
Abstract
The present study was performed to investigate in vivo the effect of high-mobility group box-1 protein (HMGB1) on the maturation of dendritic cell (DC) and the influence on T-cell-mediated immunity after thermal injury. Rats were randomly divided into 3 groups as follows: sham burn group, burn group, and burn with ethyl pyruvate (EP) treatment group, and they were sacrificed on post burn days (PBD) 1, 3, 5, and 7 respectively. MACS microbeads were used to isolate splenic DCs and column of nylon wool to obtain T cells. Phenotypes were analyzed by flow cytometry and cytokines were determined with ELISA kits. The expression levels of splenic HMGB1 were significantly elevated during PBD 1–7. DC expressed similar CD80 levels, strongly enhanced CD86, and slightly elevated MHC class II levels in comparison to DC from sham-injured rats, and protein levels of IL-12 were not increased after thermal injury. Administration of EP to inhibit HMGB1 could significantly enhance expression levels of CD80, MHC class II on DC surface, and IL-12 production after burns. Simultaneously, proliferative activity and expression levels of IL-2 as well as IL-2Rα of T cell were restored. These results suggested that the excessively released HMGB1 might stimulate splenic DC to mature abnormally and down-regulate the IL-12 production, and further shifting of Th1 to Th2 with suppression of T-lymphocyte immune function following burn injury.
Introduction
D
Endogenous factors (ie, TNF-α and heat-shock proteins) that evoke DCs to mature are an important class of stimuli that might contribute to the initiation or perpetuation of an immune response against pathogens. In contrast, if these factors are released chronically and/or in the absence of infection, they could potentially contribute to the activation of self-reactive T cells and play roles in the development of autoimmunity (Gallucci and others 1999; Gallucci and Matzinger 2001). However, it seems that its response to trauma, burns, hemorrhagic shock, and microbial infection is associated with only a transient proinflammatory period followed by a more prolonged period of immune suppression (Efron and Moldawer 2003). Thus, it is speculated that there are some other factors in this procedure.
High-mobility group box-1 protein (HMGB1) is a non-histone, DNA-binding protein, which plays a critical role in regulating gene transcription (Thomas and Travers 2001). Recently, HMGB1 was identified as a late proinflammatory cytokine when extracellularly released may result in endotoxin-like lethality, local inflammation, and macrophage activation (Chen, Ward and others 2004; Sunden-Cullberg and others 2006). Circulating and cerebrospinal fluid HMGB1 concentrations are elevated in bacterial infection with severe sepsis (Angus and others 2007; van Zoelen and others 2007; Tang and others 2008). HMGB1, which is found as a late mediator of endotoxin challenge (Wang and others 1999), has triggered the subsequent search for the extra-chromosomal activities and functions of HMGB1 released by activated macrophages (Rendon-Mitchell and others 2003; Chen, Li and others 2004; Wang and others 2004), and also released from necrotic tissue (Scaffidi and others 2002; Rovere-Querini and others 2004), DCs (Dumitriu, Baruah, Valentinis and others 2005), or NK cells (Semino and others 2005). Extracellular HMGB1 has been shown to be able to provoke inflammation, to regulate the migration of monocytes (Rouhiainen and others 2004), and to contribute to DC maturation via the receptor for advanced glycation end products (RAGE) as well as induction of immune responses in vitro (Messmer and others 2004; Dumitriu, Baruah, Valentinis and others 2005; Dumitriu, Baruah, Bianchi and others 2005; Yang and others 2007; Zhu and others 2009). In addition, HMGB1 regulates neurite outgrowth by binding to RAGE (Hori and others 1995; Huttunen and others 1999).
HMGB1 acts as the second messengers and effectors throughout all steps of DC differentiation and regulates important DC functions in vitro, suggesting that its release at sites of cell injury or damage might play a role in the initiation and/or perpetuation of an immune response. However, it is not clear whether HMGB1 can induce the activation of DCs and subsequently activate T cells after burn injury in vivo. The present study was performed in order to illustrate the role of HMGB1 in DC biology in vivo.
