
Abstract
Biological membranes are described as a mosaic of different domains where interactions between membrane components induce the formation of subdomains with different characteristics and functions. Lipids play an important role in the formation of lipid-enriched microdomains where they dynamically associate to form platforms important for membrane protein sorting and construction of signaling complexes. Cholesterol confined in lipid domains is a crucial component required by microorganisms, directly or indirectly, to enter or exit the intracellular compartment. Cellular activation mediated by superficial bacterial component may be modified by local cholesterol depletion. Therefore, new perspectives for unconventional therapeutic intervention in Gram-negative infections may be envisaged. We tested this hypothesis by using methyl-β-cyclodextrin (mβCD) as a cholesterol-complexing agent to alter the U937 plasma membrane cholesterol content. Our results demonstrate that cholesterol depletion of U937 cells inhibited Salmonella enterica serovar Typhimurium porins-mediated phosphorylation of Src kinase family, protein kinase C (PKC), JNK, and p38, while cholesterol repletion restored the phosphorylation. Lipopolysaccharide (LPS) extracted from the same bacterial strain has been used as a control. Our data demonstrate that the lack of activation of signal transduction pathway observed following cholesterol depletion differently modulates the release of interleukin-6 (IL-6) or tumor necrosis factor-α (TNF-α), suggesting that Src, associated to lipid domains, may represent an important pathway in Gram-negative-induced cellular signal.
Introduction
T
At the moment eight kinases belonging to the Src family are known (Src, Lyn, Fyn, Yes, HcK, Fgr, LcK, BlK) (Roskoski 2004), and all present a common domain and differently from other NT-PTKs they are able to bind the cytoplasmic membrane. The coupling occurs within specialized regions in the plasma membrane, enriched in sphingolipids and cholesterol. Membrane structures with this composition are very resistant to solubilization by nonionic detergents such as Triton X-100 (TX-100). These isolated detergent-resistant membranes (DRMs) are enriched in cholesterol, sphingomyelin, glycosphingolipids, and saturated glycerophospholipids, as well as dually acylated Src family kinases and glycosylphosphatidylinositol (GPI)-anchored proteins (Wollscheid and others 2004). These plasma membrane domains may function as centers for signal transduction and membrane trafficking (Hanzal-Bayer and Hancock 2007).
The LPS-mediated signaling takes place through Toll receptors, and moreover LPS binds to CD14 on lipid-enriched low-density domains of the human monocyte–macrophage THP-1 cells plasma membrane. LPS most likely interacts with DRMs via its GPI-anchored receptor CD14 and induces MAPK activation and cytokine production, probably because aggregates of LPS cross-link CD14 molecules leading to the formation of DRM-signaling patches (Olsson and Sundler 2006). Motoyama and others (Motoyama and others 2005) describe the inhibitory effects of cyclodextrin on LPS-induced cytokine production in macrophages. In the present report LPS has been used as a positive control.
Whole bacteria might also trigger DRMs in order to parasite host-signaling cascades. This might be the case of adhering Escherichia coli (DAEC), which induces microvilli injury and actin disassembly in intestinal cells. Actin disassembly requires the GPI-anchored protein DAF/CD55 (Kansau and others 2004). DRMs play an important role in human immunodeficiency virus infection of CD4+ T cells (Popik and others 2002) in Mycobacterium tuberculosis infection of macrophages (Gatfield and Pieters 2000) in cholera toxin (Lencer 2001) and vacuolating cytotoxin A (Patel and others 2002). Mycobacteria are also capable of interacting with the DRM-cholesterol (Pieters 2001). Cholesterol-rich domains are also targeted by Chlamydia (Jutras and others 2003), and by Helicobacter pylori (Hessey and others 1990). Moreover, the early contact of Shigella flexneri with epithelial cells is initiated within DRMs (Lafont and others 2002). GPI-anchored proteins, cholesterol, or unidentified components of lipid domains are utilized by several viruses as their cellular receptors. GPI-anchored DAF (CD55), a member of the complement regulatory proteins, is targeted by at least six echovirus serotypes as well as by coxsackievirus A21, B1, B3, and B5 (Stuart and others 2002; Milstone and others 2005; Pettigrew and others 2006). Cholesterol is required to maintain the functionality of lipid domains and integral signaling capacity of plasma membrane.
