
Abstract
Immunoregulatory activity of type I interferons (IFNs) and estrogen is convergent in some cases of autoimmune disorders. The aim of our study was to determine whether a potent interaction of IFN and estradiol (E2) has an influence on immune response and estrogen receptor alpha (ER-α) expression in antigen-presenting cells in a model of experimental autoimmune orchitis (EAO). C3H/He/W male mice were immunized with testicular germ cells (TGCs) and orally treated with interferon tau (IFN-τ), E2, or both simultaneously. The delayed-type hypersensitivity reaction was intensified after the administration of either IFN-τ or E2, but their co-administration had no effect. IFN-τ treatment increased immunoglobulin G2a (IgG2a) and decreased IgG1 levels of TGC-specific antibodies, whereas E2 abolished the effects of the used cytokine. The total splenic cellularity and the number of spleen CD11c+MHC II+ and F4/80+MHC II− cells were increased after IFN-τ treatment, whereas E2 antagonized this effect. After IFN-τ administration the level of ER-α was significantly higher in F4/80+MHC II− cells, whereas E2 had no effect. However, the administration of E2 significantly reduced the ER-α level in F4/80+MHC II+ and CD11c+MHC II+ cells in comparison with the IFN-τ–treated groups. In the EAO model, the type I IFN and E2 cooperated at the general and cellular levels of immune response, but E2 treatment usually abolished the effects exerted by the cytokine.
Introduction
E
Type I interferons (IFNs) are cytokines known for their antiviral action, but they can also regulate various other immune mechanisms. Moreover, type I IFNs are used in the therapy of some autoimmune diseases. Similar to E2, IFNs can increase or decrease the intensity of autoimmune diseases, and surprisingly, the effect of IFNs is convergent with the effect of E2 in a given disorder. In the case of both multiple sclerosis (MS) and CIA (van Holten and others 2004; Goodin 2005), type I IFN improved the clinical picture, whereas SLE tended to get worse after the administration of type I IFN (Ramanujam and others 2009) and MG can even be evoked after the administration of IFN (Stübgen 2009). These observations suggest that there may be a connection between estrogen and the IFN intracellular signaling system. Taking into consideration the role of APCs in regulation of the immune response under the influence of E2, we speculate that the connection between E2 and IFN should be apparent especially in dendritic cells (DCs) and macrophages (Mφ).
The effects of estrogen are mediated via interaction of the hormone with nuclear or membrane receptors (Maggiolini and Picard 2010). There is evidence that mainly estrogen receptor alpha (ER-α), one of the estrogen-specific nuclear receptors, is responsible for the effects of estrogenic compounds on the immune response, especially in the context of autoimmunity (Polanczyk and others 2003; Elloso and others 2005; Zhu and others 2007; Feng and others 2010). ER-α is expressed in APCs, and in DCs it is indispensable for their differentiation and estrogen-dependent activity (Douin-Echinard and others 2008). It also takes part in the regulation of cytokine production by APCs (Suzuki and others 2007a; Kawasaki and others 2008). The expression of ER-α is regulated by estrogen itself and the level of ER-α protein may be associated with the effect of estrogens on cell function (Webb and others 1992). Interferon tau (IFN-τ), one of the type I IFNs produced during the preimplantation period by ruminant embryos, can influence the expression of ER-α in ovine uterine cells (Spencer and Bazer 1996; Fleming and others 2001). Moreover, oral IFN-τ exerts beneficial effects in the course of autoimmune disorders, such as experimental allergic encephalomyelitis (EAE) (Soos and others 1997, 2002) and diabetes in NOD mice (Sobel and others 2008). The effects of IFN-τ are also observable in the case of fetal loss in abortion-prone CBAxDBA/J combination (Chaouat and others 1995).
It is suspected that in leukocytes IFN-τ exerts similar activity, because it binds to the classical IFN receptors IFNAR1 and IFNAR2 and activates similar, but not identical, intracellular pathways (Subramaniam and others 1995). On the other hand, E2 can influence the signaling pathways of type I IFNs, especially by regulating the expression of transcription factors involved in IFN signal transduction (Dai and others 2009) or IFN target genes (Panchanathan and others 2009). Thus, we hypothesize that there is a relation between estrogens and IFNs at both the cellular and systemic levels of the immune response.
The aim of our study was to investigate the possible connections between estrogens and IFN-τ in a model of experimental autoimmune orchitis (EAO) in mice. We chose this model of inducing the autoimmune response because of the natural low and stable systemic level of estrogen that is characteristic in males. This allowed us to modulate the level of estrogen after oral administration (Gordon and others 1986) and to exclude the influence of the female cycle. Autoimmune response to testicular germ cell (TGC) antigens was not studied in the context of type I IFNs and estrogens. This article presents the data concerning the interactions between E2 and IFN-τ in the context of the overall immune response during autoimmune orchitis. Additionally, taking into consideration the functional activity of ER-α in APCs, we assessed the level of this receptor and the number of these cells in the spleens of experimental animals.
Materials and Methods
Animals
C3H/He/W and C57Bl6/J male mice housed in specific pathogen-free conditions were purchased from Maria Skłodowska-Curie Memorial Center and Institute of Oncology in Warsaw, Poland, at the age of 6 weeks. All procedures were approved by the local ethical committee in Wrocław, Poland.
