
Abstract
It has been suggested that hepatitis C virus (HCV) infects not only hepatocytes but also immune cells, including B cells. HCV infection of B cells is the likely cause of B-cell dysregulation disorders such as mixed cryoglobulinemia, rheumatoid factor production, and B-cell lymphoproliferative disorders that may evolve into non-Hodgkin’s lymphoma. To clarify the effects of chronic HCV infection on B-cell dynamics, peripheral B cells from chronic hepatitis C patients (CHC) were characterized. We found that the frequency of CD27+ B cells, that is memory phenotype, was significantly reduced in the peripheral blood of CHC. At the same time, the amount of IFN-γ-inducible protein-10 (IP-10), a CXCR3 ligand, was markedly elevated in the plasma of CHC. Furthermore, the CD27+ B-cell population was found to highly express CXCR3 in CHC, thus suggesting that the CD27+ B-cell population was recruited from peripheral blood to the inflammatory site of the liver of CHC, where IP-10 is produced. Immunohistochemical analyses of intrahepatic lymphocytes indicated that CXCR3+ B cells were infiltrated in the liver of CHC. Our results thus offer new insight into the role of memory B cells in the HCV pathogenesis.
Introduction
N
HCV is a single-stranded, positive polarity RNA virus of the Flaviviridae family. CD81, a widely expressed tetraspanin molecule, has been shown to interact with the E2 region of HCV envelope protein (Pileri and others 1998), and is regarded as one of the key molecules for HCV infection. Although the liver is the major target organ for HCV, it also infects peripheral blood mononuclear cells (PBMCs), preferentially B cells that express CD81 (Dammacco and others 2000; Pal and others 2006). This B-tropism of HCV may be responsible, at least in part, for the extrahepatic manifestations of HCV infection, including mixed cryoglobulinemia (MC), Sjoegren-like syndrome, production of rheumatoid factor (RF), and B-lymphoproliferative disorder, which may evolve into non-Hodgkin’s lymphoma (NHL; Galossi and others 2007; Zignego and others 2007). However, the mechanisms that underlie these immunological dysfunctions are poorly understood.
It has been demonstrated that CD81 molecule on human B cells forms a co-stimulatory complex with CD19 and CD21, and that cross-linking this complex lowers the threshold required for antigen-induced B-cell proliferation (Levy and others 1998). In the same way, binding of HCV E2 protein to the CD81/CD19/CD21 complex may affect the immune system by facilitating B-cell activation, and this may result in lymphoproliferative disorders.
The primary site of HCV infection and inflammatory response is the liver, in which various kinds of immunopathogenic events associated with the HCV infection take place. The infiltration of activated B cells recruited from draining lymph nodes increases during the course of disease progression, suggesting that infiltrating B cells play a role in the pathogenesis of chronic hepatitis (Vallat and others 2004), although the phenotype and characteristics of such B cells have not been well documented to date. It is plausible that infiltrating B cells in the liver are the malignant precursor population, as they are probably stimulated by inflammatory cytokines, and because HCV E2 protein and HCV have been known to induce DNA damage and mutation of cellular genes (Machida and others 2004, 2005, 2006).
In this study, we sought to analyze the effects of HCV infection on the fate of peripheral CD27+ memory B cells that are thought to be vulnerable to antigenic stimulation. It was found that the frequencies of CD27+ B cells that strongly express CXCR3 molecules, a receptor for IP-10, in the peripheral blood were significantly reduced compared with healthy subjects. Furthermore, the plasma level of IP-10 in CHC was markedly elevated. In accordance with these phenomena, CXCR3+ B cells were detected in the CHC liver. The possible mechanisms controlling this memory B-cell recruitment in CHC are considered with regard to lymphocyte trafficking mediated by chemokines produced in the liver.
