
Abstract
RNA viruses in the paramyxovirus family have evolved a number of strategies to escape host cell surveillance and antiviral responses. One mechanism exploited by a number of viruses in this family is direct targeting of cytokine-inducible transcription regulators in the STAT family. Diverse members of this large virus family effectively suppress STAT signaling by the actions of their V proteins, or the related proteins derived from alternate viral mRNAs. These viral proteins have distinct means of targeting STATs, resulting in a variety of negative effects on STATs and their signal transduction. Recent developments in understanding STAT targeting will be reviewed.
Interferon Signal Transduction and Antiviral Innate Immune Responses
I
The importance of the STAT-mediated IFN-signaling systems in mediating antiviral defense is highlighted by the fact that many viruses have evolved mechanisms to evade activation of this innate immune response. For some viruses, including the paramyxoviruses, the early steps of virus detection and IFN induction are targeted by inhibitory mechanisms including dsRNA sequestration or signaling interference to antagonize IRF3 and NF-κB pathways. In other cases, virus-encoded soluble IFN receptors or receptor antagonists block cytokine signaling. Individual viruses can also block specific antiviral effectors to preserve key cellular machinery needed for their replication. One host evasion mechanism common to many paramyxoviruses is evasion of IFN signaling by direct interference with the IFN-inducible STAT proteins. The molecular mechanisms of STAT-directed IFN evasion are known to be very diverse amongst individual viruses in this family, and most have been reviewed in detail elsewhere (Gotoh and others 2002; Horvath 2004a, 2004b). In this article, more recent developments in understanding STAT targeting by different paramyxoviruses are highlighted.
Paramyxoviruses
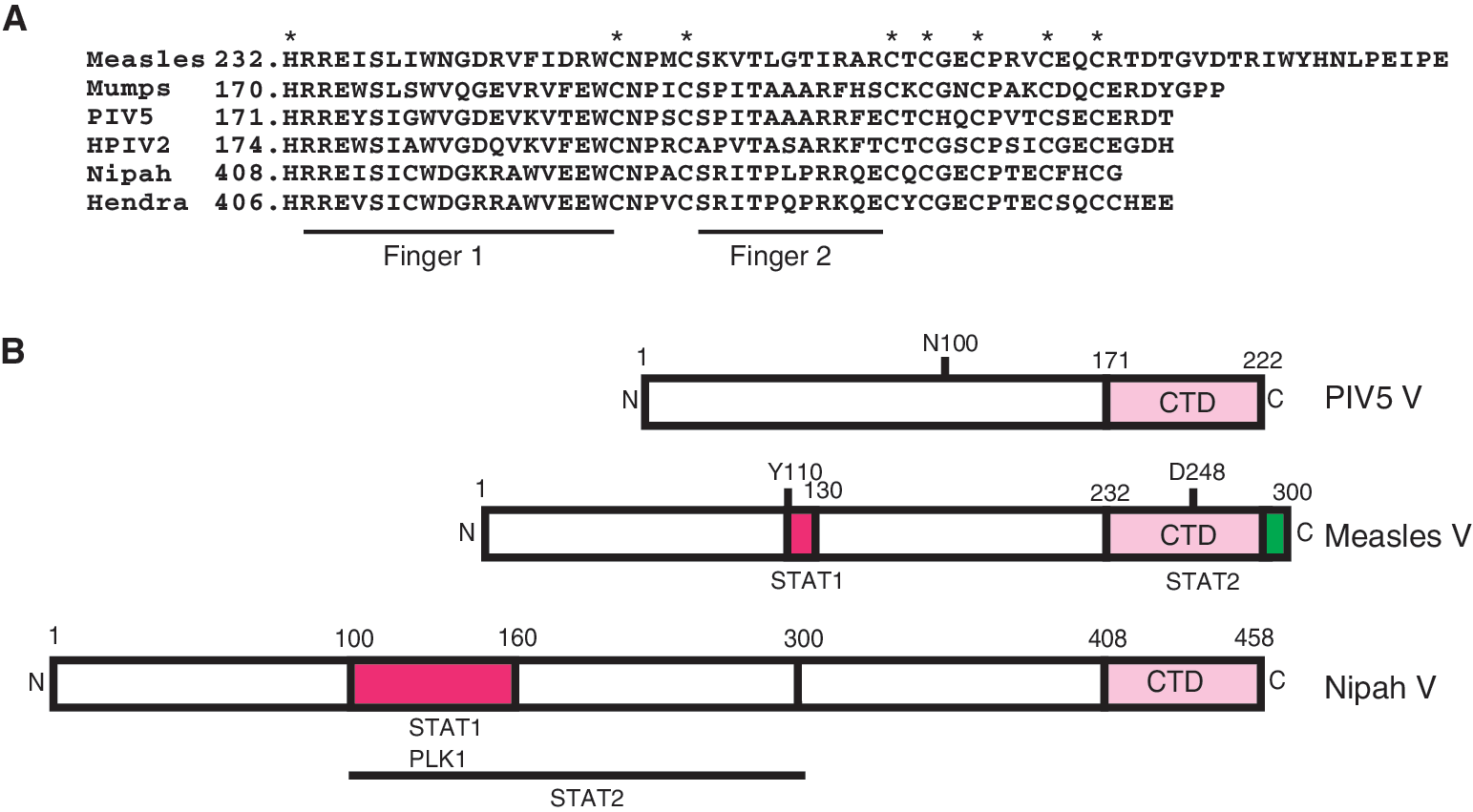
The paramyxoviruses are enveloped negative strand RNA viruses that are subdivided into several genera including Respirovirus (eg, Sendai virus), Rubulavirus (eg, PIV5, HPIV2, mumps), Avulavirus (eg, Newcastle disease virus), Morbillivirus (eg, measles virus), and henipaviruses (eg, Hendra and Nipah virus). All of these viruses share common