Cell culture media
Estrogen and IFN effects were investigated in cultures with a hormone-deficient medium. DCs were cultured in an RPMI-DC medium [RPMI 1640 without phenol red (Sigma-Aldrich), 10% charcoal-treated fetal calf serum (FCS; Invitrogen), 2 mM
Generation of bone marrow-DC and bone marrow-Mf
DCs and Mφ for an in vitro experiment were generated from the bone marrow (BM) of C57Bl/6 mice, and DCs for DC-aided immunization were obtained from BM of C3H/He/W mice according to the method described by Lutz and others (1999). Briefly, BM cells were obtained by washing out long bones of leg of the mice under sterile conditions into RPMI 1640 (Sigma-Aldrich) supplemented with 5% FCS. The cells were seeded at a density of 2×106 cells in 10 mL of RPMI-DC medium per 100-mm Petri dish at day 0. The medium was supplemented with E2 (30 and 200 pg/mL), and for each concentration of E2, IFN-τ (1×105 AUV/mL) was added. The concentration of IFN-τ added to cell culture was experimentally established on the basis of cytotoxic effects. On days 6 and 8, half of the medium was replaced. On day 10, nonadherent cells were harvested and stained for flow cytometry. The purity of generated cells was between 70% and 80%, and the ratio of mature to immature cells was about 5:1.
Mφ were generated in DMEM-Mφ according to Tadokoro and Abrahamsohn (2001) but with modifications. A total of 4×106 BM cells were seated in a 100-mm cell culture dish (Nunc) in 10 mL of medium. On day 3, an additional 10 mL of the medium was added. On days 6 and 8, the total volume of the medium with nonadherent cells was discarded and 10 mL of fresh medium was added to the dishes. On day 10, the medium was aspirated with nonadherent cells. Adherent cells were treated with proper combinations of E2 and IFNs for 48 h. The purity of generated cells was above 90% and the proportion between mature and immature cells was 3:1.
Model of EAO, administration of E2 and IFN, and delayed-type hypersensitivity measurement
Mice were immunized with TGCs according to the protocol described by Itoh and others (1991) with modifications. Briefly, aseptically collected testes were gently pressed through a 40-μm plastic mesh, centrifuged at 400 g for 5 min, washed 3 times in cold phosphate-buffered saline (PBS), and counted using trypan blue dye cell exclusion. Mice were immunized 3 times at 2-week intervals with 1×107 TGCs and 1×106 syngeneic BM-DCs in 0.15 mL of PBS subcutaneously. Control mice were injected with 1×106 syngeneic BM-DCs in 0.15 mL PBS. E2 (5 μg/mouse) and IFN-τ (1×105 antiviral activity unit (AVU)/mouse) suspended in 0.2 mL of PBS were administered per os via a stomach tube daily for 5 days each week. The other animals from the immunized and control groups were given PBS in the same way. As E2 was dissolved in ethanol at 1 mg/mL, PBS and IFN-τ given to mice were supplemented with 0.01% ethanol.
On day 41 the footpad thickness was measured with a micrometer and animals from all groups were challenged by a 50 μL injection of TGC suspension (1×106 in PBS) in both hind footpads. After 24 h the footpad thickness was measured and delayed-type hypersensitivity (DTH) reaction intensity for each mouse was calculated as the difference in the mean of both footpad thicknesses at 24 h after and before the injection. Then, the animals were sacrificed and blood, spleens, and testes were collected.
Splenocyte suspension preparation
Splenocytes were isolated by pressing the spleens through a plastic mesh into cold 0.84% ammonium chloride solution. After red blood cell lysis (2 min), cell suspensions were centrifuged and washed twice in PBS. The cells were counted and 1.5×106 cells were used for flow cytometric analysis.
Flow cytometry
Surface antigens were double-stained with anti-CD11c-PE (Becton Dickinson), anti-F4/80-PE (AbD Serotec), and anti-MHC class II (Ab/Ad)-FITC antibodies (AbD Serotec) in DCs and Mφ, whereas spleen cells were stained with anti-F4/80-PE (eBioscience), anti-CD11c-PE (Becton Dickinson), and anti-MHC class II (Ak) (Becton Dickinson) for 30 min at 4°C. Appropriate isotype controls were used for each staining. After washing with PBS, the cells were fixed in 2% buffered formaldehyde (15 min, 4°C) and washed with permeabilization buffer [PB: 0.15% Triton X-100 (Sigma-Aldrich), 2% FCS, and 0.1% sodium azide (Sigma-Aldrich)]. Intracellular ER-α–specific immunostaining was prepared with rabbit immunoglobulin (IgG) anti-ER-α antibody (Santa Cruz Biotechnology) (20 μg/mL) or with normal rabbit IgG (Santa Cruz Biotechnology) as an isotype control in the same concentration diluted in PB. The cells were incubated for 75 min at 4°C. After washing with PB, the cells were stained with secondary goat anti-rabbit APC antibody (Jackson ImmunoResearch) at 1:50 dilution in PB (60 min, 4°C), then washed in PB, and fixed in 1% buffered formaldehyde solution. Stained cells were examined by flow cytometry on a Becton Dickinson FacsCalibur flow cytometer. The relative ER-α level was calculated as mean fluorescence intensity (MFI) based on the difference between the median fluorescence intensity of the ER-α–stained cells and the isotype-matched control stained cells gated for the populations of interest.