Materials and Methods
Reagents and kits
Ethyl pyruvate (EP) and collagenase D from Clostridium histolyticum were purchased from Sigma, St. Louis, MO. Ficoll-Paque was purchased from Axis-shield Co., Norway. RPMI 1640, fetal calf serum (FCS), glutamine, penicillin, streptomycin, and HEPES were purchased from TianRunShanda Biotech Co. LTD, Beijing, China. Anti-DC (OX62) MicroBeads (IgG1, Clone OX62) was purchased from Miltenyi Biotec GmbH, Germany. Concanavalin A (Con A), Thiazolyl blue (MTT), and Triton X-100 were purchased from Sigma, St. Louis, MO. Antibodies used for flow cytometry analysis, including FITC-conjugated anti-rat MHC-II (IgG1, Clone HIS19), PE-conjugated mouse anti-rat CD80 (IgG1, κ., Clone 3H5), PE-conjugated mouse anti-rat CD86 (IgG1, κ., Clone 24F), PE-conjugated mouse anti-rat CD25 (IgG1, κ. Clone OX-39), FITC-conjugated mouse anti-rat CD3 (IgM, κ. Clone 1F4), as well as mouse anti-rat CD16/CD32 (Fcγ-β/α-R) (IgG.1b, κ. Clone D34-485) were purchased from BD/PharMingen, San Diego, CA. Mouse anti-rat RAGE monoclonal antibody (IgG2A, Clone 175410) was purchased from R&D Systems, Minneapolis, MN. FITC-conjugated goat anti-mouse IgG2A was purchased from Biosynthesis Biotechnology Co., Ltd, Beijing, China. Total RNA isolation system and reverse transcription system were purchased from Promega, Madison, WI. SYBR Green PCR Master MIX was purchased from Applied Biosystems, Foster City, CA. HMGB1 ELISA kit was purchased from Shino-Test Corporation, Kanagawa, Japan. sIL-2R ELISA kit was purchased from Market, Kanagawa, Japan. ELISA kits of IL-12, IL-2, IL-4, and IFN-γ were purchased from Biosource, Worcester, MA.
Animal thermal injury model
A widely used technique for producing full-thickness scald injury was used in these studies. Wistar rats (weight range 250–300 g), purchased from the Laboratory Animal Center, Beijing, China, were housed in separate cages in a temperature-controlled room with 12 h light and 12 h darkness to acclimatize for at least 7 days before being used. All animals had free access to water, but were fasted overnight prior to the experiment. For rats with burn injury, they were anesthetized, and the dorsal and lateral surfaces of the rats were shaved. Rats were placed on their backs and secured in a protective template with an opening corresponding to 30% of the total body surface area, and the exposed skin was immersed in 99°C water for 12 s. Sham-injured rats were subjected to all of the procedures except the temperature of the bath was of room temperature. All experimental manipulations were undertaken in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals, with the approval of the Scientific Investigation Board of the Burn Institute, Postgraduate Medical College of PLA, Beijing.
Experimental design
Ninety-six rats were randomly divided into 3 groups as follows: sham-burned group (32 rats), burn group (32 rats), and burn with EP treatment group (32 rats), and all these groups were further divided into 4 subgroups of 8 rats each, and they were sacrificed on post burn days (PBD) 1, 3, 5, and 7, respectively. In addition, 8 rats were taken to serve as normal controls (the main parameters determined in the current study were found to be highly constant within sham-injured animals, so the results were not shown). The 40 mL/kg lactated Ringer's solution was administered i.p. in all 3 groups for delayed resuscitation 6 h after the injury and 4 mL at 12, 24, 36, and 48 h after burn injury, respectively. EP was added to lactated Ringer's solution (EP 28 mM) in EP treatment group. Animals of all groups were sacrificed at designated time points, and blood as well as spleen samples were harvested aseptically to determine organ damage-related variables and levels of various cytokines. Spleen samples were divided into 2 parts, one of which was used to procure DC and T cells immediately, and the other was snap-frozen in liquid nitrogen and stored at −80°C until use for measurement of cytokine levels and total RNA extraction.