structural, biochemical, and genetic elements including the single-stranded RNA genome that encodes a small number of proteins, including surface glycoproteins and several subunits of an RNA-dependent RNA polymerase (reviewed in Lamb (2001)). A noteworthy property found in all the paramyxoviruses is the ability of its P gene to code for more than 1 protein (Thomas and others 1988). Sendai virus P gene, for example, directs the expression of at least 7 proteins, including P, V, W, C′, C, Y1, and Y2. Other paramyxoviruses express fewer but more than 1 protein from their P/V/C gene, either from overlapping open-reading frames accessed by alternate translation initiation or by a unique mechanism referred to as RNA editing that produces alternate mRNAs by site-specific cotranscriptional insertion of non-templated nucleotides (Thomas and others 1988). As a result of the RNA editing phenomenon, the P locus produces proteins that share a common N terminus but differ in their C-terminal domain (CTD). Most commonly, this process produces 2 proteins called P and V, but in some viruses the third reading frame can produce a protein referred to as W (Harcourt and others 2000).
The V protein CTD is highly conserved among the paramyxoviruses (Fig. 1A). The CTD of the V protein is cysteine-rich and binds 2 zinc (Zn) molecules per V protein (Liston and Briedis 1994; Paterson and others 1995). A diverse range of host evasion activities, including IFN-signaling inhibition (Didcock and others 1999), prevention of apoptosis (He and others 2002; Wansley and Parks 2002), cell cycle alterations (Lin and Lamb 2000), inhibition of double-stranded RNA signaling (He and others 2002; Poole and others 2002), and prevention of IFN biosynthesis (He and others 2002; Poole and others 2002; Wansley and Parks 2002), have been specifically ascribed to the paramyxovirus V proteins. The other proteins derived from this locus, including C and W, have also been implicated in evading IFN responses and antiviral signaling. In most cases, a fundamental activity associated with the host evasion proteins is in direct interference with STAT protein function by mechanisms that rely on protein interactions, but it is recognized that individual genera within the family exhibit remarkably distinct mechanisms of STAT inhibition (Fig. 1B).