RNA isolation
About 1×106 Mφ were centrifuged and the pellet was resuspended in 500 μL of Trizol reagent (Invitrogen). Isolation was performed according to the manufacturer's instructions.
ER-α mRNA real-time polymerase chain reaction analysis
First-strand cDNA was synthesized using the SuperScript III First-Strand Synthesis System (Invitrogen) and a gene-specific primer (5′-CAGCTCCACTTCAGCACA-3′). The relative amount of murine ER-α m-RNA was determined by quantitative real-time PCR (RT-PCR) using an iQ5 Optical System (BioRad) with the iQ SYBR Green Supermix (BioRad) as appropriate. GAPDH was used as a reference gene. The primers used were as follows: S-ER-α (5′-GCTACGTCAAGTCGGTTCC-3′), AS-ER-α (5′-AAGGTGCTGGACAGAAACG-3′), S-GAPDH (5′TCACTGCCACCCAGAAGA-3′), and AS-GAPDH (5′-TACCAGGAAATGAGCTTGAC-3′). The reactions were performed under the following conditions: first cycle of 94°C for 2 min, 35 cycles of 94°C for 30 s and 58°C for 30 s, followed by 72°C for 1 min. The specificity of the PCR was determined by melt-curve analysis for each reaction.
PCR array system
The responses of 84 type I IFN-dependent genes were studied with RT2 Profiler PCR Array System (SABiosciences, Cat. No. PAMM-016) according to the manufacturer's instructions. This PCR array enables the analysis of expression of type I IFNs and their receptors, signaling molecules of the type I IFN system, genes containing IFN-responsive elements, and genes connected with IFN resistance. Data were analyzed with the recommended RT2 Profiler PCR Data Analysis software accessible online at the manufacturer's Web site (
Histology
Immediately after sacrificing the mice, the testes were fixed in 5% formalin solution, washed for 24 h in tap water, dehydrated in an alcohol series, cleared in methyl benzoate and xylene, and then embedded in paraffin. The paraffin blocks were sliced in a Micron HM310 microtome into 6-μm sections. The paraffin sections were stained with hematoxylin and eosin and observed under a light microscope (Nikon Eclipse 80I).
TGC-specific and estrogen enzyme-linked immunosorbent assay
Relative levels of anti-TGC antibodies were measured in the sera of examined animals. TGCs isolated from the testes were suspended in PBS at a concentration of 2×106 cells/mL. The 96-well flat-bottomed plates were coated with 100 μL of TGC suspension in PBS and incubated for 2 h at 4°C. Adherent cells were fixed with methanol (Sigma-Aldrich) at 37°C. After methanol evaporation and drying, the plates were frozen at−20°C until use. The sera of immunized and control animals were diluted 1:20 in PBST [PBS and 0.05% Tween 20 (Sigma-Aldrich)] and incubated on plates (1.5 h, 37°C). After washing, anti-murine IgG1, IgG2, IgG2b+IgG3, and IgA+IgM antibodies conjugated with horseradish peroxidase (Santa Cruz Biotechnology) were added and incubated for 2 h at room temperature. After a final wash with PBST, 200 μL of TMB liquid substrate (Sigma-Aldrich) per well were added and plates were incubated in the dark at room temperature for 10 min. The reaction was stopped with 0.5 M H2SO4, and after 30 min, the absorbance was read at 450 nm.
Serum estradiol concentration was measured by competitive enzyme-linked immunosorbent assay using DRG estradiol-sensitive ELISA (DRG Instruments GmbH) according to manufacturer's procedure.
Statistical analysis
Statistical analysis was performed by 1- or 2-way analysis of variance (ANOVA). For the post hoc test, all pairwise comparisons were calculated with the Tukey honestly significant difference (HSD) method. Results with a P value lower than 0.05 were considered statistically significant. Data analysis was conducted in “R” statistical environment v. 2.7.0. The results are presented as mean±standard error of the mean.
Results
The influence of E2 and IFN-τ was investigated in the model of EAO in 2 independent experiments. In the first experiment, all animals were treated only with DCs, but without TGC (DC treated). DCs were used as a kind of natural adjuvant, because C3H/He/W substrain mice seem to be more resistant to immunization with TGC alone in contrast to C3H/He mice, originally used by Itoh and others (1991). Such immunization resulted in the production of specific antibodies without the development of a DTH reaction) and histological changes (data not shown). The mice treated with syngeneic BM-DCs (DC-treated) were divided into 4 groups: fed with PBS only, with E2, with IFN-τ, or simultaneously with E2 and IFN-τ (n=10 mice per group). In the second experiment, all DC-treated mice were immunized with TGC (TGC-treated) and 1 additional group was treated only with DCs, as a control for immunization. Immunized animals were fed as in the first experiment (n=10 per group). Data were analyzed by 2-way ANOVA for each experiment independently.
Disturbed testicular morphology in TGC immunized mice was not improved after E2 and IFN-τ treatment
Histological examination of testicular tissues from the control group showed closely packed seminiferous tubules containing the germinal epithelium at all stages of maturation from spermatogonia to spermatozoa. The histological view demonstrated regular tissue (Fig. 1a).