Isolation of splenic DCs
Spleens were digested by collagenase D and cut the tissue into tiny pieces. Pass the digested material through a 30-μm stainless steel mesh using a syringe and wash the Petri dish and the mesh with buffer solution. Pre-enrichment of splenocytes was performed by Ficoll-Paque density gradient centrifugation. The splenocytes were blocked with 50 μg/mL of Fc block for 15 min on ice. Splenic DCs were isolated using Anti-DC (OX62) MicroBeads and a MiniMACS™ Separator with a positive selection MS column according to manufacturer's instructions. DCs were then cultured in RPMI 1640 FCS (10%) overnight for recovery. The supernatants were collected to determine cytokine levels.
Isolation of splenic T cells
Spleens were teased in 5 mL RPMI 1640. Cells were dissociated through a 30-μm stainless steel mesh and collected after centrifugation at 300g for 10 min, and they were resuspended in 4 mL RPMI 1640. Then mononuclear cells were obtained using Ficoll-Paque density gradient centrifugation and incubated with nylon wool-packed columns for 2 h at 37°C. T cells were obtained by eluting the columns with 30–40 mL of RPMI 1640 at a flow rate of one drop per second. The cells thus obtained were 90%–95% CD3-positive T cells (data not shown). T cells with concanavalin A (Con A) stimulation were cultured for 18 h at 37°C. Supernatants were collected to determine cytokines and cells were divided into 2 portions. One (5 × 107/mL) was used to extract total RNA, and the other (5 × 105/mL) was used to detect expression levels of interleukin-2 receptor α (IL-2Rα).
Flow cytometric analysis
DCs (1 × 105) were reacted for 15 min at 4°C in 100 μL of PBS 5% FCS 0.1% sodium azide (staining buffer) with PE-conjugated IgG specific for CD80, CD86 or with FITC-conjugated IgG specific for MHC-II. In some experiments, DCs were treated with anti-RAGE monoclonal antibody followed by FITC-conjugated goat anti-mouse IgG. In all experiments, isotype controls were included using an appropriate PE- or FITC-conjugated irrelevant mAb of the same Ig class. T cells (1 × 105) were blocked with 1 μg of Fc blocker for 15 min at 4°C in 100 μL of PBS 5% FCS 0.1%, and then reacted for 20 min in darkness with PE-conjugated CD25 and FITC-conjugated CD3. Cells were then washed twice with PBS 5% FCS, fixed in 10% formaldehyde in PBS (pH 7.2–7.4), and examined by flow cytometry using a FACScan (BD Biosciences, Mountain View, CA).
T-cell proliferation assay
T cells were suspended in RPMI 1640 culture medium supplemented with 10% FCS, 100 U/mL penicillin, and 100 μg/mL streptomycin, and placed in 96-well round-bottomed plates in sextuple for proliferation, giving the final cell density of 5 × 106/mL. Cells were treated with or without 5 μg/mL Con A for 68 h at 37°C in 5% CO2/100% humidified air. Thiazolyl blue (MTT) (5 mg/mL, 10 μL/well) was then added and incubation was continued for 4 h, and 100 μg acid isopropanol was added to dissolve the MTT crystals. When all the crystals had been dissolved through the repeated blowing with a pipette, the optical density was measured by the use of a microplate reader (Spectra MR, Dynex, Chantilly, VA) at the wavelength of 540 nm.
SYBR green real-time RT-PCR
Total RNA was extracted from the splenic tissue or T cells using the single-step technique of acid guanidinium thiocyanate–chloroform extraction according to the manufacturer's instruction. The concentration of purified total RNA is determined spectrophotometrically at 260 nm. mRNA for HMGB1, IL-12p40 in splenic tissue and IL-2, IL-2Rα in splenic T cell, and GAPDH were quantified in duplicate by SYBR Green 2-step, real-time RT-PCR. After removal of potentially contaminating DNA with DNase I, 1 μg of total RNA from each sample was used for RT with an oligo-dT and a Superscript II to generate first-strand cDNA. PCR mixture was prepared using SYBR Green PCR Master Mix and using the primers as shown in Table 1. Thermal cycling conditions were 10 min at 95°C followed by 40 cycles of 95°C for 15 s and 60°C for 1 min on a Sequence Detection System (ABI PRISM 7500, USA). Each gene expression was normalized with GAPDH mRNA content. Primer software was provided to design the primers for HMGB1 (NM_002128) and IL-2Rα (NM_000417). Sequences of rat primer for SYBR Green PCR were shown in Table 1.