Materials and Methods
Patients and samples
Nineteen CHC were enrolled in this study. Demographic and clinical data were as follows: gender (M:F = 10:9); mean age (range) = 59.6 years (38–79 years); HCV genotype (No.) = 1b (18) and 2a (1); mean HCV RNA (KIU/mL) (SD) = 2,077 (1,016); mean alanine aminotransferase (ALT) (U/L) (SD) = 64.4 (23.5); and mean aspartate aminotransferase (AST) (U/L) (SD) = 61.3 (19.3). The details of this study were approved by the Review Board at the National Institute of Infectious Disease. All donors gave written informed consent before phlebotomy. Fourteen normal control subjects (CS) included anonymous volunteer blood donors visiting the Japanese Red Cross Blood Center.
Liver biopsy specimens were collected and stored until use at −80°C from 8 patients who gave written informed consent, including 5 patients with persistent HCV infection and 3 patients without HCV infection. Genotypes and plasma RNA titers of these patients with HCV infection are similar to those of the above 19 CHC.
Clinical tests
HCV genotype was determined by PCR of the core region with genotype-specific primers (Ohno and others 1997). HCV RNA was quantified by the Roche Amplicor assay (Roche Diagnostics, Branchberg, NJ), and results were standardized to international units (IU). Determination of the serum levels of ALT and AST was performed using standard methods.
Antibodies
The following directly conjugated Abs were used for flow cytometry: Allophycocyanin-anti-CD19 (MHCD1905; Invitrogen, Carlsbad, CA); FITC-anti-CD27 (555440; BD Biosciences, Franklin Lakes, NJ); and PE-anti-CXCR3 (FAB160P; R&D Systems, Minneapolis, MN). The following primary antibodies were used for immunohistochemical analyses: anti-CD19 (MCA1940; AbD serotec, Germany); anti-CXCR3 (SC-1395; Santa Cruz Biotechnology, Santa Cruz, CA); and anti-IP-10 (AF-266-NA; R&D Systems). The following Alexa-labeled secondary antibodies were purchased from Molecular Probes Inc., Invitrogen, Carlsbad, CA: Alexa635-Goat anti-mouse IgG (A31574); Alexa647-Rabbit anti-goat IgG (A21446); and Alexa488-Goat anti-mouse IgG (A11001).
Flow cytometry
Cells were washed twice with cold PBS containing 0.2% BSA, followed by incubation with an appropriate combination of directly conjugated Abs for 30 min on ice. Stained cells were analyzed by FACSCalibur (Becton Dickinson, San Jose, CA). Data were collected using CellQuest software (Becton Dickinson) and were analyzed using FlowJo software (Tree Star Inc., Ashland, OR).
Double fluorescence immunohistochemical analysis
Frozen sections (3 mm) of liver biopsy samples were fixed with ethanol for 10 min and with acetone for 1 min at −20°C, and then washed once with PBS. Primary antibodies were added, followed by incubation for 30 min at room temperature and washing with PBS (3×). Secondary antibodies were then added, followed by incubation for 30 min at room temperature and washing with PBS (3×). Additional primary antibodies were added, followed by incubation for 30 min at room temperature and washing with PBS (3×). Additional secondary antibodies were added, followed by incubation for 30 min at room temperature and washing with PBS (3×). Concentrations of primary antibodies used for double fluorescence staining were as follows: anti-CD19 (1:400); anti-CXCR3 (1:100); and anti-IP-10 (15 µL/mL). Concentrations of fluorescence-labeled secondary antibodies were as follows: Alexa635-Goat anti-mouse IgG (1:2,000); Alexa647-Rabbit anti-goat IgG (1:2,000); and Alexa488-Goat anti-mouse IgG (1:2,000). During the incubation period, light transmission was carefully avoided. Stained samples were then washed with distilled water for 1 min and mounted using Aqua poly/Mount (#18606, Polysciences, Inc., Warrington, PA). Stained samples were analyzed under a fluorescence microscope (Nikon C-SHG1, Nikon ECLIPSE E600; Nikon Co., Tokyo, Japan). BZ8000 fluorescence microscope (Keyence Co., Osaka, Japan) was used for deconvoluted fluorescence imaging in co-localization analyses.