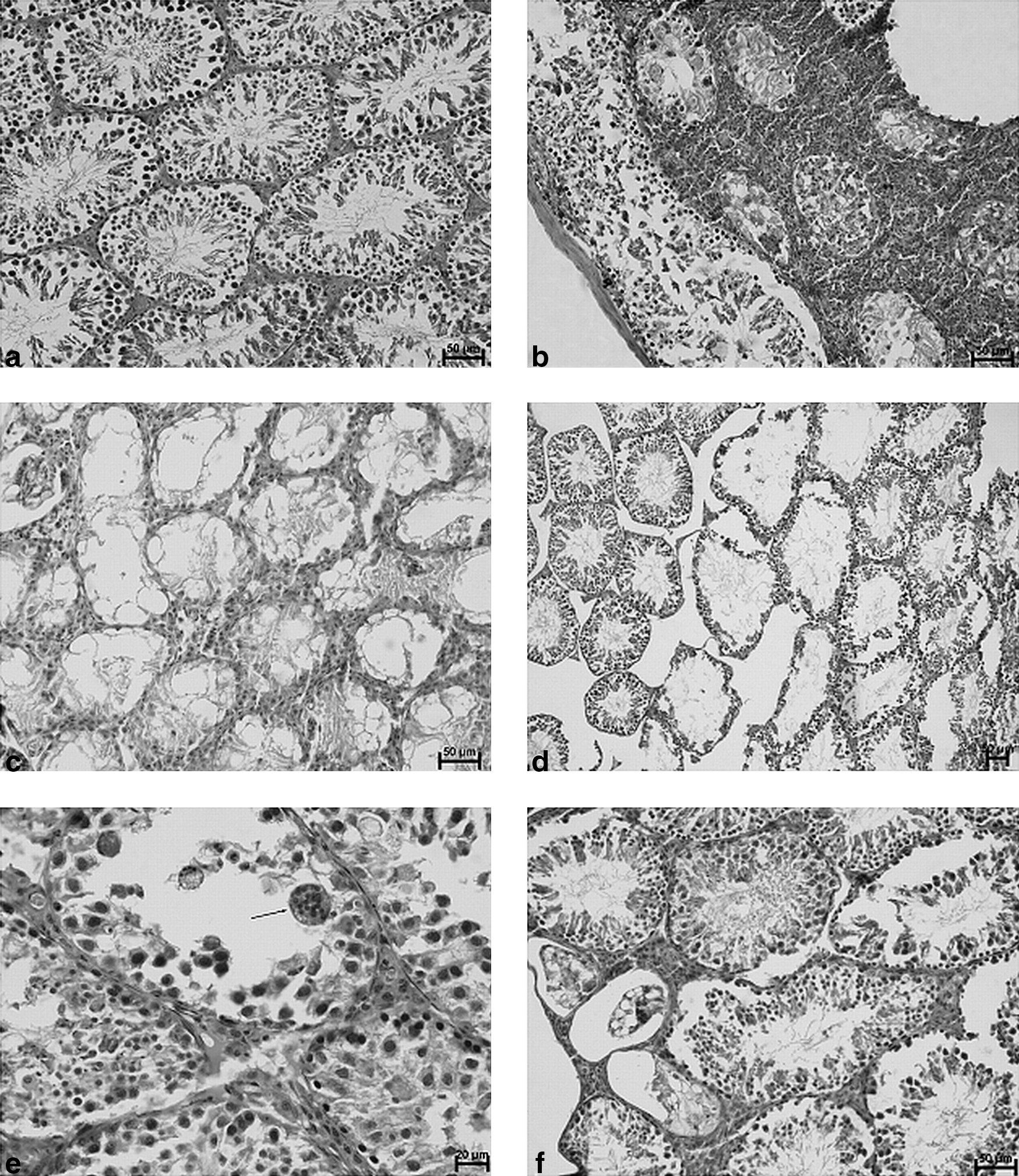
Histological picture of murine testes in EAO animals treated with IFN-τ or estradiol.
However, all immunized groups manifested various degrees of spermatogenesis impairment (Fig. 1b–f).
In the testes of animals from the IFN-treated group, focal interstitial infiltrations composed of neutrophils were observed. Some of the granulocytes invaded the seminiferous tubules. In the regions of inflammatory infiltration, the seminiferous tubules exhibited degenerative changes, associated with epithelial destruction. Accumulation of cellular debris in the tubule lumen, vacuolation of epithelium, and lack of spermatogenesis were noted. Apart from the area of inflammation, seminiferous tubules maintained a normal appearance (Fig. 1b). In the testes of animals from the estradiol-treated group, areas of interstitial infiltration decreased in comparison to the IFN-treated group. Destruction of the tubular architecture of the testes was observed. Some of the seminiferous tubules were distorted, and the lumen of the tubules was widened. The height of the epithelium had decreased, but spermatogenesis existed. The interstitial tissue between seminiferous tubules was reduced. Wide spaces between tubules exhibited the presence of interstitial edema (Fig 1d). There were also observed many degenerated, empty seminiferous tubules, with vacuolated epithelium (Fig. 1c). In the testes of estradiol- and IFN-treated animals, inflammatory infiltrations formed by neutrophils were observed. Granulocytes were present in the interstitium and also inside the seminiferous tubules. Germinal epithelium was disorganized. Shedding of degenerated germinal cells to the lumen was noted. In the lumen of seminiferous tubules, multinucleated giant cells were observed (Fig. 1e). Most of the seminiferous tubules in the testes of immunized animals, not treated with IFN-τ or E2, revealed normal structure of the germinal epithelium. A few degenerated tubules with vacuolated epithelium were observed (Fig. 1f).