Cytokine measurements by ELISAs
HMGB1, IL-12, IL-2, sIL-2, IL-4, and IFN-γ were determined with ELISAs, strictly following the protocols provided by the manufacturer. The color reaction was terminated by adding 100 μL of orthophosphoric acid. Plates were read in a microplate reader (Spectra MR, Dynex, Chantilly, VA). The standard concentration curve for HMGB1 was linear from 0 to 116.8 ng/mL, for IL-12 from 0 to 2,000 pg/mL, for IL-2 from 0 to 1,500 pg/mL, for sIL-2 from 0 to 3,000 pg/mL, for IL-4 from 0 to 62.5 pg/mL, for IFN-γ from 0 to 1,400 pg/mL, respectively. The HMGB1 concentration in spleen was calculated in nanogram per milligram of protein in tissue determined using the principle of protein–dye binding (Bradford 1976).
Statistical analysis
Data were expressed as mean ± standard deviation (SD) and analyzed with a one-way analysis of variance (ANOVA). Unpaired Student's t-test was used to evaluate significant differences between groups. A P value of 0.05 or less was considered to indicate statistical significance.
Results
HMGB1 content in serum and spleen induced by burn injury
As shown in Figure 1, splenic HMGB1 gene as well as protein expressions and serum HMGB1 levels were significantly elevated during PBD 1–7, and they were markedly inhibited by treatment with EP (though they did not fall to sham-injury levels during PBD 1–3.). The peak HMGB1 levels in spleen tissues appeared on PBD 1, and in serum on PBD 3.

ELISA and SYBR green real-time RT-PCR analysis of the expression levels of splenic gene (
Phenotypic and functional changes in DCs after injury
To look for the effects of HMGB1 on splenic DC, these cells were analyzed at different time points following severe burn injury, sham burn, and EP treatment. As shown in Figure 2, splenic DC expressed similar levels of CD80, strongly enhanced levels of CD86 during PBD 1–7, and slightly enhanced levels of MHC class II on PBD 1, in comparison to DC from sham-injured rats. Treatment with EP to inhibit HMGB1 could significantly increase the expression levels of CD80, MHC class II on DC, and the expression of CD86 still kept high values, which suggested that HMGB1 might suppress the burn-induced CD80 and MHC class II expression.
