
Abstract
The full-length coding sequence of chicken interferon-γ (ChIFN-γ) was cloned into a baculovirus nonfusion vector, pFastBacDual, and expressed in Sf21 insect cells. Recombinant ChIFN-γ (rChIFN-γ) protein was found to be expressed both intracellularly as well as in the culture supernatants. The affinity-purified rChIFN-γ contained 14, 17, and 28 kDa proteins, possibly representing both glycosylated and nonglycosylated protein forms of ChIFN-γ. The bioactivity of rChIFN-γ was confirmed in vitro by production of nitric oxide in a chicken macrophage cell line (HD11) and antiviral activity against vesicular stomatitis virus in primary chicken embryonic fibroblast cells. Further, HD11 cells stimulated with rChIFN-γ showed significant upregulation of inducible nitric oxide synthases, IFN-γ, interleukin-1β, interleukin-12p35, signal transducers and activators of transcription 1, class II, major histocompatibility complex, transactivator, and major histocompatibility complex II-β chain (BL-B) transcripts. In conclusion, the present study provides information on the ability of functionally active rChIFN-γ expressed in a baculovirus system in inducing significant transcriptional upregulation of various immune system-related genes, including those that encode cytokines, antigen-presenting molecules, and transcription factors involved in the major histocompatibility complex and IFN-signaling pathway.
Introduction
I
Considering the central role of IFN-γ in mediating effective immune responses against pathogens, there has been a considerable interest in the use of this cytokine as a vaccine adjuvant. In this regard, IFN-γ may be administered in the form of a plasmid DNA expressing this cytokine or as a purified recombinant protein. Recombinant chicken IFN-γ (rChIFN-γ) has been produced using different expression hosts, including bacteria, animal cells, viruses, T cell hybridoma, plants, brevibacillus, and baculoviruses (Weining and others 1996; Lowenthal and others 1997; Song and others 1997; Lillehoj and Choi 1998; Lambrecht and others 1999; Michalski and others 1999; Schijns and others 2000; Koji and others 2001; Takehara and others 2002; Li and others 2010). However, the baculovirus expression system seems to overcome some of the disadvantages of other recombinant systems, such as lipopolysaccharide contamination of the purified protein, undesirable post-translational modification, nonspecific cleavage, shorter half-life, and insufficient bioactivity associated with using other systems (King and Possee 1992; Lambrecht and others 1999; Shrestha and others 2008).
Here, we report the expression of a functionally bioactive rChIFN-γ using the baculovirus expression system. Importantly, we demonstrate the ability of rChIFN-γ to induce several immune system-related genes in chicken macrophages.
Materials and Methods
Cells and viruses
The baculovirus nonfusion vector pFastBacDual (Invitrogen) was used as the cloning vector, whereas Spodoptera frugiperda (Sf21) cells were used for infection and rChIFN-γ expression. Cells were grown in Sf900 medium (Invitrogen) supplemented with penicillin and streptomycin (100 U/mL). Primary chicken embryo fibroblast (CEF) primary cells were prepared from 11-day-old chicken embryos for the in vitro antiviral assay, whereas the chicken macrophage cell line-HD11 was used for the in vitro nitric oxide (NO) production assay and gene expression studies.
Synthesis of chicken IFN-γ cDNA
Complementary DNA encoding chicken IFN-γ was synthesized from concanavalin A-stimulated chicken splenocytes as per the method described previously (Parvizi and others 2009). Briefly, splenocytes from a 4-week-old SPF chickens were stimulated with concanavalin A (5 μg/mL; Sigma) and incubated for 72 h at 41°C in a humidified incubator with 5% CO2 pressure. After cell lysis, mRNA was extracted using RNeasy-micro mRNA purification kit (Qiagen) and the cDNA was synthesized using the First-Strand cDNA synthesis kit (Invitrogen) as per the manufacturer's instructions. Gene-specific primers were designed with suitable restriction sites and 1xFLAG, an octapeptide fusion tag (AspTyrLysAspAspAspAspLys) to amplify the full-length coding sequence of ChIFN-γ (forward: 5′-GCGCAG
Cloning, expression, and purification of ChIFN-γ
Coding sequences of chicken IFN-γ gene were cloned into the multiple cloning site of pFastBacDual between NheI and KpnI restriction sites and the plasmid construct was transformed into DH5-α competent cells by electroporation. The positive clones were confirmed by polymerase chain reaction (PCR) amplification of the ChIFN-γ gene followed by sequencing. The recombinant baculovirus expression cassette pFastBacDual-ChIFN-γ was used for generating a bacmid by site-specific transposition mediated by baculovirus shuttle vector with a mini-attTn7 target site and a helper plasmid, followed by transformation into Escherichia coli DH10Bac host. The positive transformants were screened by PCR, and recombinant bacmid DNA encoding the rChIFN-γ constructs was transfected into Sf21 cells to generate recombinant baculovirus expressing rChIFN-γ. The presence of recombinant baculovirus expressing rChIFN-γ in Sf21 culture was determined by observing cell cytopathic effects and Western blotting. Further, rChIFN-γ was purified from the Sf21 cell lysates and culture supernatants by affinity chromatography as per the manufacturer's instructions (Sigma). The purity of the recombinant protein was confirmed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and Western blot analysis using anti-FLAG antibodies. A parallel set of mock purification was performed using the culture supernatant of uninfected Sf21 cells and used as negative control for subsequent in vitro bioassay experiments.