E2 and IFN-τ regulate immune phenomena
Oral administration of estrogens increased the level of circulated estradiol in both DC-treated and TGC-treated animals when compared with control males
Significant rise of estradiol level was observed in estradiol-treated animals (Fig. 2). Co-administration of IFN-τ resulted in significantly higher estradiol concentration in TGC-treated animals. This tendency was also observed in DC-treated animals.

Serum estrogen (E2) level in DC- and TGC-treated mice. Oral administration of estrogen (5 μg/mouse) for 6 weeks (5 days per week) caused significant rise of the level of circulated E2. Concomitant IFN-τ treatment sustained increased concentration of the hormone. Nonparametric ANOVA with post hoc Wilcoxon test (Holm P value adjustment): *P<0.05, **P<0.01; n=9 mice per group. E2, 17β-estradiol; DC, dendritic cell; TGC, testicular germ cell; ANOVA, analysis of variance.
IFN-τ and E2 change spleen cellularity and frequency of APCs
In both immunized and nonimmunized animals, the overall weight of spleens did not significantly change (data not shown). However, fluctuations in spleen cellularity were observed (Fig. 3) in both DC-alone and TGC-treated mice.
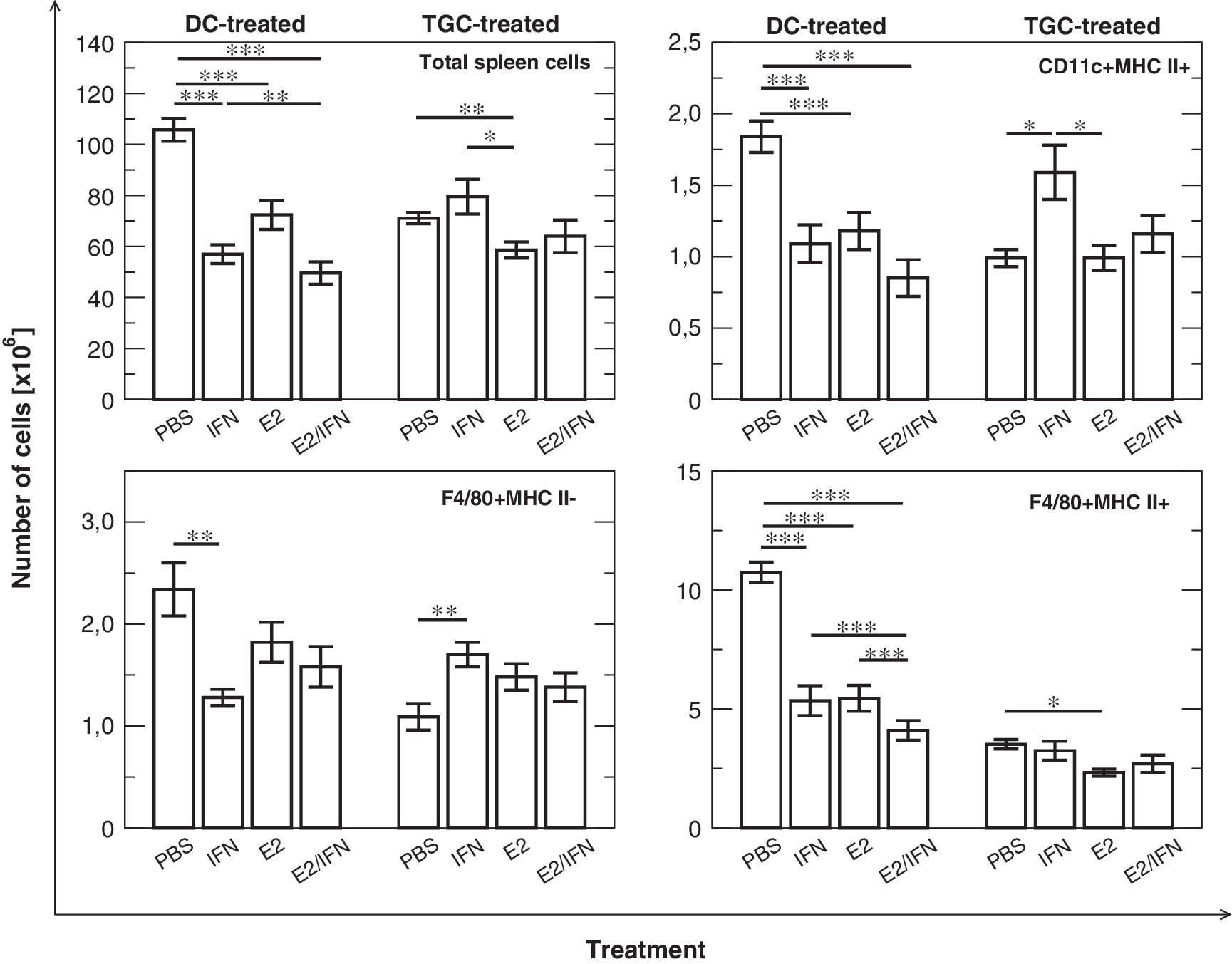
Changes in spleen cellularity and the total number of splenic antigen-presenting cells in mice treated with E2 and IFN-τ. In nonimmunized DC-treated animals, the administration of E2 or IFN-τ tends to lead to a decrease in cell number, whereas after immunization we observe mainly the effect of IFN-τ, which augments the number of cells. E2 antagonizes the effect of IFN-τ. Only in the case of F4/80+MHC II+ cells does E2 lead to a decrease in cell number after immunization. PBS: animals fed with PBS; IFN: animals fed with IFN-τ; E2: animals fed with E2; E2/IFN: animals fed with estradiol and IFN-τ simultaneously. One-way ANOVA with Tukey honestly significant difference (HSD) post hoc test: *P<0.05, **P<0.01, ***P<0.001; n=10 mice per group. PBS, phosphate-buffered saline.