Distinct mechanism of STAT inhibition by different paramyxoviruses. (
Rubulaviruses
The Rubulavirus genus includes parainfluenza virus 5 (PIV5, formerly known as SV5), mumps virus, and the type 2 human parainfluenza virus (HPIV2). In this genus the V protein is encoded by the collinear transcript; it is the product of the unedited mRNA. Insertion of 2 non-templated guanine nucleotides produces an mRNA encoding the P protein (Thomas and others 1988). The PIV5 V protein was the first to be recognized as an IFN-signaling inhibitor, and sole expression of this protein in human cells results in rapid and specific loss of STAT1 by a proteasome-mediated degradation (Didcock and others 1999). In fact, all of the Rubulavirus V proteins have been shown to efficiently target STAT proteins for proteasome-mediated destruction with remarkable targeting specificity. The PIV5 V protein targets STAT1, the HPIV2 V protein targets STAT2 (although in some cases can also target STAT1), and the mumps virus V protein targets both STAT1 and STAT3 (Didcock and others 1999; Goodbourn and others 2000; Young and others 2000; Kubota and others 2001; Parisien and others 2001; Parisien and others 2002a, 2002b; Yokosawa and others 2002; Ulane and others 2003). While IFN signal transduction is not required for STAT targeting, somatic cell genetic analysis has demonstrated that both PIV5 V and mumps V protein require the cellular expression of STAT2 in order to target STAT1, while HPIV2 requires STAT1 to target STAT2. This requirement restricts the host range of PIV5, as the divergent murine STAT2 fails to associate with PIV5 V protein, and cannot support targeting of STAT1 (Parisien and others 2002a). A single amino acid N100 in the PIV5 V protein has been shown to provide species-specific interaction with human STAT2 (Young and others 2001). The requirement for human STAT2 in STAT1 destruction provides the first example of an intracellular signaling and transcription factor that functions as a host range determinant for any virus. This observation was further validated by the creation of a human STAT2 transgenic mouse model system in which human STAT2 was ubiquitously expressed in an immune-competent intact mouse (Kraus and others 2008). The presence of human STAT2 in this transgenic mouse enabled targeting of the murine STAT1 protein, and resulted in specific interference with IFN antiviral responses only in infected cells. This resulted in increased virus replication in the lungs of the humanized mouse, and created an enhanced cytokine and ISG induction profile in the infected animals.
Biochemical studies of STAT targeting by Rubulavirus V proteins indicates that they achieve STAT degradation by coordinating the assembly of a multicomponent E3 ubiquitin ligase complex that includes a number of cellular proteins including DDB1, the cullin family member Cul4A, and both STAT1 and STAT2 (Lin and others 1998; Andrejeva and others 2002; Ulane and Horvath 2002). This V-mediated targeting complex is referred to as VDC, an acronym for V-dependent degradation complex, which fortuitously also denotes its central components, V, DDB1, and Cul4A (Fig. 2A). DDB1 is an essential component of the VDC, required for