We previously demonstrated that porins are able to activate NT-PTKs in U937 cells (Galdiero and others 2003). In other studies it has been shown that several components required to activate the MAPK pathway are enriched in cholesterol-rich microdomains (Simons and Toomre 2000; Cheon and others 2008).
In this study, we investigated the hypothesis that cholesterol-rich domains carry out an important role for cytokines release following a bacterial stimulus and through the activation of NT-PTK kinases. We demonstrated that cholesterol is required for signaling the activation of Src, Protein kinase C (PKC), p38, JNK, leading to the tumor necrosis factor-α (TNF-α) release by porins or LPS in U937 cells. We have used methyl-β-cyclodextrin (mβCD) to selectively deplete cholesterol from U937 cell plasma membrane.
Materials and Methods
Cell lines
U937 monocytes (ATCC CRL-1593.2) were grown at 37°C in 5% CO2 in RPMI 1640 supplemented with 10% heat-inactivated FCS, glutamine (2 mM), penicillin (100 U/mL), and streptomycin (100 U/mL) and differentiated as previously described (Galdiero and others 2005).
Cellular cholesterol depletion and repletion
Cells (3 × 106/mL) undergoing cholesterol depletion were washed with phosphate buffer saline (PBS) and then treated with mβCD (10 mM) in serum-free bovine serum albumin (BSA)/RPMI 1640 for 30 min at 37°C and 5% CO2. For cholesterol repletion experiments, cholesterol (5 μg/mL) was added back to cholesterol-depleted cells by incubation for 30 min at 37°C 5% CO2 in serum-free BSA/RPMI 1640. These treatment were effected similarly to a previously described procedure (Sheets and others 1999). All depletion agents were diluted in RPMI 1640 containing fatty acid-free BSA (10 μg/mL).
Lipid extractions and analyses
U937 cells were depleted of cholesterol (or not) and repleted (or not) as described earlier. After treatment, the cells were washed with RPMI/BSA and then resuspended in PBS. Cells were washed three times with PBS and extracted with hexane/propan-2-ol (3:2, v/v). The lipid extracts were evaporated under a stream of nitrogen and resuspended in propan-2-ol for total cholesterol determination by an enzyme assay kit from BioVision (BioVision Research, USA).
Preparation of porins and LPS
Porins were isolated from the lysozyme–EDTA extracted envelopes of Salmonella enterica serovar Typhimurium strain SH5014 as previously reported (Galdiero and others 2002). The protein content of the porins preparation was determined by the method of Lowry and others (1951) and checked by SDS-PAGE according to Laemmli (1970). SDS-PAGE revealed two bands with molecular weights of 34 and 36 kDa, confirming the purity of the preparation. All possible traces of LPS were revealed on SDS-PAGE stained with silver nitrate as described by Tsai and Frasch (1982) and by the Limulus-amoebocyte-lysate assay (Limulus test, pbi international, Milan Italy) according to Yin and others (1972). LPS was isolated using the phenol/chloroform/ether method described previously (Galdiero and others 2002). Purity of the porins and LPS preparations from S. enterica serovar Typhimurium were reported in a previous paper (Galdiero and others 2002).
Cell stimulation and preparation of cell lysates
U937 cells (3 × 106/mL) depleted of cholesterol (or not) and repleted (or not), suspended in serum-free BSA/RPMI, were stimulated with different concentrations (1, 5, and 10 μg/mL) of stimuli for different time periods (10, 15, 30, and 60 min). After incubation, the cells were prepared as previously reported (Galdiero and others 2002) and used for enhanced chemiluminescence’s (ECL) Western Blot analysis.