Assessment of antiviral activity and production of NO
To determine the antiviral activity of rChIFN-γ, primary CEF cells (5 × 104/well) were treated overnight with rChIFN-γ followed by vesicular stomatitis virus (VSV)-green fluorescent protein (GFP) (multiplicity of infection [MOI] of 0.1) in serum-free α-minimum essential medium Eagle (MEM) for 1 h, as described previously (Skellick and Marcus 1986) with modification (Askhar and others 2008). The GFP fluorescence intensity was measured 24 h later on a Typhoon Trio (GE Healthcare) and the signals were quantified using the ImageQuant™ 5.2 software. PolyI:C, a known Toll-like receptor (TLR)3 agonist with anti-VSV activity, was used as a positive control and a TLR ligand that has no anti-VSV activity (short double-stranded RNA) or medium alone was used as a negative control.
Bioactivity of rChIFN-γ was assessed by measuring the amount of nitrite (NO2 −) in the culture supernatant of HD11 macrophage cell line as described by (Lowenthal and others 1995). Briefly, HD11 cells were cultured at l × 105 cells/well and aliquots of 50 μL of 2-fold serial dilutions of rChIFN-γ protein (1 μg to 20 ng/mL) were added to each of the triplicate wells and incubated for 48 h. The presence of NO in the culture supernatant was determined by measuring the nitrite (NO2 −) using Griess reagent as per the manufacturer's instructions (Promega).
Cytokine gene expression in HD11 cells stimulated with rChIFN-γ
Expression of several immune-related genes in HD11 cells treated with rChIFN- γ was quantified by real-time PCR using Lightcycler LC480 (Roche Diagnostics GmbH), as described by Abdul-Careem and others (2006). These genes included IFN-γ, interleukin-1β (IL-1β), IL-12p35, inducible NO synthases (iNOS), signal transducers and activators of transcription 1 (STAT1), class II, major histocompatibility complex, transactivator (CIITA), and chicken MHC Class II β-chain (B-LB). The gene-specific primer sequences are given in Table 1. Total RNA was extracted from the cells collected at 2, 12, and 24 h poststimulation, and cDNA was synthesized using Superscript First-Strand cDNA Synthesis kit (Invitrogen). Gene expression was calculated relative to the expression of the β-actin house-keeping gene using the Pfaffl's formula (Pfaffl 2001). The average relative expression of each gene at a given time point was compared between stimulated and control cells using the Student's t-test.
BL-B, MHC Class II β-chain; CIITA, class II, major histocompatibility complex transactivator; F, forward primer; IFN, interferon; IL, interleukin; iNOS, inducible nitric oxide synthases; R, reverse primer; STAT1, signal transducers and activators of transcription 1.
Results
Cloning, expression, and purification of rChIFN-γ
Analysis of the full-length coding sequence of ChIFN-γ cloned into pFastBacDual was found 100% identical to the previously published sequence (GenBank accession no. NM 205149). The sequence obtained in the present study was also closely related to another published sequence (GenBank accession no. U272645), with one synonymous mismatch at position 292. Electrophoretic and Western blot analysis of the purified preparations of the culture supernatants of Sf21 cells infected with recombinant baculovirus expressing rChIFN-γ revealed 3 bands with approximate molecular weights of 14, 17, and 28 kDa (Fig. 1A–C).