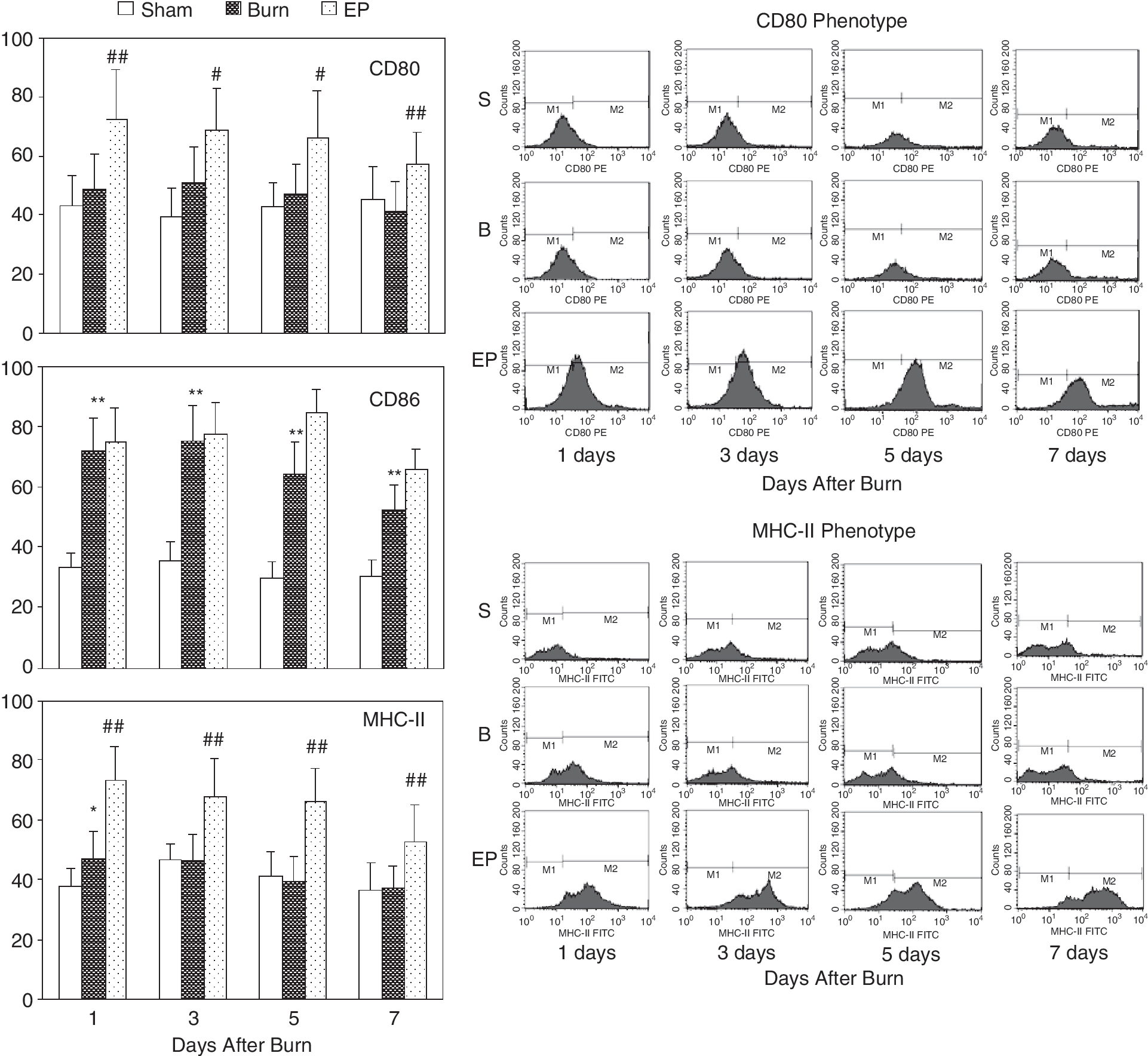
Flow cytometric analysis of phenotypes of dendritic cells (DCs). Splenic DC expressed similar levels of CD80, strongly enhanced levels of CD86 during PBD 1–7 and slightly enhanced levels of MHC class II on PBD 1 compared with DC from sham-injured rats. Treatment with ethyl pyruvate (EP) to inhibit high-mobility group box-1 protein (HMGB1) could significantly enhance the expression of CD80, MHC class II on DC, and the expression of CD86 still remained high. *P < 0.05, **P < 0.01 as burn group versus sham burn group; #P < 0.05, ##P < 0.01 as EP group versus burn group.
Levels of IL-12 in DC supernatant and in serum after injury
The capacity of DC to produce IL-12, which is one of the markers of function of mature DC and the key mediator for Th1 cell polarization, was analyzed in the present experiment. As shown in Figure 3, the expression levels of IL-12 mRNA in spleen tissue were markedly decreased in burn-injured rats in comparison with sham-injured group during PBD 1–7. Levels of IL-12 in both supernatants of DC and serum were not elevated after burn injury. Treatment with EP could significantly raise the expression levels of IL-12 mRNA in spleen tissue and levels of IL-12 production in serum as well as culture supernatants of DC.