Quantification of immunofluorescent pictures
Quantification of green (CXCR3) or red (CD19 and IP-10) fluorescing dots in the pictures was performed with the image analysis software, MetaMorph (Molecular Devices Co., Japan, Tokyo, Japan).
Plasma cytokine/chemokine/growth factor assay
Plasma samples were analyzed for IL-1β, IL-1RA, IL-2, IL-2R, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12p40/p70, IL-13, IL-15, IL-17, TNF-α, IFN-α, IFN-γ, GM-CSF, MIP-1α, MIP-1β, IP-10, MIG, Eotaxin, RANTES, MCP-1, VEGF, G-CSF, EGF, FGF-basic and HGF using a Human Cytokine Thirty-plex antibody bead kit (BioSource Int. Inc., Camarillo, CA) according to the manufacturer’s instructions. Assay results were obtained using a Luminex 100™ reader (Luminex Co., Austin, TX).
Statistics
Unpaired (two-tailed) Student’s t-test was applied at 95% confidence level (P < 0.05) using Prism ver. 4 (GraphPad Software, Inc., San Diego, CA) in all cases.
Results and Discussion
Decreased frequency of peripheral CD27+ B cell in CHC
Two major B-cell subsets can be identified in human peripheral blood based on the expression of CD27, a member of the TNF receptor family. Functional differences between the two subsets have been extensively investigated and it is now generally accepted that CD27 is a memory B-cell marker (Agematsu and others 2000). As memory B cells are quickly activated upon antigenic stimulation and can rapidly produce high levels of immunoglobulins, we initially speculated that the frequency of CD27+ memory B cells is increased in the peripheral blood of patients with persistent HCV infection as a consequence of chronic antigenic stimulation by HCV antigens, such as the aforementioned E2 protein. We thus analyzed the frequencies of CD27+ B cells in peripheral blood of control subjects (CS; n = 14) and chronic hepatitis C patients (CHC; n = 19) by flow cytometry.
Representative profiles (CD19 vs. CD27) of both CS and CHC are shown in Figure. 1A and 1B, respectively. It should be noted that the percentages of peripheral CD19+ B cells in CHC were comparable to those of CS (P = 0.7233) (data not shown). In contrast to our expectations, the frequency of CD27+CD19+ B cells in CHC was lower than that in CS. As summarized in Figure 1C, it was concluded that the percentages of CD27+ cells in CD19+ B cells are significantly reduced (P < 0.0001) in CHC compared with CS.

Flow cytometry analysis of peripheral blood mononuclear cells (PBMCs) from control subjects (CS) and chronic hepatitis C patients (CHC). Lymphocytes were identified by forward and orthogonal light scatter characteristics. (
Rosa et al. previously reported that HCV patients display a higher percentage of peripheral memory (CD27+) B cells when compared with healthy controls (Rosa and others 2005). In contrast, Racanelli et al. found that the frequencies of CD27+ B cells in persistently HCV-infected patients were significantly lower than those in healthy donors (Racanelli and others 2006). Ni et al. demonstrated that the frequencies of CD27+ B cells in both normal subjects and HCV patients are comparable (Ni and others 2003). Among these controversial reports, our results are in good agreement with Racanelli and others (2006). In their report, patients with higher plasma viral loads had lower percentages of CD27+ B cells, thus suggesting that high viral replication is associated with a reduction in CD27+ B cells, although the negative correlation was not statistically significant in their study (P = 0.0566; r = −0.4328). In our study, the above negative correlation was statistically significant (P = 0.0031; r = −0.7285; Fig. 1D) and thus agreeing with their suggestion.
Racanelli et al. hypothesized that under conditions of persisting HCV antigenemia, memory B cells not receiving specific B-cell receptor triggering before having T-cell help would be pushed to enhance Ig production and prone to apoptosis (Racanelli and others 2006), which may explain the reduction of CD27+ memory B cells in HCV-infected patients. In subsequent experiments, we sought to examine an alternative possibility that the CD27+ memory B cells are recruited from peripheral blood to the liver, where HCV infection triggers the production of IFN-γ and consequently induces attractants such as IFN-γ-inducible chemokines. We therefore examined plasma levels of various cytokines/chemokines/growth factors by a multiplex suspension bead array immunoassay.