Antiviral response of rChIFN-γ in primary CEF cells
The ability of rChIFN-γ to induce an antiviral response was evaluated utilizing the VSV-GFP IFN bioassay. The findings showed that rChIFN-γ-treated CEF cells displayed a strong resistance to infection by VSV, as determined by a marked reduction in the cytopathic effects to the levels comparable to that induced by polyI:C (Fig. 2A). These results were further supported by the observation that untreated or mock TLR ligand-treated CEF cells displayed no protection from VSV infections. The inhibitory concentration of rChIFN-γ to induce its anti-VSV activity was found to be as low as 100 ng/mL (Fig. 2B).
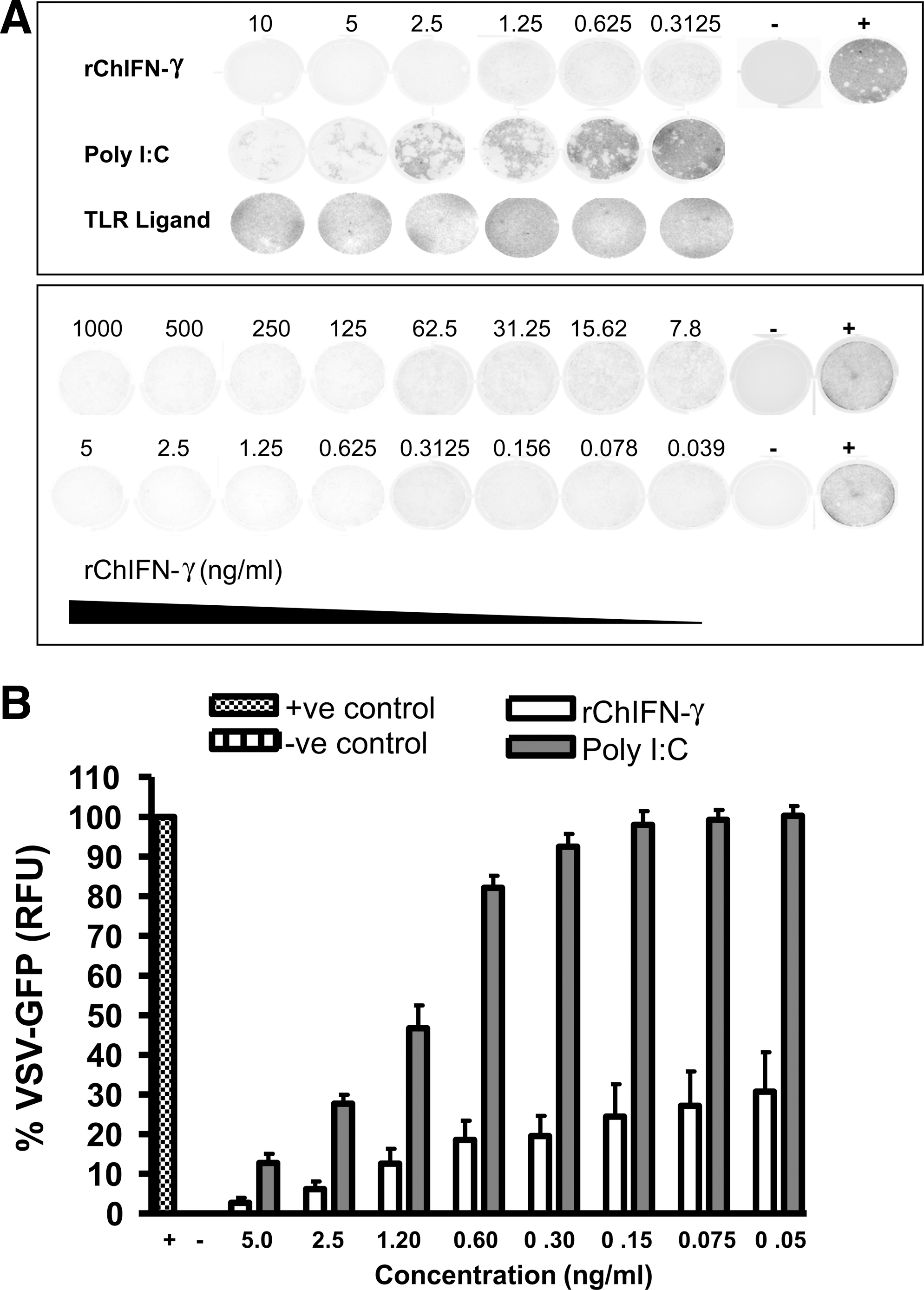
Antiviral activity of rChIFN-γ against VSV virus. Bioactivity is inversely proportional to the amount/intensity of GFP fluorescence as measured by Typhoon Trio scanning. Primary chick embryo fibroblast (CEF) cells treated with a serial dilution of rChIFN-γ were infected with GFP-tagged vesicular stomatitis virus (VSV)-GFP at 50% TCID.