ELISA and SYBR green real-time RT-PCR analysis of the expression levels of splenic IL-12 gene (
T-cell activation after injury
In order to understand the involvement of DC involved in the effect of HMGB1 on T-cell immune function, T-cell proliferative activity and production of cytokines were analyzed. The T-cell proliferative activities (Fig. 4A) in response to Con A in burn-injured group were significantly suppressed during PBD 1–7 as compared with sham-injured group, which could be restored by treatment with EP. Gene or protein expressions of IL-2 by T cells (Fig. 4B) and IL-2Rα on surface of T cells (Fig. 4C) were simultaneously suppressed to certain extent after burn injury, and EP treatment could restore their expressions. Conversely, serum sIL-2R levels (Fig. 4C) after burn injury were decreased at first on PBD 1, and then rapidly increased during PBD 3–7, peaking on PBD 3, and treatment with EP decreased serum sIL-2R levels only on PBD 7.

Using MTT assay for T-cell proliferation, immunofluorescence techniques for expression of IL-2Rα, ELISA for IL-2 and sIL-2R, and SYBR green real-time RT-PCR analysis for mRNA measurement. T-cell proliferative activity in response to Con A in burn-injured rats was significantly suppressed during PBD 1–7 as compared with sham-injured rats, and gene or protein expressions of IL-2 and IL-2Rα on surface of T cells were simultaneously suppressed after burn injury. Ethyl pyruvate (EP) treatment could restore T-cell proliferative activity response to Con A, gene, or protein expressions of IL-2 and IL-2Rα after burn injury. Conversely, serum sIL-2R levels (Fig. 5C) after burn injury were decreased on PBD 1 and then rapidly increased during PBD 3–7, and treatment with EP decreased serum sIL-2R levels only on PBD 7 or 5–7. Statistical significance: *P < 0.05, **P < 0.01 as burn group versus Sham burn group; #P < 0.05, ##P < 0.01 as EP group versus burn group.
Polarization of T cell after injury
Th1 cells produce IFN-γ and Th2 cells produce IL-4. Thus, using ELISA we detected the 2 cytokines produced by T cells to identify polarization of naive T cell. As shown in Figure 5, after burn injury the levels of IL-4 produced by T cell as a response to Con A were markedly increased, whereas the levels of IFN-γ markedly reduced, indicating that naive T cells develop into Th2 cells. EP treatment could significantly inhibit increased levels of IL-4 and reduced levels of IFN-γ after thermal injury, indicating that EP treatment might influence the polarization of T cells in animals subjected to thermal injury and induced naive T cells to shift to Th1 cells.