Increased levels of CXCR3 ligand chemokines in plasma of CHC
The amounts of 30 different cytokines/chemokines/growth factors (see Materials and Methods) were simultaneously determined in the available plasma samples from both CS (n = 14) and CHC (n = 11; 11 out of 19 patients’ plasma were available; Fig. 2). Statistically significant (P < 0.05) differences between the 2 groups were noted for the following analytes: HGF (hepatocyte growth factor), IL-12, IL-1β, IL-2R, IL-4, IP-10 (IFN-γ-inducible protein-10), MCP-1 (monocyte chemotactic protein-1), MIG (monokine induced by IFN-γ), and MIP (macrophage inflammatory protein)-1β as shown in Figure 2. Among these, plasma levels of IP-10, a CXCR3 ligand, in CHC were markedly elevated (P < 0.0001) when compared with CS. Plasma levels of another CXCR3 ligand, MIG, were also elevated (P = 0.0048).

Plasma levels of cytokine/chemokine/growth factors. The amounts of 30 different cytokines/chemokines/growth factors (see Materials and Methods) were simultaneously determined in the available plasma samples from both control subjects (CS; n = 14) and chronic hepatitis C patients (CHC; n = 11). Nine analytes that showed statistically significant (P < 0.05) differences between CS and CHC were indicated.
These results prompted us to hypothesize that CD27+ B cells express CXCR3 and that they migrate to inflammation sites in the liver of CHC, where the levels of IFN-γ are thought to be elevated and thus IP-10 is produced. It should be noted here that the enhanced expression of IP-10 mRNA has been demonstrated in the liver of CHC (Patzwahl and others 2001). The clinical significance of elevated serum levels of IP-10 in chronic HCV carriers has been already reported, and the levels of IP-10 in patient serum are regarded as a marker for therapeutic outcome (Narumi and others 1997; Itoh and others 2001; Patzwahl and others 2001). However, the physiological and immunological grounds for IP-10 production in HCV patients remain uncertain. If our predictions were correct, peripheral CD27+ memory B cells in CHC would highly express CXCR3 and accumulate within chronically inflamed tissue, that is, HCV-infected liver. We therefore examined this possibility by flow cytometry in the following experiments.
Expression of CXCR3 on peripheral CD27+ B cells in CHC
CD19+ B cells in CS and CHC were analyzed for their CXCR3 expression levels by flow cytometry. Representative profiles are shown in Figure 3A and 3B. Both CD27− and CD27+ B cells in CS expressed CXCR3 to some extent (Fig. 3A). Expression of CXCR3 on CD27− B cells in CHC was comparable to that of CS (Fig. 3B). As expected, CXCR3 was highly expressed on CD27+ B cells in CHC (Fig. 3B), thus supporting our hypothesis that CD27+ B cells expressing CXCR3 are recruited to the liver, where levels of CXCR3 ligands, that is, IP-10 and MIG, are thought to be elevated. Figure 3C summarizes the expression levels of CXCR3 on CD27+ B cells in both CS and CHC. The results shown in Figure 3D further confirmed that CD27+ B cells of CHC express the highest amounts of CXCR3 among all the cell populations analyzed.

Expression of CXCR3 on CD19+CD27+ cells. (
These results are compatible with a study by Muehlinghaus et al. in which CXCR3 was found to be expressed on a fraction of CD27+ memory B cells but not on CD27− naïve B cells (Muehlinghaus and others 2005). Manz et al. proposed a possible mechanism for how the CXCR3-mediated recruitment of autoreactive memory B cells into inflamed tissues contributes to a break in self-tolerance (Manz and others 2006). As memory B cells have a lower threshold for antigenic activation and stably express CXCR3, which supports their accumulation within chronically inflamed sites, they may be resistant to the induction of peripheral tolerance. Furthermore, in the inflamed tissue, inflammatory cytokines and autoantigens provide survival signals that promote plasma cell differentiation and survival, which may result in chronic inflammation and progressive tissue destruction. The present results are in line with this scenario when the HCV E2 protein is regarded as an autoantigen.
