cytokine's effect on the immune cells themselves. These cytokines not only alter cytolytic activity, but also affect immune cell proliferation (Biron and others 1999). Our data demonstrated that IL-2, IL-12, and IL-18 stimulated splenic NK cell cytolytic activity, and that chronic ethanol consumption decreased the NK cell response to these cytokines. These data are consistent with our previous findings that ethanol consumption decreases basal NK cell cytolytic activity, and granzyme B and IFN-γ production in the spleen levels (Boyadjieva and others 2001; Dokur and others 2001). The present data showing ethanol's suppression of IL-2-induced NK cell cytolytic activity in rats are also in agreement with the previous finding that ethanol consumption suppresses IL-2-induced cytolytic activity possibly via decreasing NF-κB and AP1-binding activity in murine splenic NK cells (Zhou and Meadows 2003). Like IL-2, both IL-12 and IL-18 have been shown to induce NK cell activity and IFN-γ production (Walker and others 1999; Brunda and others 1993). Previously, it has been shown that IL-12 and IL-18 are the most efficient inducers of NK cell activity and IFN-γ production (Gately and others 1992, 1994; Biron and others 1999; Walker and others 1999; Gately and others 1998; Nishimura and others 1996; Sato and others 2002). In our study we also found that both IL-12 and IL-18 increase NK cell cytolytic activity and IFN-γ production. We also demonstrated here that chronic ethanol consumption depresses the ability of these cytokines to enhance NK cell activity and IFN-γ production from splenocytes. Other cytokines, like IL-4 and IL-6 as well as IL-2, play a role in NK cell proliferation and differentiation (Jansen and others 1990; Bonavida and others 1993; Trinchieri and others 1993). We showed here that the production of these cytokines was decreased by ethanol treatments. We have previously demonstrated that chronic alcohol consumption decreased the mitogen-stimulated proliferation of lymphocytes, and the number and percentage of various lymphocyte subsets including NK cells in the spleen (Boyadjieva and others 2002). Together these data suggest that ethanol not only suppresses the NK cell production of cytotoxic factors like granzyme B and INF-γ critical for cytolytic activity but also the splenic production and action of various cytokines critical for NK cell and other immune cells proliferation and function.
Our present studies demonstrated that chronic naltrexone administration enhanced both basal and the cytokine-modulated NK cell cytolytic activity and IFN-γ production in both pair-fed and ethanol-fed rats. Naltrexone also increased spenocyte production of IL-2, IL-4, IL-6. Naltrexone is an opioid antagonist when administered acutely, but shows δ-opioid-like activity following chronic administration (Boyadjieva and others 2001, 2004). Previously, it has been shown that opioid peptides play a significant regulatory role in controlling NK cell cytolytic activity. It has been shown that chronic treatment with ethanol reduces central and peripheral levels of BEP, and that the reduction of hypothalamic and plasma β-endorphin (BEP), following chronic ethanol intake, correlates with the reduction of NK cell cytolytic activity. Furthermore, hypothalamic administration of BEP raises the reduced NK cell cytolytic activity to a normal level in ethanol-treated animals (Boyadjieva and others 2001). Previous studies also demonstrated opioid's stimulatory effect on cytokine-like IL-6 production following peripheral and central routes of administrations (Bertolucci and others 1996; Zubelewicz and others 1999). Since BEP is known to reduce stress axis function (Plotsky 1986), the question arose whether naltrexone action of immune cells is secondary to its action on corticosterone. Treatment of naltrexone in mice showed no effect on basal levels of plasma corticosterone (Wang and others 2002). Furthermore, naltrexone increased cytokine and NK cell cytolytic function in both pair-fed and alcohol-fed rats in this study. Hence, it is unlikely that naltrexone increased cytokines production from spleen via suppressing corticosterone levels. These data support that chronic naltrexone produce an opioid-like activity in cytokine production and cytokine actions on NK and other immune cells.
We have recently shown that naltrexone preferentially prevented µ-opioid receptor activity but increased δ-opioid receptor activity following chronic administration (Boyadjieva and others 2004). The δ-opioid receptor stimulatory action of chronic naltrexone in the spleen is consistent with the observation that chronic naltrexone increases the δ-opioid receptor-mediated antinociception response as well as the levels of δ-receptor binding in brain tissues (Kest and others 1998). Hence, it appears that δ-opioid receptor up-regulation may potentiate cytokines production and activation of NK cell function. However, further study is needed to determine changes in opioid receptor expression and binding activity in the spleen after ethanol and cytokine treatments in order to evaluate the role of specific opioid receptors in the mediation of ethanol's action on cytokine-regulated NK cell function.
Footnotes
Acknowledgment
This investigation was supported by National Institutes of Health Grant AA016296.