In vitro NO production assay
Culture supernatants collected from rChIFN-γ-treated HD11 cells were analyzed for NO production. Treatment with rChIFN-γ was shown to induce a significant increase in the NO production at a concentration of as low as 13 ng/mL of rChIFN-γ (Fig. 3). The increase in NO production in HD11 cells in response to treatment by rChIFN-γ was found to be dose dependent.
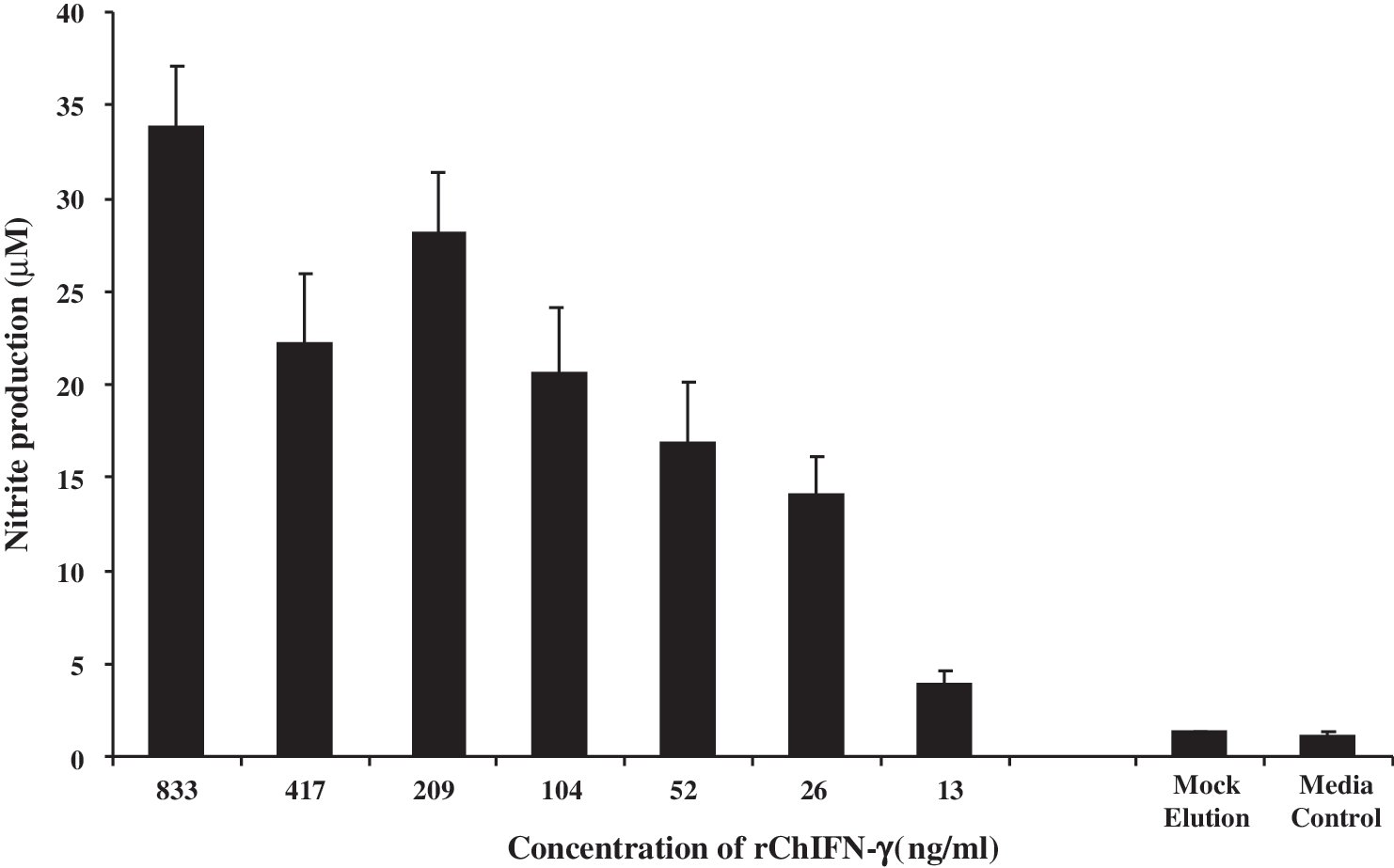
Dose-dependent increase in NO2 − in chicken macrophage culture supernatants. Chicken macrophage cells (HD11) were treated with various concentrations of rChIFN-γ, mock elution, or medium for 48 h, and culture supernatants were collected and assayed for nitrite with Griess reagent.