Kinase phosphorylation
Cell lysates were immunoprecipitated and used for Western blotting. Immunoprecipitation was carried out with the appropriate antibodies: anti-phospho-PKC antibody recognize an short amino acid sequence containing phosphorylated Ser657 of PKC-α of human origin; anti phospho-p38 antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA), which is a rabbit polyclonal antibody raised against a peptide mapping at the amino terminus of p38 of mouse origin identical to the corresponding human sequence and is directed against Thr180 and Tyr182-phosphorylated p38 (New England Biolabs, Ipswich, MA); anti-phospho-JNK antibody, which is a mouse monoclonal IgG1 antibody raised against a peptide corresponding to a short amino acid sequence phosphorylated on Thr183 and Tyr185 of JNK of human origin (Santa Cruz Biotechnology, Inc.); and anti-phospho-Src family antibody recognize a synthetic phosphopeptide corresponding to residues surrounding Tyr416 of human Src (Cell Signaling technology, Inc., Danvers, MA). Blots were blocked for 1 h at room temperature in Tris-buffered saline (TBS [150 mM NaCl, 20 mM Tris–HCl, pH 7.5]) containing 1% BSA (Sigma-Aldrich S.r.l., Milano, Italy) plus 1% blotting grade blocker non-fat milk (Bio-Rad-Laboratories, Hercules, CA) and subsequently membranes were washed twice with TBS containing 0.05% Tween-20 (TTBS) before incubation for 1 h at room temperature with anti-phosphokinase antibodies, as described earlier, diluted in TBS containing 1% BSA. After being washed six times with TTBS for 3 min, poly(vinylidene difluoride) (PVDF) membranes were incubated at room temperature for 2 h with anti-mouse or anti-rabbit IgG horseradish peroxidase-linked (HRP) secondary antibodies diluted 1:3,000.
Cytokine release
All assays were carried out using 3 × 106/mL U937 cells depleted of cholesterol (or not) and repleted (or not) and then stimulated with different concentrations of stimuli (5 µg/mL of porins or 1 µg/mL of LPS) for 24 h at 37°C in 5% CO2; the time points and concentrations have been established in preliminary experiments as that giving maximum release. After incubation the samples were centrifuged at 1,800 rpm at 4°C for 10 min and the supernatants were collected and stored at −70°C. TNF-α and interleukin-6 (IL-6) releases were measured by enzyme-linked immunoassay (ELISA) using pair-matched monoclonal antibodies, according to the manufacturer’s recommendations (Roche Diagnostic SpA, Milan, Italy). In selected experiments, after alteration of the cellular cholesterol (or not), before stimulation, U937 cells were pretreated, for 60 min, with AG490 (10 μM) (Calbiochem-Novabiochem Corp., San Diego, CA), a specific JAK/STAT pathway inhibitor; H7 (25 μM), a serine/threonine kinase inhibitor; PP2 (5 μM), a potent and selective inhibitor of the Src family of protein tyrosine kinases; calphostin (1 μM), highly specific inhibitor of PKC. The optimal concentrations of inhibitors were calculated according to 50% inhibitory concentration (IC50) values and specificities reported in literature (Jarvis and others 1994; Ku and others 1997; Kirken and others 1999; Salazar and Rozengurt 2001).
Lactate dehydrogenase (LDH) assay
Lactate dehydrogenase (LDH) assays were carried out according to manufacturer’s instructions using a cytotoxicity detection kit (Roche Diagnostic SpA, Milan, Italy). LDH is a stable cytoplasmic enzyme present in all cells and is rapidly released into cell culture supernatant when the plasma membrane is damaged. LDH activity was determined by a coupled enzymatic reaction whereby the tetrazolium salt was reduced to formazan. An increase in the number of dead or damaged cells resulted in an increase in LDH activity in the culture supernatant. The LDH assays were performed at the same time points as the cytokine assays.
Reproducibility
Gels were scanned for densitometry analysis by Sigma Gel software (Sigma-Aldrich S.r.l., Milano, Italy). The results shown are from a single experiment typical of at least three giving similar results. The results were expressed as mean values ± standard errors of three independent observations. Statistically significant values were compared using Student’s t-test and P ≤ 0.01 were considered statistically significant.