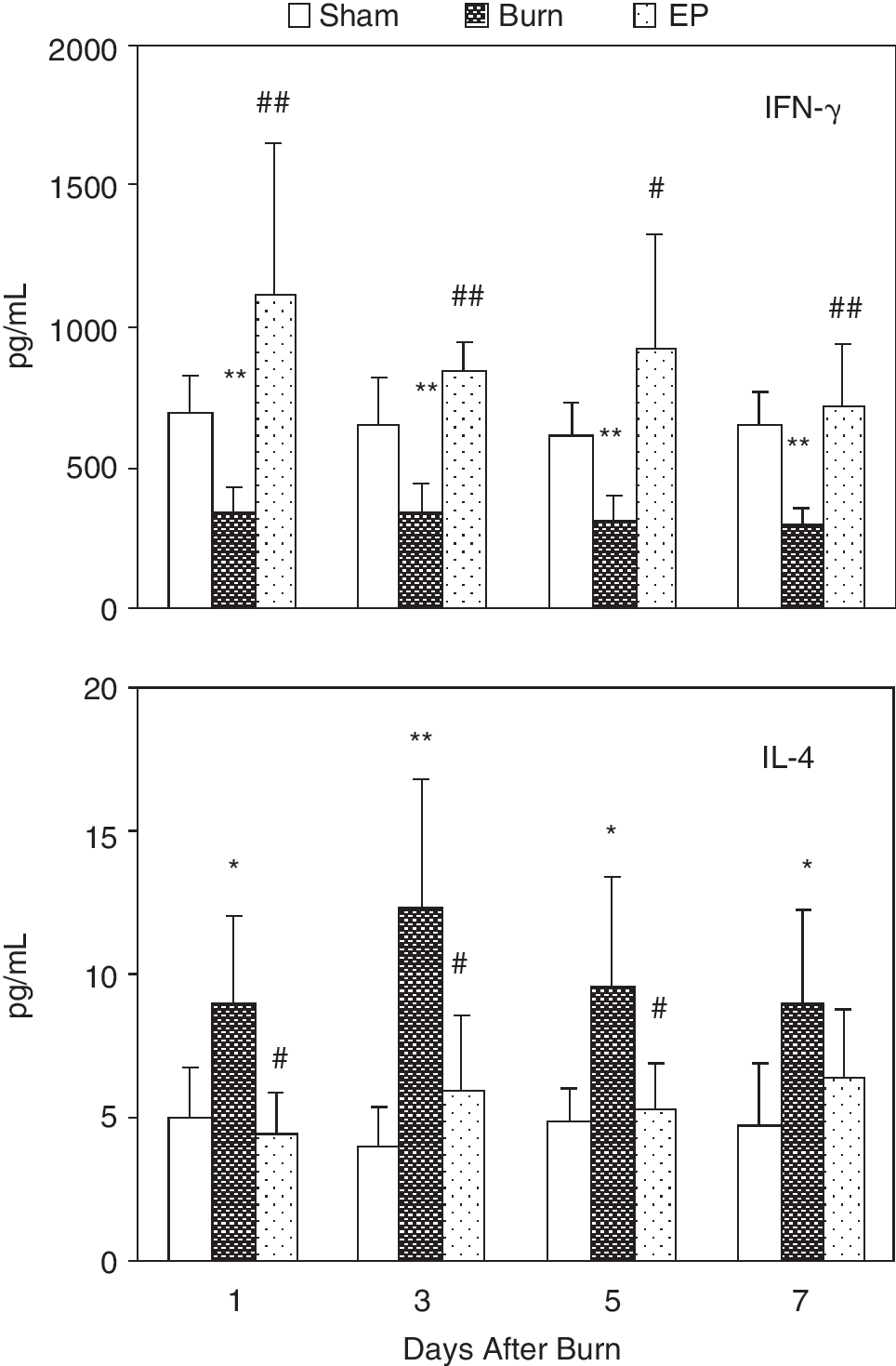
ELISA was used to determine IL-4 and IFN-γ levels in culture supernatant of T cells. After burn injury IL-4 levels produced by T cells response to Con A were markedly increased, whereas IFN-γ levels were markedly lowered. Ethyl pyruvate (EP) treatment could significantly revert elevated levels of IL-4 and lowered levels of IFN-γ after thermal injury. Statistical significance: *P < 0.05, **P < 0.01 as burn group versus sham burn group; #P < 0.05, ##P < 0.01 as EP group versus burn group.
Discussion
DCs are essential for an effective immune response to burn injury or sepsis. They are highly efficient at presenting Ag to T cells. It is well known that sepsis causes marked immune suppression. We therefore would expect that there would be a dysfunction of antigen-presenting capacity of DCs derived from burn-injured rats, similar to what has been reported for macrophages. Active DCs capture and process antigens, express co-stimulatory molecules, migrate to lymphoid organs, secrete cytokines to initiate immune responses, and activate lymphocytes (Banchereau and Steinman 1998). In the present study, we showed that DC expressed similar levels of CD80, strongly enhanced levels of CD86, and slightly enhanced levels of MHC class II in burn rats in comparison with DC from sham-injured rats. It appears that burn injury induced maturation of splenic DC with low expressions of some phenotypes. These mature DC might be a dysfunction of stimulating T cells. The cytokine secretion pattern of these DC was detected and it was found that these DC produced lower levels of IL-12 after burn injury. DCs are the major source of IL-12 (Cella and others 1996; Koch and others 1996) that is a proinflammatory cytokine that plays an important role in modulating the body's immune response and can direct T cells to differentiate into Th1 (Langenkamp and others 2000; Das and others 2001; Wu and others 2006). The inability of mature splenic DC to produce cytokines such as IL-12 maybe interpreted that there is a dysfunction of these DC. The same phenomenon was in cecal ligation and puncture (CLP)-induced sepsis (Ding and others 2004; Flohé and others 2006).