Relative expression of cytokine genes in rChIFN-γ-stimulated HD11 cell line
Real-time reverse transcriptase (RT)-PCR was performed to determine the relative expression of IFN-γ, IL-1β, IL-12p35, iNOS, STAT1, BL-B, and CIITA genes in rChIFN-γ-stimulated HD11 cells. There was significant upregulation of several gene transcripts at 2, 12, or 24 h poststimulation (Fig. 4A–G). Gene expression of IFN-γ, IL-1β, IL-12p35, iNOS, BL-B, and CIITA was found significantly upregulated at both 12 and 24 h poststimulation. In addition, IFN-γ and IL-12p35 transcripts were significantly upregulated as early as 2 h poststimulation (Fig. 4A, C). Transcription of STAT1 gene was also significantly upregulated at 12 h poststimulation (Fig. 4D).
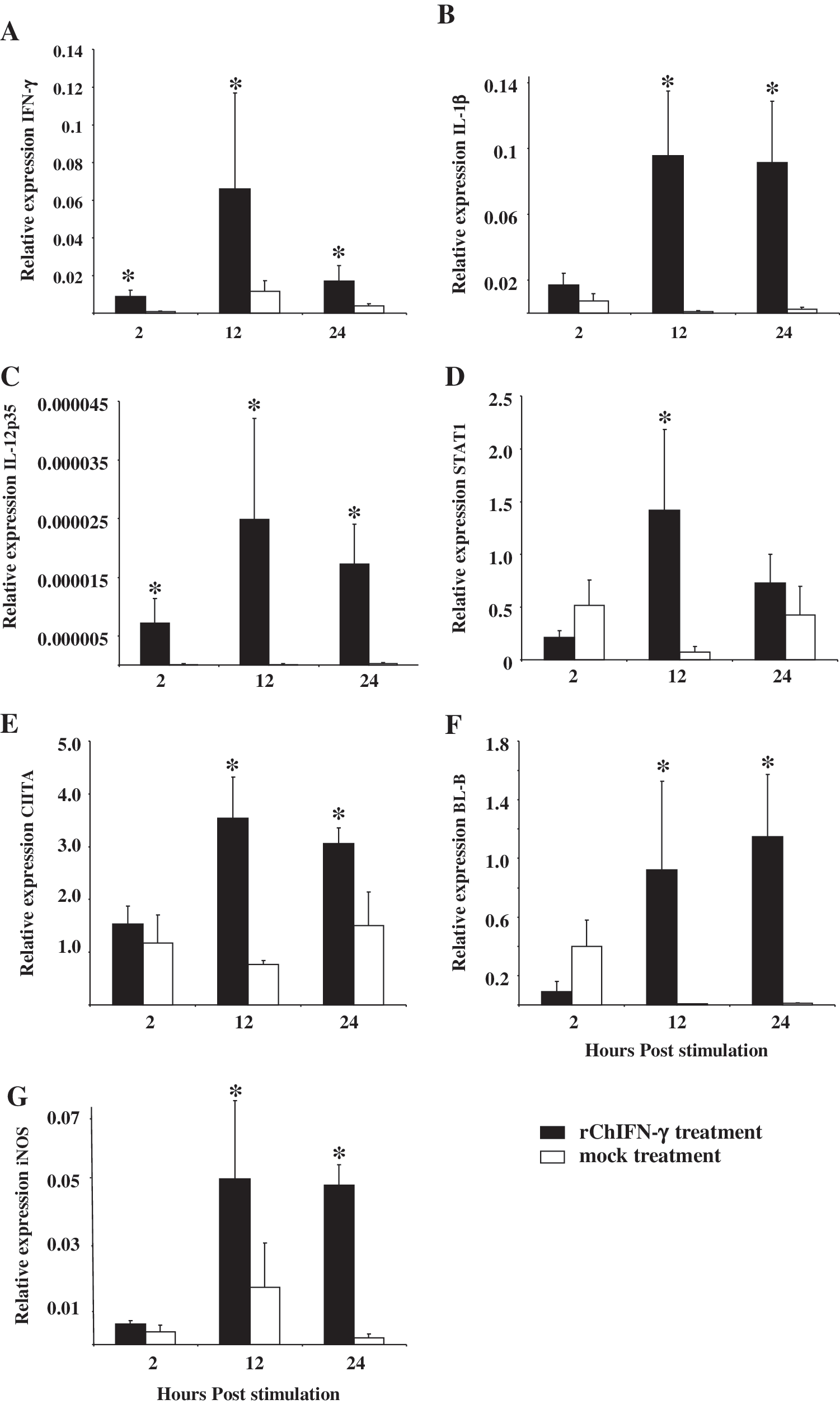
Relative expression of
Discussion
The present study demonstrated the successful cloning and expression of ChIFN-γ in a baculovirus expression system and its ability to induce a marked, dose-dependent NO production as well as induction of antiviral activities in chicken cells. In particular, the effect of rChIFN-γ on transcriptional activation of cytokines, transcription factors, and molecules involved in antigen presentation was investigated. Findings from this study provide novel information on the downstream effects of IFN-γ in cultured chicken macrophage cells treated with this cytokine.
To date, there have been a few other studies that have reported expression of rChIFN-γ using both prokaryotic and eukaryotic expression systems (Song and others 1997; Lambrecht and others 1999). The findings from these studies are in agreement that, compared to prokaryotic expression systems, rChIFN-γ molecules expressed in eukaryotic expression systems, such as the baculovirus system, have an increased biological half-life and superior bioactivity in chicken cells.
In the present study, rChIFN-γ was expressed in the baculovirus expression system in both soluble and insoluble fractions. Electrophoretic analysis of rChIFN-γ revealed 2 bands of 14 and 17 kDa sizes with traces of a 28 kDa band. On the basis of the findings of others, it is likely that the 14 and 17 kDa proteins were the