targeting STATs by all the Rubulavirus V proteins (Precious and others 2005; Ulane and others 2005). Data indicate that V proteins form separate interactions with their STAT substrates and the cellular ubiquitin ligase components. Furthermore, it has been observed that the V proteins can homo- or hetero-oligomerize through their highly conserved C-terminal domain (CTD) and form macromolecular spherical particles visible in the electron microscope. Based on these facts, it has been suggested that the V protein particle acts as an enzymatic scaffold for combining the hijacked E3 ubiquitin ligase activities with the V-associated STAT substrate, resulting in highly efficient targeting (Ulane and others 2005). The obligatory role of STAT2 is to act as a substrate adaptor, bridging STAT1 and the DDB1 complex (Precious and others 2005; Ulane and others 2005). Irrespective of the structural considerations, the Rubulavirus V protein acts as an efficient catalyst for the degradation of STAT1 to prevent the establishment of the IFN-induced antiviral state in the cells (Carlos and others 2005; Precious and others 2007).
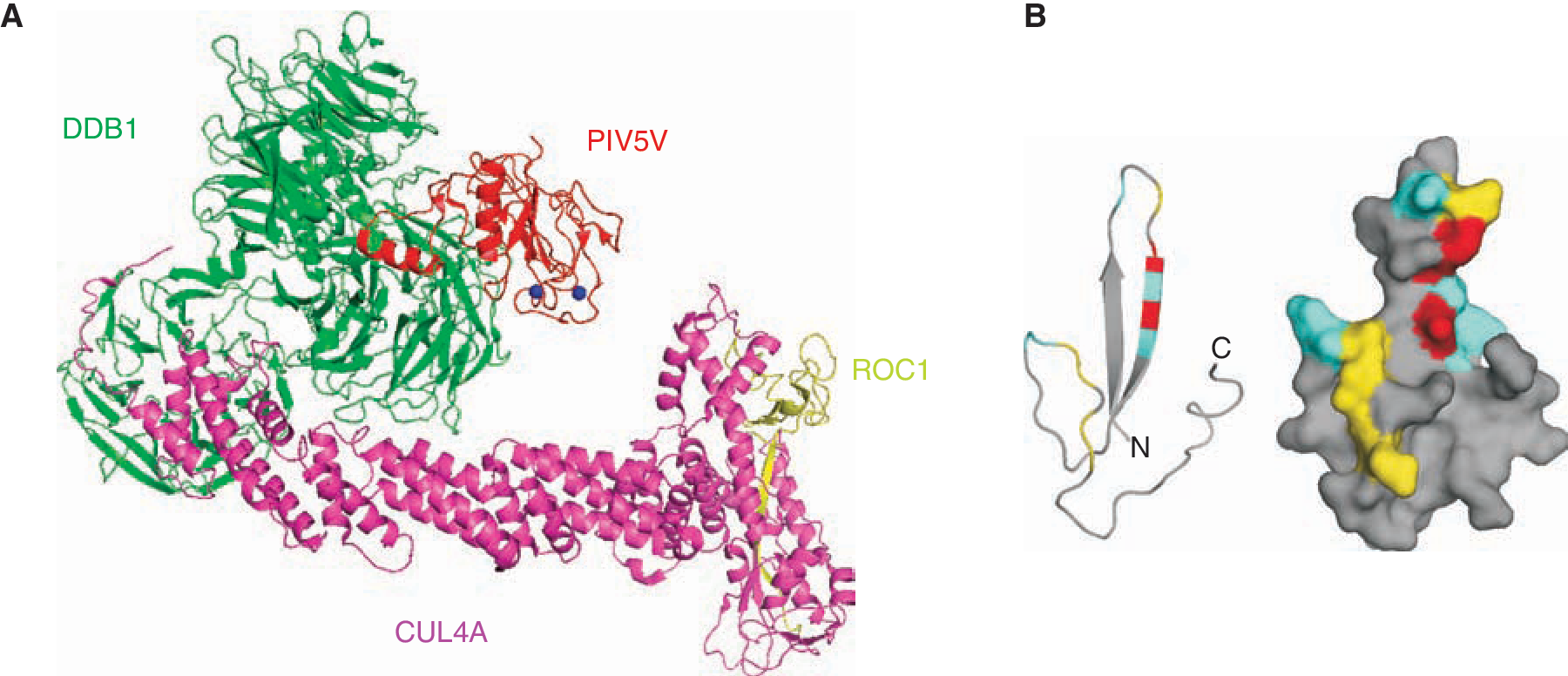
Structural representations of paramyxovirus V proteins. (
Structural studies strongly support the molecular and biochemical conclusions regarding the Ub ligase complex assembled by Rubulavirus V proteins. Crystallographic visualization of PIV5 V protein in complex with DDB1 revealed molecular details about the V protein and its association with DDB1 (Li and others 2006) (Fig. 2A). This study showed that a unique zinc finger is formed that involves the first histidine of the V CTD, and 7 cysteine residues. The structure also demonstrated the importance of both N and C terminus in interacting with DDB1 confirming the previous work that STAT1 can be degraded by only full-length protein of PIV5 V (Andrejeva and others 2002) and that only V and not P protein can participate in targeting STAT1 (Didcock and others 1999). The data are also consistent with the findings that the C-terminal cysteine cluster is important for DDB1 binding (Lin and others 1998; Andrejeva and others 2002). In addition, a series of tryptophans with characteristic spacing (W-X3-W-X9-W) has also been shown to be important for STAT protein interactions with mumps and HPIV2 V proteins (Nishio and others 2002; Nishio and others 2005a).
Based on the structural and genetic features of the rubulaviruses, a new paramyxovirus, mapeura virus, has been recently isolated from bats. But unlike the rubulaviruses, this virus evades the IFN response by sequestering STAT proteins and preventing their nuclear translocation rather than degrading STATs (Hagmaier and others 2007). Another Rubulavirus, HPIV4, has been shown to be completely unable to evade the IFN response (Nishio and others 2005b). Sequence comparison reveals different amino acid composition in the N and C terminus of the virus V protein. However, as cell type and species-specific factors are already known to strongly influence the outcome of V-mediated STAT destruction, further work is needed to better understand the nature of these viruses, and their abilities to interface with the host IFN response and STAT proteins.