MC, a chronic autoimmune disorder, is usually associated with chronic liver inflammation by HCV infection (Dore and others 2007; Galossi and others 2007; Zignego and others 2007). Excessive production of antibodies known as RF is the main cause of MC. The fact that the majority of circulating B cells of patients with HCV-associated MC are of the CD27+ phenotype (Carbonari and others 2005) further supports the contribution of memory B cells to extrahepatic autoimmune disorders.
Migration of B cells expressing CXCR3 into the CHC liver
In order to directly assess the migration of B cells expressing CXCR3 to the liver, immunohistochemical analyses were carried out by staining liver biopsy specimens. Unfortunately, as we could not obtain the biopsy specimens from the CHC analyzed in the above experiments, they were collected from other chronic hepatitis C patients. Liver biopsy specimens of alcohol-induced HCV-negative hepatitis patients instead of normal subjects were used as controls, since we could not obtain normal liver biopsy specimens.
As shown in Figure 4, the infiltration of CD19+ B cells were recognized in the liver of a HCV-positive hepatitis C patient, and the distribution of the CD19+ cells in the liver was almost identical to that of CXCR3+ cells (panels B vs. C). These CXCR3+ B-cell infiltrations were not observed in an HCV-negative alcohol-induced hepatitis patient specimen (panels E and F). Five HCV-positive and three HCV-negative independent liver biopsy specimens were analyzed, and similar staining profiles as shown in the figure were obtained. As expected, IP-10 was expressed in the liver of a CHC (panel H) but not in an HCV-negative specimen (panel J). Figure 4K–4N indicated that CXCR3+ cells are localized at the sites where IP-10 is expressed positive. Together, these results are concordant with our hypothesis that CD27+ B cells expressing CXCR3 migrate into the liver of PIP.

Infiltration of B cells expressing CXCR3 to the chronic hepatitis C patient (CHC) liver. Panels
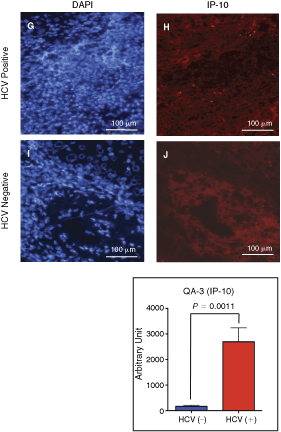

In this study, it was demonstrated that significant reductions in peripheral CD27+ B cells are seen in CHC, while plasma levels of IP-10, a CXCR3 ligand, are markedly increased in CHC. Furthermore, it was found that the CD27+ B cells highly express the CXCR3. In accordance with these phenomena, immunohistochemical analyses revealed that CXCR3+ B cells migrate in the liver of CHC where IP-10 is produced.
Finally, it is worth mentioning here that the recruitment of peripheral memory B cells into inflamed tissue may not be unique to CHC but rather is a general phenomenon. A recent report by Hansen et al. showed accumulation of memory B cells in the inflamed salivary glands of Sjoegren’s syndrome patients (Hansen and others 2002), and Malbran et al. found that peripheral CD27+ memory B cells are reduced in patients with X-linked lymphoproliferative disease (Malbran and others 2004). Further investigation into the dynamics of memory B cells in chronically HCV-infected patients will be necessary, not only to better understand the HCV pathogenesis in particular, but also to gain insight into immunological disorders such as tolerance breakdown in general.
Footnotes
Acknowledgments
We are grateful to Dr. Masayuki Saijo for multiplex suspension bead array immunoassay and for operation of the Luminex analyzer. We would also like to thank Ms. Tomoko Nagai for immunohistochemical staining, and Dr. Yoshimasa Takahashi for valuable discussion and critical review of the manuscript. This study was supported by Grants-in-Aid from the Ministry of Health, Labor, and Welfare.
Author Disclosure Statement
The authors have no conflicting financial interests.