Results
Cholesterol depletion inhibits the phosphorylation of Src, PKC, JNK, and p38 by porins in U937 cells
DRMs are enriched in molecules that play pivotal roles in intracellular signal transduction. These molecules include MAPKs, Src family kinases, and PKC (Liu and others 1997). These signaling pathways, predominantly Src family kinases, PKC and MAP kinases, have been implicated in LPS or porin responses in macrophages (Riethmüller and others 2006).
To evaluate whether the reduction of the cellular cholesterol content may affect the kinase phosphorylation induced by S. enterica serovar Typhimurium porins, U937 cells (3 × 106 cells/mL) were incubated with 10 mM mβCD for 30 min at 37°C to deplete cellular cholesterol before stimulation with 5 μg of porins. LPS extracted from the same strain at a concentration of 1 μg/mL has been used as a positive control. In particular, mβCD is an highly specific cholesterol acceptor, enhancing the movement of cholesterol from the cell membrane into its hydrophobic pocket, which results in mβCD remaining outside the cell and thereby disrupting DRM in such a way as to the leave the cell intact (Ilangumaran and Hoessli 1998).
The optimal concentrations and time points of stimulations were selected by preliminary experiments (data not shown) and according to our previous experience (Galdiero and others 2002; Galdiero and others 2003). Both the concentration and time of U937 cells treatments with mβCD were titrated to minimize exposure.
Figure 1A shows a representative anti-pSrc Western blot of lysates from cell control, mβCD-treated and repleted in cholesterol cells and cells that have been afterward stimulated with an optimal dose of porins (5 μg/mL) or LPS (1 μg/mL) for 10 min (optimal time point). The mβCD cell treatment decreased of 66 ± 0.2% and 60 ± 0.7% porins- and LPS-mediated Src phosphorylation, respectively; this effect was reversed by cholesterol repletion. Figure 1A also depicts a representative anti-pJNK and anti-phospho-p38 Western blot of lysates from control and mβCD-treated cells that have been stimulated with an optimal dose of porins or LPS. The treatment of monocytes with porins or LPS induced an evident increase in JNK phosphorylation, while the pretreatment with mβCD reduced the JNK activation in porins or LPS-stimulated U937 cells (Fig. 1A). In these conditions, JNK phosphorylation decreased of 75 ± 0.7% and 73 ± 1.7% following mβCD treatment for porins and LPS stimulation, respectively.
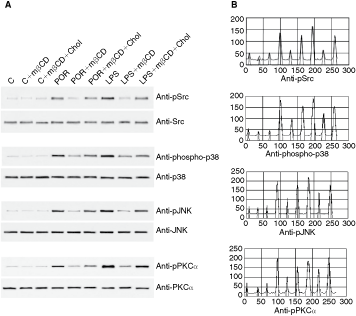
Effect of cholesterol depletion on kinase phosphorylation. U937 cells (3 × 106/mL), incubated with 10 mM mβCD for 30 min a 37°C to deplete cellular cholesterol, were stimulated at 37°C with Salmonella enterica serovar Typhimurium porins (5 μg/mL) or lipopolysaccharide (LPS) (1 μg/mL) for 10 min. The immunoprecipitated proteins for Src, p38, JNK, or PKC-α were separated by electrophoresis, transferred to PVDF membrane, and probed with anti-phospho-specific form antibodies of each enzyme. Shifts in band mobility on SDS-PAGE due to phosphorylation were obtained with anti-Src, anti-p38, anti-JNK, and anti-PKC-α (
Moreover, we found that p38 phosphorylation, induced by porins or LPS stimulation, was reduced by mβCD cholesterol depletion (60 ± 2.1% and 46 ± 1.4%, respectively), while the subsequent cholesterol repletion restored the intensity of p38 and JNK immunoprecipitation bands (Fig. 1A).