Henipaviruses
Hendra virus and Nipah virus are the 2 prototype species in the emerging Henipavirus genus that was recently identified and causes lethal diseases in humans. In the henipaviruses, the P protein is derived from the collinear transcript, and addition of non-templated guanine nucleotides produces mRNAs encoding 2 additional proteins, V and W with different carboxyl termini (Harcourt and others 2000). In addition, the C protein is produced from an alternate translational site within the P gene. Comparison of the mRNA editing frequency of the P gene of henipaviruses with other viruses has revealed that henipaviruses produce edited transcripts at twice the frequency of other viruses (Kulkarni and others 2009; Lo and others 2009); although the biological significance, if any, of this enhanced editing is not clear, it may lead to greater capacity for host immune evasion. Immunoflouresence analysis of Nipah virus-infected cells with specific antiserum has demonstrated that the P protein localizes throughout the cell, the C protein accumulates in the cytoplasm in a punctate pattern, and the V protein accumulates exclusively in cytoplasm while the W protein is predominantly in the nucleus. This steady state picture of protein accumulation does not give justice to the dynamics of nucleocytoplasmic movement, as it is established that the Henipavirus V protein shuttle between nucleus and cytoplasm with net nuclear export due to the presence of a potent export signal (Rodriguez and others 2004; Lo and others 2009). The C, V, and W proteins of Nipah virus inhibit virus transcription and RNA replication when expressed in a minigenome replicon assay (Sleeman and others 2008) suggesting that these proteins share similar functional properties.
Henipavirus P, V, and W proteins have been implicated in blocking the host IFN response by targeting cellular host proteins. Nipah virus P, V, and W proteins all form high-affinity interactions with STAT1 through their common N-terminal domain (Rodriguez and others 2004; Shaw and others 2004), and this activity maps to residues 100–160, which is also present and functional in Hendra virus (Fig. 1B). Additional residues as well as the presence of STAT1 are required for these proteins to associate with STAT2 (Rodriguez and others 2004). The Henipavirus V proteins inhibit IFN responses by sequestering STAT1 and STAT2 in high-molecular-weight cytoplasmic complexes, thereby preventing their IFN-mediated tyrosine phosphorylation (Rodriguez and others 2002; Rodriguez and others 2003). The short CTD of the Nipah virus W protein possesses a potent nuclear localization signal (NLS) that interacts with host karyopherin alpha 3 and 4 (Shaw and others 2005), giving rise to it net nuclear accumulation. It has also been shown that W protein but not V protein can inhibit TLR3-mediated signaling due to loss of activated IRF3, but the mechanism of TLR3 signaling inhibition by W protein remains uncharacterized (Shaw and others 2005). Recently another cellular protein, Polo-like kinase 1 (PLK1), has been identified as a binding partner for Henipavirus P, V, and W proteins. The PLK1-binding site overlaps with the STAT1-binding site on the P, V, and W proteins (Ludlow and others 2008), as it maps between residues 100–160 of their shared N terminus (Rodriguez and others 2002). Mutagenic evaluation of PLK1 and STAT1 binding to Henipavirus V protein has revealed that Ser 130 and Ser 131 are both needed for binding to STAT1 (and therefore STAT2) and normal IFN-signaling interference (Ludlow and others 2008). The interaction of PlK1 with V and W are independent of the STAT interactions suggesting V:STAT and V:PLK exist as separate complexes, but the precise biological significance of PLK1 association with Henipavirus V proteins remains to be determined. A therapeutically relevant outcome of this study was the demonstration that Nipah virus P proteins that no longer interact with STATs or PLK1 retain the ability to function in minigenome replication assays. As defective STAT1 interference is an obvious strategy for attenuation of henipaviruses, this result suggests that reverse genetics approaches will be successful in creation of vaccine strains. Indeed it was earlier found that a Nipah virus V protein with a single amino acid substitution fails to engage STAT1 (Hagmaier and others 2006).
Morbilliviruses
Measles virus is the prototype of the Morbillivirus genus that infects millions of people each year despite the availability of effective vaccines. For measles, the P mRNA is derived from the collinear transcript, and both V and C proteins are generated from RNA editing or translational mechanisms. The P/V/C locus of measles virus, like other paramyxoviruses, is associated with host immune evasion, and can disengage IFN signaling. The evasion activities are ascribed to the V protein, but specific cases of P- and C-mediated host evasion have been revealed (Devaux and others 2007; Fontana and others 2008), although it has been reported that V proteins from either attenuated or wild-type strains are more potent inhibitors of IFN-inducible reporter gene expression than C proteins (Fontana and others 2008). The C protein has also been implicated as a regulator of viral RNA synthesis (Nakatsu and others 2008) and shown to be important for virus spread in infected macaque lymphatic tissue (Takeuchi and others 2005). Other morbilliviruses such as rinderpest virus and canine distemper virus also block type I and type II IFN through their structural and nonstructural proteins (Nanda and Baron 2006; von Messling and others 2006).
