
Abstract
The clinical prognosis of children with high-stage neuroblastoma is still poor. Therapeutic approaches include surgery and cellular differentiation by retinoic acid, but also experimental interleukin-based immune modulation. However, the molecular mechanisms of all-trans retinoic acid (ATRA)-induced differentiation of neuroblastoma cells are incompletely understood. Herein, we examined the effect of ATRA on the activity of the interleukin-18 (IL-18) system in human SH-SY5Y neuroblastoma cells. It is shown that SH-SY5Y cells express IL-18 receptor (IL-18R) and the secreted antagonist IL-18-binding protein (IL-18BP), but no IL-18. SH-SY5Y cells are highly sensitive to ATRA treatment and react by cellular differentiation from a neuroblastic toward a more neuronal phenotype. This was associated with induction of IL-18 and reduction of IL-18BP expression, while IL-18R expression remained stable. Thereby, we identified the IL-18 system as a novel target of ATRA in neuroblastoma cells that might contribute to the therapeutic properties of retinoids in treatment of neuroblastoma.
Introduction
N
Retinoids play an essential role during normal development of the nervous system. They stimulate primary neurogenesis and promote neuronal specification as well as neurite elongation. Nevertheless, high-dose retinoids are neurotoxic and cause neurodevelopmental disorders (McCaffery and others 2003). In vitro, some neuroblastoma cell lines are very sensitive to retinoic acid treatment. Human SH-SY5Y neuroblastoma cells, for example, respond to all-trans retinoic acid (ATRA) by growth inhibition and cellular differentiation, but also by inducing early proapoptotic genes (Reynolds and others 2003). Binding of ATRA to retinoic acid receptor (RAR) and/or retinoid X receptor (RXR) induces formation of transcriptionally active complexes. The known downstream targets of ATRA include neurotrophin receptor genes (Clagett-Dame and others 2006), but the complex molecular mechanisms underlying retinoic acid action in neuroblastoma cells are still incompletely understood (Reynolds and others 2003).
Interleukin-mediated immune modulation is a novel experimental approach for treatment of neuroblastoma. Interleukin-18 (interferon-γ-inducing factor; IL-1γ) has been shown to reduce the tumor size in animal models of neuroblastoma, likely by activating the immune response against the tumor (Redlinger and others 2003). The interleukin-18 (IL-18) system consists of interleukin-18, its receptor (IL-18R), and the secreted antagonist interleukin-18-binding protein (IL-18BP). We hypothesized that the IL-18 system is available in neuroblastoma cells as they share many properties with neural progenitor cells, which are known to express the IL-18 system (Felderhoff-Mueser and others 2005). We furthermore hypothesized that the IL-18 system in neuroblastoma cells is regulated by ATRA, since retinoids have been shown to induce IL-18 expression in embryonic stem cells (Sarkar and Sharma 2002).
First, we analyzed the expression of IL-18, IL-18R, and IL-18BP mRNA in undifferentiated SH-SY5Y cells. Preparation of mRNA and RT-PCR analysis were carried out as previously described (Reinhold and others 2007). The primer sequences and annealing temperatures were published elsewhere (Yoshino and others 2001), only the annealing temperature for IL-18R was changed to 58°C. We found almost no expression of IL-18 mRNA (35 cycles), while IL-18R and IL-18BP mRNA (31 cycles) were clearly expressed in SH-SY5Y cells (Fig. 1A).

Modulation of the interleukin-18 (IL-18) system by all-trans retinoic acid (ATRA) in human SH-SY5Y neuroblastoma cells. (
To test whether the IL-18 system is regulated by ATRA, SH-SY5Y cells were exposed to 10 µM ATRA treatment for up to 8 days. The cellular differentiation was confirmed by the immunocytochemical detection of microtubulus-associated protein-1 (MAP-1), which is known to be expressed in neurites and in the cytoplasm of neuroblastoma cells (Wiche and others 1983) (Fig. 2). Undifferentiated SH-SY5Y cells appeared round-shaped and displayed short neurites (Fig. 2A), while differentiated cells clearly showed staining of the prolonged neurites (Fig. 2B).
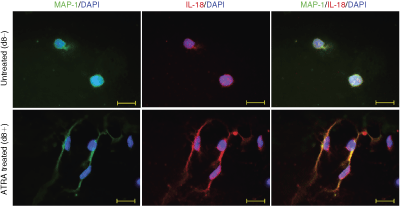
Effects of all-trans retinoic acid (ATRA) on the cellular differentiation and on interleukin-18 (IL-18) protein expression in human SH-SY5Y neuroblastoma cells. For immunocytochemical staining, cells were grown without (controls) or with 10 µM ATRA on glass slides with removable plastic wells. After 8 days, cells were fixed with 4% PFA for 10 min at room temperature. Permeabilization and nonspecific binding block were carried out by incubating the slides for 1 h at room temperature in blocking solution (1% bovine serum albumin, 2% rabbit serum, and 0.1% Triton X-100 in PBS). For MAP-1 detection, cells were incubated overnight (4°C) with a monoclonal anti-MAP-1 antibody (No. 4278, 1:500; Sigma-Aldrich Chemie). After washing three times with PBS, the cells were incubated overnight with polyclonal anti-human pro-IL-18/IL-1F4 antibody (No. AF646, 1:250; R&D Systems) at 4°C and rinsed again with PBS. Negative controls were prepared by following the same staining procedure in the absence of primary antibodies. Fluorescent goat anti-mouse (Alexa Fluor 488, MoBiTec; green) or donkey anti-goat antibodies (Cy3-conjugated anti-goat IgG (No. 705-226-147), Jackson ImmunoResearch, red, each at a dilution of 1:1,000) were used for detection of the respective protein and incubated for 1 h at room temperature. Counterstaining was performed with aqueous 4,6-diamidino-2-phenylindole (DAPI) containing fluorescence protecting mounting media (Vectashield, Vector Laboratories). Microscopic analysis was performed under an epifluorescence microscope (AxioPlan 2e, Imaging System; Carl Zeiss). Photographs were taken with a connected digital camera (AxioCAM MRc; Carl Zeiss) using AxioVision 4.2 software (Carl Zeiss). Bar = 20 µm.