DC-treated mice
E2 or IFN-τ administration decreased the total cell number (P<0.001) in DC-treated mice. These effects were synergistic, because co-administration of E2 and IFN-τ led to a further decrease in the total spleen cell number (statistically significant interaction, P=0.008).
As our hypothesis is concentrated on APCs, we analyzed the total number of splenic DCs and Mφ. The number of APCs in lymphoid tissue is low, but these cells play a crucial role in activation and regulation of the immune response (Banchereau and others 2000). Flow cytometric analysis revealed that changes were detectable in the case of the total number of mature splenic DCs (CD11c+MHC II+) and Mφ (F4/80+MHC II+) in DC-treated animals. The mice fed with E2 or IFN-τ exhibited lower numbers of CD11c+MHC II+ cells (P<0.001 in both cases). Co-administration of both mediators also led to a decrease in the number of CD11c+MHC II+ cells (P<0.001), but the effect of the interaction was not significant (P=0.1). The total number of mature F4/80 Mφ was also lower after E2 or IFN-τ administration (P<0.001) and simultaneous feeding with IFN-τ and E2 led to a stronger effect (significant interaction, P<0.001). In the case of F4/80+MHC II−, we observed a diminished cell number in IFN-τ–treated mice (P=0.003). Administration of E2 did not change the number of immature Mφ, but abolished the effect of IFN-τ (significant interaction, P=0.045).
TGC-treated mice
In the case of TGC-treated animals, total spleen cell number decreased only after E2 treatment, with or without IFN-τ (P=0.009). IFN-τ did not influence spleen cellularity and did not change the effect of E2. In TGC-treated animals, IFN-τ administration changed the number of total splenic CD11c+MHC II+ cells, and in contrast to DC-treated mice, an increase in the number of these cells was visible, both in comparison to mice fed with PBS only (P=0.014) and with E2 only (P=0.01). E2 seemed to abolish the effect of IFN-τ, when both substances were co-administered. A similar effect was noticeable in the case of F4/80+MHC II− cells: IFN-τ treatment led to augmentation of the number of these cells (P=0.01), whereas E2 alone did not exert any effect, but it neutralized the influence of IFN-τ (significant interaction, P=0.009). On the other hand, the number of mature Mφ (F4/80+MHC II+) in the spleen was regulated by E2 alone (P=0.033) and IFN-τ did not change this effect.
DTH reaction in response to TGC is regulated by IFN-τ and E2
The DTH reaction and analysis of the concentration of anti–TGC-specific antibodies was performed to assess the cellular and humoral response. DTH reaction intensity (Fig. 4) was elevated in immunized animals in comparison with mice treated with DC only (P<0.001). In TGC-treated animals, the intensity of the DTH reaction was further increased after administration of E2 (P=0.02) and IFN-τ (P<0.001), but co-administration of E2 and IFN-τ did not change it in comparison to TGC-only-treated animals. Thus, E2 and IFN-τ neutralize the effects of each other on the DTH reaction.

Delayed-type hypersensitivity reaction. Both E2 and IFN-τ increase the footpad swelling in immunized mice. E2 and IFN-τ antagonize the effect on each other, but do not reduce the intensity of DTH to the level observed in nonimmunized mice. DTH in the control group is significantly lower in comparison with all other groups. ANOVA with Tukey HSD post hoc test: **P<0.05, ***P<0.001; n=10 mice per group. DTH, delayed-type hypersensitivity.
INF-τ and E2 differentially influence IgG1 and IgG2a production in immunized animals
In the case of the humoral response, the changes in IgG1, IgG2a, IgG2b+IgG3, and IgA+IgM were investigated and analyzed with 1-way ANOVA (Fig. 5). Immunization of mice with TGC led to an increase in the concentration of TGC-specific IgG1 (P=0.006) and IgG2b+IgG3 (P=0.01), whereas the levels of other studied classes were unchanged.