glycosylated and nonglycosylated forms, respectively, and the 28 kDa protein was a homodimer (Song and others 1997; Lambrecht and others 1999). Although there is some suggestion that glycosylation may improve the protein half-life and prevent its post-translational modifications (Sareneva and others 1993), the role of glycosylation in the biological activity of chicken IFN-γ remains unknown and needs further investigation (Takehara and others 2002).
The importance of NO production by phagocytes in response to infection has been well documented (Lambrecht and others 1999; Takehara and others 2002). In this context, we observed a dose-dependent increase in NO production in HD11 macrophages after stimulation by rChIFN-γ and this observation was further supported by a significant increase in iNOS gene transcription at 12 and 24 h poststimulation with rChIFN-γ in these cells. Although NO production in chicken cells in response to IFN-γ stimulation has been reported previously (Digby and Lowenthal 1995; Weining and others 1996; Li and others 2010), our findings also demonstrated the association between transcriptional upregulation of iNOS genes and NO production in chicken macrophages. Further, in agreement with other reports (Takehara and others 2002; Wu and others 2009), the antiviral activity of rChIFN-γ was markedly evident in the IFN-bioassay against VSV using CEF cells. rChIFN-γ was found to induce a potent anti-VSV activity at concentrations as low as 100 ng/mL, suggesting that rChIFN-γ expressed in the present study was bioactive.
A novel aspect of the present study was its examination of immune system gene expression profiles in cells treated with IFN-γ. These genes included those encoding cytokines, transcription factors, and molecules involved in antigen presentation. This study was designed in such a manner to capture temporal changes in the expression of these genes. On the basis of the data presented here, it appears that gene expression induced by recombinant IFN-γ in chicken macrophages is tightly controlled in that it is induced as early as 2 h, peaks at 12 h, and either subsides or plateaus by 24 h post-IFN-γ treatment.