Finally, we repetitively analyzed the mRNA expression levels of IL-18, IL-18BP, and IL-18R in ATRA-treated SH-SY5Y cells at days 4 and 8 by means of RT-PCR. At a dose of 10 µM ATRA, which has been found to be optimal for in vitro differentiation (Påhlman and others 1984), we found increased expression of IL-18 mRNA and decreased expression of IL-18BP, while IL-18R was constitutively expressed (Fig. 1B). The effects of ATRA on the IL-18 system were also obtained in SH-SY5Y cells treated with 1 µM ATRA, a 10-fold lower dose than usually used for in vitro differentiation of SH-SY5Y cells (Påhlman and others 1984; Klinz and others 1987; Cuende and others 2008). If cells were treated with 0.3 µM, which corresponds to an ATRA serum concentration of 100 ng/mL that is normally reached in patients 1 to 5 h after ATRA application (Bonhomme-Faivre and others 1996), we still found up-regulation of IL-18 mRNA expression, but no significant effect on IL-18BP mRNA expression (Fig. 1B). We confirmed the effect of 10 µM ATRA on the protein levels by Western blot analysis (IL-18BP and IL-18R, Fig. 1C) and ELISA (IL-18, Fig. 1D). Notably, the effect of ATRA on IL-18 was not observed in Kelly cells, another neuroblastoma cell line that amplifies MYCN. Since Kelly cells treated with ATRA undergo earlier apoptosis and showed in comparison to SH-SY5Y cells differential expression of other genes in response to ATRA (such as erythropoietin; Hoene V. and Dame C, unpublished). ATRA may modify the IL-18-mediated NK cell-based antitumor response only in a certain type of neuroblastoma cells.
In conclusion, we identified the IL-18 system in certain human neuroblastoma cells as a novel target of ATRA-induced differentiation. ATRA induces the expression of IL-18, whereas it reduces the expression of IL-18BP. The induction of IL-18 is concordant with previous data showing that ATRA increases IL-18 mRNA expression in murine embryonic stem (ES-D3) cells after 11 to 21 days of treatment (Sarkar and Sharma 2002). The combined data provide evidence that IL-18 secretion is directly mediated by ATRA in a dose-dependent manner. In our experiments, the expression of IL-18R was not modified by ATRA. Therefore, one might conclude that IL-18R is constitutively expressed. Nevertheless, Jeon and others have recently shown that IL-18R is up-regulated in murine astrocytes of the hippocampus in response to kainic acid-induced excitotoxic brain damage (Jeon and others 2008), suggesting that neural IL-18R expression can be regulated by yet unknown factors.
IL-18 stimulates a NK cell-based antitumor response in animal models of neuroblastoma (Redlinger and others 2003). It is therefore likely that endogenous IL-18 from ATRA-exposed neuroblastoma cells enhances this immune response. Furthermore, increased IL-18 activity in neuroblastoma cells could be directly involved in neuroblastoma cell apoptosis in a manner similar to its autocrine and/or paracrine action on neural progenitor cells (Felderhoff-Mueser and others 2005). In fact, recent work showed that recombinant IL-18 induces the protein expression of p35, cyclin-dependent kinase 5 (Cdk5), glycogen synthetase kinase GSK-3β, and Ser15-phosphorylated p53 in differentiated SH-SY5Y neuroblastoma cells within 24 h, and Tau phosphorylation and expression of cyclin G1 within 72 h (Ojala and others 2008). Thereby, IL-18 may mediate beneficial effects of all-trans retinoic acid on tumor cell differentiation. For example, GSK-3β is involved in signaling pathways that are implicated in neuronal polarity (Beffert and others 2002), and Cdk5, which is activated by p35, contributes to neuronal differentiation by affecting cytoskeleton structure and organization (Ohshima and others 1996).
Nevertheless, further in vivo experiments with animal models of neuroblastoma (Redlinger and others 2003; Lode and others 2008) are highly warranted to elucidate the molecular mechanisms of IL-18-based immune modulation as well as ATRA-induced cellular differentiation of neuroblastoma in order to improve the therapeutical strategies against this disease.
Footnotes
Acknowledgments
The expert technical assistance of Evelyn Strauss is acknowledged. The work of C.D. is supported by the Fritz Thyssen Stiftung (Essen, Germany; Az. 10.05.2.162).
Author Disclosure Statement
No competing financial interests exist.