Porins (5 μg/mL) or LPS (1 μg/mL) treatment induced PKC-α phosphorylation in U937 cells. PKC-α was immunoprecipitated from porins or LPS-activated U937 and Western blot experiments, using purified anti-phospho-PKC-α antibodies, were performed. The pretreatment with mβCD decreased PKC-α activation in porins or LPS-stimulated U937 cells (Fig. 1A). PKC-α phosphorylation was inhibited by mβCD of 61.2 ± 1.3% and 59.2 ± 1.2% for porins and LPS stimulation, respectively. The following cholesterol repletion reported the intensity of PKC-α immunoprecipitation bands approximately to phosphorylation levels induced by porins or LPS. These results suggest a role for plasma membrane cholesterol in the control of porins or LPS cell activation by the PKC-α signaling pathway.
To further characterize the cells under conditions of mβCD treatment, we determined that untreated cells contain 9,887 ± 61.5 ng of cellular cholesterol/106 cells and after treatment with mβCD, the amount of cholesterol was determined to be 3,953 ± 38.2 ng (Table 1). These results are consistent with previously described levels of cellular cholesterol depletion for a variety of cell types under similar conditions (Christian and others 1997; Gimpl and others 1997). All experiments in which cellular cholesterol levels were altered were done in the absence of FCS in order to ensure that cholesterol did not re-incorporate back into the membrane, unless otherwise stated.
C
In view of the impact of cholesterol on membrane integrity, we also assessed the possibility that mβCD treatment might cause membrane damage and, in turn, lead to leakage of cytosolic molecules such as LDH. In fact it did not induce any significant release of LDH in the cell supernatants after cholesterol deprivation by mβCD (data not shown).
TNF-α and IL-6 release in cholesterol-depleted U937 cells stimulated with porins
Since TNF-α and IL-6 production involve Src, PKC, and MAP kinases signaling pathways activation, we investigated how cholesterol depletion affects cytokines release in U937 cells stimulated with porins. Porins and LPS induce the release of IL-6 and TNF-α in several cell types (Galdiero and others 2004; Munford 2005).
To determine the ability of porins to induce TNF-α and IL-6 production by cholesterol-depleted U937, cells were incubated with optimal concentration of porins for 24 h, and then the culture supernatant were assayed for the levels of cytokines (Figs. 2 and 3).

Effect of cholesterol depletion/repletion on tumor necrosis factor-α (TNF-α;) P ≤ 0.01 versus untreated cells; ( ) P ≤ 0.01 versus mβCD-treated cells; (*) P ≤ 0.01 versus mβCD+cholesterol-treated cells (Student's t-test).
) P ≤ 0.01 versus mβCD-treated cells; (*) P ≤ 0.01 versus mβCD+cholesterol-treated cells (Student's t-test).

Analysis of the effects of select inhibitors on tumor necrosis factor-α (TNF-α;) P ≤ 0.01 versus untreated cells; (○) P ≤ 0.01 versus inhibitor pretreated cells; ( ) P ≤ 0.01 versus mβCD-treated cells; (*) P ≤ 0.01 versus mβCD+cholesterol-treated cells (Student’s t-test).
) P ≤ 0.01 versus mβCD-treated cells; (*) P ≤ 0.01 versus mβCD+cholesterol-treated cells (Student’s t-test).
As shown in Figure 2A, TNF-α release in U937 cells, treated with mβCD and then stimulated with porins, was considerably decreased (67 ± 1.4%); while the TNF-α release was slightly reduced when the cells were preincubated with mβCD and stimulated with LPS (16 ± 1.06%). The cholesterol repletion reported TNF-α release to porins-induced activation level.
In contrast, IL-6 production as a consequence of porin or LPS stimulations was unaltered in cholesterol-depleted cells (Fig. 2B).
To demonstrate the requirement of Src, PKC, and MAP kinases signaling pathways in porins or LPS-induced TNF-α production, we next assessed the effect of specific pharmacological inhibitors (Fig. 3). Specifically, we looked at the effects of the PP2, H7, calphostin C, and AG490 inhibitors. Initially, we determined the effect of the dose of each inhibitor on the cell viability. The inhibitor concentration used, as well as the duration of the treatment, did not indicate significant cytotoxicity in these cultures as determined by LDH assay (data not shown).