The mechanism of measles V protein-mediated inhibition of IFN signaling is distinct from other paramyxoviruses. The measles virus V protein does not degrade STATs, but effectively prevents IFN-induced STAT1 and STAT2 nuclear import (Palosaari and others 2003) by an unknown mechanism. Measles virus V protein can efficiently antagonize IFN-α/β signaling but the IFN-γ signaling inhibition has not always been confirmed (Palosaari and others 2003; Takeuchi and others 2003; Caignard and others 2007; Devaux and others 2007; Fontana and others 2008). Recently, distinct means for disengaging the IFN-γ and IFN-α/β signal transduction machinery through separate associations with STAT1 and STAT2 have been revealed (Fig. 1B). Results indicate that STAT2 is the primary target for measles V protein-mediated IFN-α/β-signaling evasion. Measles virus V protein engages STAT2 in the absence of STAT1 and in the absence of the V protein's STAT1-binding site (Ramachandran and others 2008). Unlike other paramyxovirus V proteins, measles V engages STAT2 specifically through the highly conserved CTD, and this domain is both necessary and sufficient for interaction with STAT2 and IFN-α/β-signaling evasion. Results demonstrate that both the zinc finger structure and specific amino acids present on the fingers are required for mediating interactions with STAT2 (Fig. 2B). Similar findings by other groups were also reported, not only verifying the importance of the CTD for STAT2 interaction, but also implicating conserved tryptophan residues W240 and W250 in mediating STAT2 interaction (Caignard and others 2009). The direct and fundamental association with STAT2 clearly represents a target for design of a small molecule that could disengage the measles virus IFN-signaling evasion, perhaps creating an alternative therapy for measles outbreaks. This would be complicated however, as a distinct region of the molecule associates with STAT1. The STAT1-binding site on measles V has been mapped to residues 110–130 in the region shared between P and V proteins (Ramachandran and others 2008; Caignard and others 2009). This region is noted for high sequence conservation amongst morbilliviruses (Devaux and others 2007). This region originates with tyrosine 110 that has been shown to be important in the P protein-mediated block of STAT1 tyrosine phosphorylation (Devaux and others 2007). Interestingly, this residue was first implicated as an attenuating mutation in the original measles virus infectious clone, Ed-TAG, that was shown to be defective in IFN evasion (Radecke and others 1995; Combredet and others 2003; Ohno and others 2004). Subsequent reports of studies using the Edmonston ATCC strain, Moraten, Schwartz, or CAM70 strains confirmed the importance of Y110 in STAT1 interference (Caignard and others 2007; Devaux and others 2007; Fontana and others 2008; Ramachandran and others 2008). It is clear from our study and prior investigations (Takeuchi and others 2003) that measles virus V protein more effectively achieves IFN-α/β signal interference than IFN-γ interference. Both P and V have the intrinsic capacity to engage STAT1 via residues 110–130, consistent with the finding that infectious measles virus lacking V protein expression retains the ability to antagonize STAT1 signaling via the P protein in a Y110-dependent reaction (Devaux and others 2007). It is likely that the differential effects of measles virus on IFN-γ signaling may become more apparent during its normal life cycle in whole animals, and can only be revealed by the study of animal models or in the context of natural measles virus infections with wild-type strains.
Conclusions
The mechanisms of STAT-directed IFN-signaling evasion by the biomedically important paramyxovirus family has been a paradigm for the study of host innate immune evasion by viruses, and has revealed that fascinating diversity can be achieved by virus evolution in accomplishing the single task of overcoming host antiviral responses. These studies have provided further insight into the molecular underpinnings of virus replication, and provided targets for therapeutic intervention and support for novel attenuation strategies. While much has been learned in a short time, we are confident that the ongoing study of these small RNA viruses will yield more novel insights into the mechanics of innate immunity, and continue to provide amusement to the dedicated scientists who are passionately intrigued by their diversity.
Footnotes
Acknowledgments
The authors acknowledge that the large family of viruses reviewed here is matched by an equally large number of investigators who are committed to their study. Unfortunately limitations on space and time prevent us from mentioning them comprehensively, and we apologize for any who might feel left out of this general review. C.M.H. is particularly grateful for the long-term advice and mentoring from Dr. Robert A. Lamb, who discovered the V proteins in his laboratory in 1988, and for the contributions of Dr. Richard Randall, who first noted the disappearance of STAT1 following infection of cells with PIV5. Work on paramyxovirus interactions with STAT signaling in the Horvath laboratory is supported by NIH grant AI050707 to C.M.H.