Furthermore, significantly suppressive T-cell proliferation during PBD 1–7 was found, and expression levels of IL-2 in T-cell supernatant and IL-2Rα on T-cell surface were simultaneously suppressed to certain extent. It was also found that T cells polarized to Th2 cells after burn injury. The above findings indicate that there is a marked immune suppression of T cells following burn injury (O'Riordain and others 1993; Choudhry and others 2002; Patenaude and others 2005; Phan and others 2005). To collaborate with our findings, it has been reported that DC in the “semi-mature” phenotype of MHC class II/CD86, but negative for secretion of proinflammatory cytokines, inhibit the proliferation of T cells and release of cytokines for polarization to antigen-specific Th1 cells (Flohé and others 2006). T-cell priming and initiation of adaptive immune responses require the maturation of the most potent APC, especially the DC. Thus, it is speculated that immune suppression was partly induced by dysfunction of DC after burn injury. Nevertheless, it is also possible that those effects were caused by the direct effect of HMGB1 on T cells (Zhang and others 2008).
Excessive induction of HMGB1 might result in inflammatory response producing tissue injury and organ failure (Mantell and others 2006). Our results showed that splenic and serum HMGB1 expression levels were significantly elevated during PBD 1–7, indicating that the kinetic of excessive formation of HMGB1 is delayed and prolonged. In order to verify whether the dysfunction of mature DC was associated with excessive release of HMGB1 after burn injury, EP was used to inhibit the effect of HMGB1 (Ulloa and others 2002). It was shown that administration of EP could significantly enhance the expression of CD80, MHC class II on DC surface and IL-12 production. However, T-cell proliferative activity and expression levels of IL-2 and IL-2Rα were markedly restored, and T cells were shifted to Th1. Our data confirmed delayed treatment with EP after burn injury to inhibit HMGB1, the function of mature DC and T-cell activity were restored, and T cell skewed to Th1. Therefore, the results suggest that excessive release of HMGB1 might stimulate splenic DC to mature abnormally, and further induce suppression of immune function of T lymphocyte. Our previous study, however, has showed that in vitro HMGB1 stimulation could result in marked up-regulation expressions of co-stimulatory molecules in a dosage-dependent manner, including CD80, CD86, and MHC-II on surface of rat splenic DC. Nevertheless, such response could be down-regulated with the high dosage and prolongation of treatment with HMGB1 (Zhu and others 2009). Recently, similar findings have been reported by others (Yang and others 2007). We therefore speculated that HMGB1 might have a dual regulatory effect on immune functions of DCs. The possible reasons causing the seemingly opposed functions may depend on differences in protein structure and concentration as well as its location (Yamada and Maruyama 2007). In the present experiments, the peak serum levels of HMGB1 were on PBD 3, which may be caused by release of burned necrotic skin. Because the structure of HMGB1 that is passively released by necrotic cells is different from that secreted by inflammatory cells (Gardella and others 2002; Bonaldi and others 2003), HMGB1 released by burned necrotic skin might be involved in the abnormal expressions of DC phenotype.
In summary, the excessively released HMGB1 may suppress the burn-induced CD80 and MHC class II expression in DC and IL-12 production, which leads to down-regulation of IL-2/IL-2R-dependent T-cell proliferation and shifting of Th1 to Th2.
Footnotes
Acknowledgments
This study was supported, in part, by grants from the National Natural Science Foundation (No. 30872683, No. 30971192, No. 30672178), the National Basic Research Program of China (No. 2005CB522602), and the National Natural Science Outstanding Youth Foundation of China (No. 30125020).
Author Disclosure Statement
No competing financial interests exist.