Cells depleted of cholesterol, pretreated with PP2 (5 μM for 60 min), a potent and selective inhibitor of the Src family of protein tyrosine kinases, and stimulated with porins (5 μg/) or LPS (1 μg/mL) decreased considerably TNF-α release. The cholesterol repletion was able to restore the levels of TNF-α release to values typical of cells stimulated with porins or LPS. The H7, a serine/threonine kinase inhibitor (25 µM for 60 min), completely inhibited the TNF-α release from porins or LPS-stimulated U937 cells and cholesterol depletion was unable to further modify this inhibition. The pretreatment with calphostin, a selective PKC inhibitor (1 μM for 60 min), also inhibited TNF-α release induced by porins or LPS. Also with this inhibitor, the modification of plasma membrane cholesterol did not further modify the TNF-α production. We finally have investigated the effect of AG490 on the induction of porins or LPS-mediated TNF-α secretion. Pretreatment of U937 cells with AG490 (10 μM for 60 min) decreased slightly porins TNF-α release (about 25 ± 2.1%), while LPS TNF-α release was decreased by AG490 (about 43 ± 0.3%). Porins TNF-α release was further decreased by cholesterol depletion in presence of AG490, while LPS TNF-α release was not modified by mβCD treatment in presence of AG490. The cholesterol repletion restored the TNF-α release to values typical of cells stimulated with porins or LPS (Fig. 3A).
The release of IL-6 was differently modulated by the inhibitors under analysis (Fig. 3B). The effect of PP2 pretreatment did not significantly influence IL-6 release in porins or LPS-stimulated U937 cells, and modification of plasma membrane cholesterol did not induce a significant decrease in the release of IL-6. The porin-mediated IL-6 secretion was considerably decreased by H7 (25 μM for 60 min) or calphostin (1 μM for 60 min) preincubation in porins or LPS-stimulated U937 cells; the depletion or repletion of cholesterol did not further modify the release. In contrast, AG490 pretreatment (10 μM for 60 min) showed a modest inhibiting effect on IL-6 secretion induced by porins, while decreased the LPS-mediated IL-6 release about 39 ± 0.3%. The cholesterol depletion obtained by mβCD did not significantly modify the reduction of IL-6 due to porins or LPS in presence of AG490 (Fig. 3B).
Discussion
Over the last few years, it has become increasingly apparent that many pathogens (bacteria, parasites, and viruses) and toxins preferentially utilize lipid domains when interacting with their target cell (Hessey and others 1990; Gatfield and Pieters 2000; Lencer 2001; Pieters 2001; Patel and others 2002; Popik and others 2002; Jutras and others 2003).
The cholesterol depletion agent mβCD has been used in several transduction-dependent systems to study the effects of cholesterol remotion, including epidermal growth factor receptor (Hur and others 2004) and T-cell receptor signaling (Nagafuku and others 2003). It was previously reported that mβCD treatment transiently induces tyrosine phosphorylation of multiple proteins including Ras-ERK pathway, but did not stimulate p38/JNK (Kabouridis and others 2000).
The mechanisms of porins and LPS stimulation of signaling pathways have been recently subjected to extensive studies. The predominant Src family kinases expressed in macrophages, Hck, Fgr, and Lyn, have been implicated in a number of studies as being the initial transducers for tyrosine phosphorylation events after LPS stimulation (Beaty and others 1994; Roskoski 2004; Lee and others 2005), but little is known concerning the relationship between porins and Src. Src family are frequently associated with the Gi proteins. Src family protein tyrosine kinases are activated in response to stimulation of a variety of G-protein-coupled receptors and are necessary for linking Gi- and Gq-coupled receptors with MAPK activation (Dikic and others 1996). Our results show that Src, connected to lipidic domains, play a fundamental role in TNF-α but not in IL-6 release by porins.
TNF-α and IL-6 are two important players in the pathogen inflammatory response. TNF-α is involved in several events including bacterial killing, leukocyte migration, fever, and acute phase responses (Zeytun and others 2007). IL-6 is a multifunctional cytokine that plays an important role in immune and inflammatory responses. TNF-α and IL-6 are produced mainly by cells of the monocyte lineage. The important role of Raf-MAPK/NF-κB pathways in the production of these cytokines in response to porins is well described (Galdiero and others 2002); the role of other signaling pathways at the membrane surface is less clear. In particular, little is known of the role of Src family kinases in porins induction of cellular phosphorylation and TNF-α and IL-6 release.