Previously, Liu and others (2004) developed an in silico statistical model to predict cytokine-regulated transcription of genes by scoring the transcription factor binding sites on the putative promoters of regulated versus nonregulated genes derived from the microarray and quantitative PCR data of IFN-γ-treated human macrophages. This model predicted that 1,387 genes are potentially regulated by IFN-γ, of which genes encoding molecules involved in antigen presentation (eg, MHC-II, CIITA, and β2 microglobulin), IFN signaling molecules (eg, STAT1, IRF2), and cytokines (eg, IL-12p40) were of note. However, the effect of IFN-γ on temporal transcriptional activation of immune system genes in chicken macrophages remains largely unknown. To this end, we observed a significant increase in the transcription of IFN-γ and IL-12p35 genes at all time-points after rChIFN-γ stimulation in HD11 cells, a chicken macrophage cell line, suggesting a rapid and sustained ability of IFN-γ to induce T helper 1-(Th1) like responses in chicken macrophages. Induction of endogenous IFN-γ transcription in these macrophages stimulated with exogenous rChIFN-γ suggested that this cytokine can also act in a positive feedback manner to stimulate further expression of IFN-γ. This is consistent with the recent work by Cheeseman and others (2008). In addition, the increase in IFN-γ transcription in these cells was supported by a significant upregulation in the gene expression of a transcription activator involved in IFN signaling, STAT1. The IFN-γ-induced IL-12 expression seems to act in a positive feedback loop that represents an important amplifying mechanism in immune responses against intracellular pathogens. The priming effect of IFN-γ for IL-12 expression has been shown to occur predominantly at the transcriptional level (but not translational or post-translational levels) for both p40 and p35 genes in murine macrophages (Ma and others 1996). However, whether a similar mechanism also exists in chicken cells needs to be further investigated. The Th1/Th2 dichotomy or type I/type II response polarization has been described recently in chickens (Göbel and others 2003; Kaiser and others 2003; Parvizi and others 2009). Therefore, based on the aforementioned observations, it is conceivable that rChIFN-γ is able to activate macrophages efficiently and can skew the tissue microenvironment to a type I phenotype. This Th1-like response may in turn assist CD8+ T cell differentiation and expansion, leading to conferring immunity against intracellular pathogens, particularly viruses.
Another hallmark of effective activation of innate defense mechanisms is the upregulation of proinflammatory cytokines and particularly IL-1β, which has various functions, including regulation of immune response, participating in acute phase reactions, and hematopoiesis (Morgan and others 2004). In this context, a significant increase in the IL-1β transcription in HD11 cells after rChIFN-γ stimulation shows the ability of IFN-γ in mediating amplification pathways important in the production of inflammatory mediators. This notion is further supported by a marked increase in NO production and a significant transcriptional upregulation of iNOS genes in HD11 cells post-rChIFN-γ stimulation. The present finding of IFN-γ-induced IL-1β production in chicken macrophages is in agreement with a previous study in murine macrophage cell lines (Martin and Dorf 1990).
An important role of IFN-γ in inducing expression of antigen-presenting molecules such as MHC class II in a variety of murine and human cell types, including phagocytes, has been previously reported (Steeg and others 1982; Glimcher and Kara 1992). Presentation of peptide antigens by phagocytic cells to B and T cells plays a pivotal role in the induction of antigen-specific effector immune responses. In this context, we observed a significant upregulation in the transcription of BL-B (MHC-II beta chain) gene in chicken macrophages stimulated with rChIFN-γ. This observation is in agreement with similar studies that showed IFN-γ-induced expression of MHC class II molecules in chicken fibroblasts (Song and others 1997) and macrophages (Weining and others 1996). More importantly, increased expression of BL-B gene in the present study was supported by a significant transcriptional upregulation of CIITA, a trans-activator that is essential for the expression of many genes involved in antigen presentation (Chang and others 1996). To our knowledge, this is the first report that describes upregulation of CIITA in chicken cells in response to stimulation by rChIFN-γ.
In summary, the findings of the present study demonstrated successful cloning and expression of ChIFN-γ in a baculovirus expression system. Also, the results showed that rChIFN-γ was functionally active, as characterized by induction of NO production and antiviral resistance in chicken cells. Further, our investigations revealed significant transcriptional upregulation of various immune system-related genes in HD11 cells treated with rChIFN-γ. Experiments are now underway to use rChIFN-γ as a possible vaccine adjuvant in chickens to enhance immune responses against viral infections.
Footnotes
Acknowledgments
Funding for this project was provided by the Poultry Industry Council, Saskatchewan Chicken Industry Development Funds, and NSERC-CRD Partnerships program. Dr. R.R. Kulkarni is a recipient of an Ontario Ministry of Research and Innovation Postdoctoral Fellowship.
Author Disclosure Statement
No competing financial interests exist.