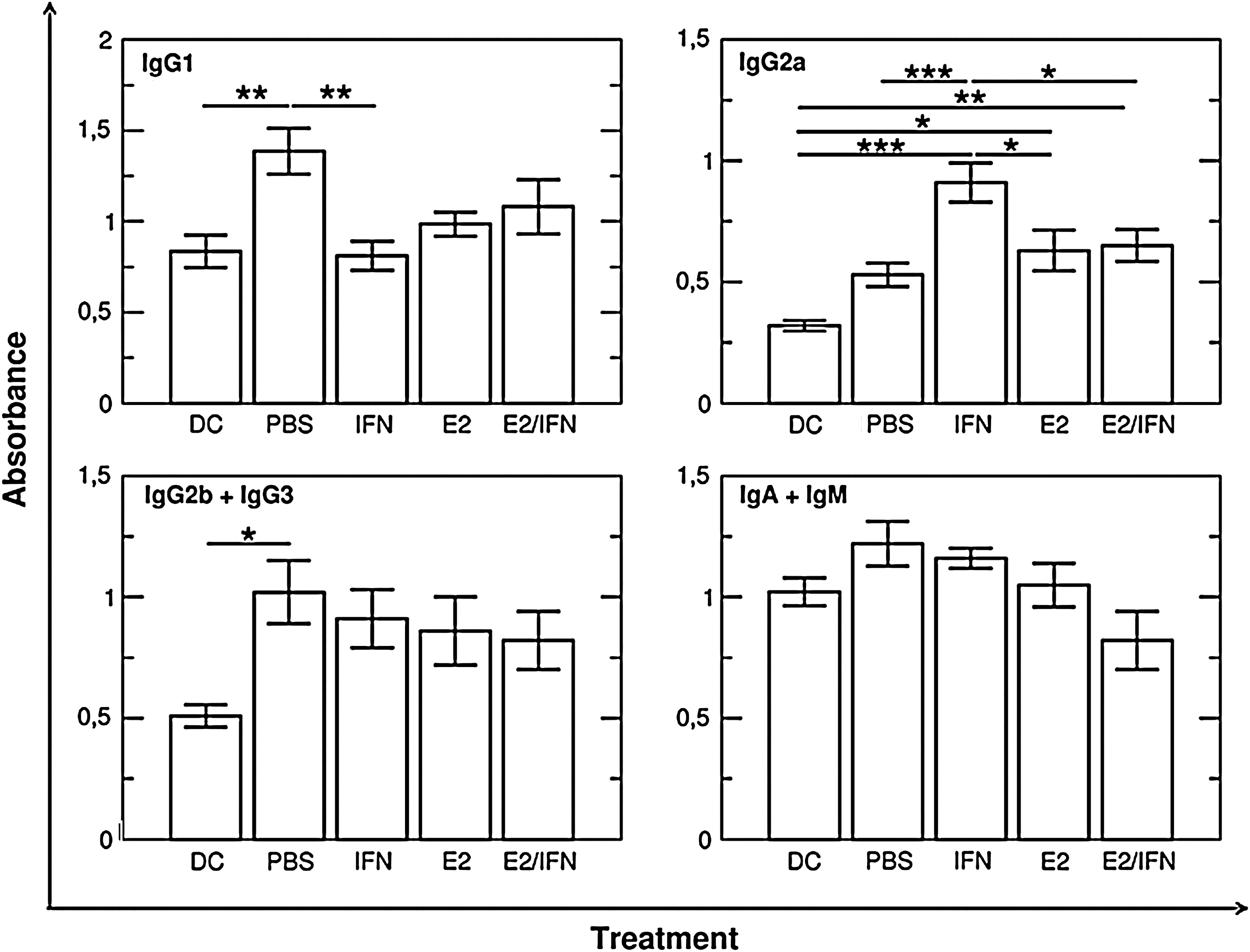
Concentration of different classes of anti–TGC-specific antibodies in serum of TGC-immunized mice. Immunization with TGC leads to significant upregulation of IgG1 and IgG2b+IgG3 production and does not influence the level of IgG2a and IgA+IgM. The administration of IFN-τ and estradiol intensifies the effect of immunization in the case of IgG2a, but does not change IgG2b+IgG3 and IgA+IgM production. Only IFN-τ administration decreases the IgG1 level. E2 co-administration abolishes the effects of IFN-τ in the case of both IgG1 and IgG2a. DC group: control (nonimmunized) animals treated with DC only; other groups were treated with TGC and fed as described on the axis. ANOVA with post hoc Tukey HSD test: *P<0.05, **P<0.01, ***P<0.001. TGC, testicular germ cell; Ig, immunoglobulin.
IgG1 concentration was lower after the administration of IFN-τ (P=0.004). E2 had no effect in this case, but it abolished the influence of IFN-τ. However, the effect of immunization was enhanced in the case of IgG2a. TGC immunization did not significantly increase the level of anti-TGC IgG2a in comparison with nonimmunized animals, but the administration of E2 (P=0.013) or IFN-τ (P<0.001) led to significantly higher production of anti-TGC IgG2a. E2 seemed to partially neutralize the effect of IFN-τ, but the production of IgG2a was still significantly higher in the E2-treated group than after TGC immunization without E2 or IFN-τ administration.
Neither E2 nor IFN-τ significantly altered the concentration of IgG2b+IgG3 and IgA+IgM.