Our results shows that depleting membrane cholesterol levels disrupts porins signaling suggesting that porins probably activate some kinases associated with DRM components. LPS instead can bind to DRMs but alternatively binds its receptors that phosphorylize other signaling pathways. A number of papers have previously described that manipulation of cholesterol in cells alters macrophage activation by LPS by mechanisms that interfere with receptor-lipid rafts recognition or by alteration in the activities of protein phosphatases associated with lipid rafts (Wang and others 2003; Koseki and others 2007; Dhungana and others 2009).
Our results demonstrate that cholesterol plays a critical role, also, in the initial step of porins interaction with cell membrane through the stimulation of p38/JNK pathway by Src. The reduction of cellular cholesterol by mβCD causes both the decrease of activation of signaling pathways and the decrease of TNF-α production. Restoration of the cholesterol content of the depleted cells restores partially the phosphorylation of the kinases under study and TNF-α production induced by porins or LPS. Cells treated with mβCD and then stimulated with porins or LPS showed a significant inhibition of phosphorylation of Src, which is completely reversed after the repletion of cholesterol.
These findings suggested that Src-p38/JNK kinases are involved in TNF-α production following porins stimulation. In fact, inhibition of Src kinases with PP2 and inhibition of p38/JNK with H7 in cells depleted of cholesterol caused a significant reduction of TNF-α released after porins treatments. Since Src kinase are localized in DRMs, that can function as signal platforms, the observation that Src kinases were necessary for the induction of TNF-α by S. enterica serovar Typhimurium porins suggests that DRMs may be involved in porins-mediated cell signaling activation.
We have also verified, using others specific pharmacological inhibitors, that the others NT-PTK (JAK and PKC), which are not correlated with lipidic domains, were involved in signal transduction by porins. The JAK inhibitor AG490 does not influence the release of TNF-α by porins, while calphostin, a PKC inhibitor, reduces drastically the release of TNF-α by porins.
The release of IL-6 was not modified in cells depleted of cholesterol stimulated either with porins or LPS. The Src inhibitor, PP2, did not modify the release of IL-6, while JAK inhibitor AG490 did not influence the release of IL-6 in cells stimulated by porins, but caused a significant reduction in cells stimulated with LPS demonstrating that porin-mediated IL-6 release can take place through pathways different from the JAK/STAT pathway that is at least partially involved by LPS stimulation. H7 and calphostin C, inhibitors of p38/JNK and PKC, respectively, reduced IL-6 production in LPS- or porin-stimulated cells in absence of mβCD treatment. When the same cells were depleted of cholesterol, the IL-6 production was no further modified after LPS or porin stimulation. PKC seems to be the major pathway involved in production of IL-6. Inhibition of PKC by calphostin C and inhibition of JNK/p38 by H7 induce in both our experimental models (cells depleted in cholesterol or not) a strong inhibition of IL-6 production. We can observe a residual production of IL-6 due to the involvement of other phosphorylation pathway, such as JAK pathway.
It is well established that different signaling pathways are involved in the release of TNF-α and IL-6 cells following porins stimulation. To our knowledge, the data presented here represent a first demonstration that depletion of cellular cholesterol levels modulate cytokine release induced by porins, raising the possibility that sepsis inflammatory processes will be modulated by Src DRM-associated. Our experimental evidences indicate that Src membrane associated with DRMs (Hoessli and others 2000) may represent clinically relevant potential targets for inflammatory and immune regulation. Recently, Zhu and others have demonstrated that cellular free cholesterol in macrophage enhances proinflammatory response to LPS in vivo (Zhu and others 2008).
Much more detailed elucidation of DRM structure and function will improve understanding of biological membrane composition–structure–function relationships and also may serve as a new avenue for the development of novel treatments for major infectious diseases.