ER-α level in splenic APCs is regulated by E2 and IFN-τ
The results of the DTH reaction, changes in spleen cellularity, and concentration of antibodies point to the possibility that E2 and IFN-τ can influence ERα level in splenic APCs. Our hypothesis is based on the regulation of ER-α by E2 and type I IFN; therefore, we conducted a semiquantitative flow cytometric analysis of ER-α content in splenic DCs and Mφ (Fig. 6).

Changes of ER-α level in splenic Mφ and DCs after E2 or IFN-τ administration in the model of EAO. In DC-treated animals, only Mφ are susceptible to E2 and IFN-τ action, and only in F4/80+MHC II−Mφ, we can observe interaction between these mediators. In TGC-treated animals, co-administration of E2 and IFN-τ reveals antagonization of IFN-τ effects. ANOVA and Tukey HSD test: *P<0.05, **P<0.01, ***P<0.001; n=10 mice per group. ER-α, estrogen receptor alpha; Mφ, macrophages.
In DC-treated animals fed with E2, IFN-τ, or both, the ER-α MFI in CD11c+MHC II+ cells did not change. However, changes were visible in immature and mature F4/80+Mφ. Only the simultaneous treatment of control animals with E2 and IFN-τ led to a decrease of the level of ER-α in MHC II− cells to nondetectable levels (pairwise Wilcoxon test, P=0.009), whereas treatment with E2 or IFN-τ did not influence the level of ER-α. In F4/80+MHC II+ cells, the administration of IFN-τ led to a decrease in ER-α level and other treatments did not change the level of ER-α in mature splenic Mφ.
A different situation was observed in TGC-treated animals. In CD11c+MHC II+ cells, the administration of IFN-τ or E2 did not lead to significant effects; however, in groups fed with E2, DCs exhibited a lower ER-α level in comparison with the group fed with IFN-τ. F4/80+MHC II− cells showed a higher level of ER-α–specific immunofluorescence (P=0.026) after feeding with IFN-τ, and E2 abolished this effect (P<0.001).
ER-α level in BM-DCs and BM-Mφ is not changed by IFN-τ and E2 treatment.
As described, orally administered E2 and IFN-τ were observed to regulate the level of ER-α; thus, we tried to answer whether this effect resulted from the direct effect on APCs. In vitro experiments were conducted on BM-DCs and BM-Mφ of C57BL6/J mice.
The cells were exposed to low (30 pg/mL) or high (200 pg/mL) concentrations of E2 and additionally stimulated with IFN-τ (n=5 per treatment). Data were analyzed with the Friedman test.
In both BM-DCs and BM-Mφ, IFN-τ did not influence the ER-α level in MHC II− and MHC II+ cells (Fig. 7). Additionally, the level of ER-α mRNA was measured in BM-Mφ, but this parameter also did not significantly change (data not shown).

ER-α content in bone-marrow–derived antigen-presenting cells. The cells were cultured in the presence of E2 at 30 and 200 pg/mL concentration and additionally treated with IFN-τ (1×105 antiviral activity unit [AVU]/mL). The differences were not significant.
Estrogen does not influence type I IFN effect
As IFN-τ did not seem to regulate the ER-α level directly, we tried to assess the influence of E2 on the effects of IFN-τ pathways. To obtain a highly pure macrophage (<90%) culture, we used bone marrow Mφ (BM-Mφ). On day 10, cultures of cells were exposed to IFN-τ alone or IFN-τ and E2 (30 pg/mL). The level of expression of 84 type I IFN-dependent genes was investigated with RT-PCR arrays in 3 independent experiments.
The scatter plot analysis showed that in comparison with Mφ treated with E2, only Mφ stimulated with both E2 and IFN-τ upregulated 8 of the 84 analyzed genes: H2-M10.2 (5.52-fold), H2-M10.4 (5.28-fold), H2-M10.6 (4.15-fold), Hoxb2 (5.48-fold), Ifna4 (5.62-fold), Ifnb1 (6.63-fold), Irf3 (4.2-fold), and Oas1a (8.5-fold). However, the changes were insignificant.
Discussion
In this article, we present data describing the influence of orally administered E2 and IFN-τ on immune parameters in mice. Our hypothesis concerned the convergent activity of estrogens and type I IFNs in EAO.
It seems that, in the case of autoimmune orchitis, at the level of target tissue in contrast to EAE but similarly to SLE, E2 and IFN-τ have a detrimental rather than a beneficial effect. We found that exogenous 17β estradiol administration at the average dose of 200 μg/kg body weight worsened impairment of spermatogenesis in adult males caused by autoimmunization. This observation is in agreement with an experiment in mice in which intramuscular administration of 17β estradiol benzoate at a dose higher than 10 μg/kg body weight caused adverse morphological changes in testes (Toyama and others 2001). On the other hand, in rats fed diets containing different concentrations of 17β estradiol, pathological changes in the seminiferous tubules developed when the concentration of blood estrogens increased over 10× above normal values, that is, from 20.3±1.8 pg/mL in the control to 257.2±32.1 pg/mL in the experimental group of rats (Cook and others 1998). Similarly, in our experiment, oral administration of E2 caused 10× increase of serum level of estradiol in TGC-treated mice (55.18±8.32 pg/mL) when compared with the control group (5.46±1.03 pg/mL).
Moreover, administration of IFN-τ caused massive infiltration of neutrophils into the testis, indicating that type I IFNs may be involved in the inflammatory process in this tissue. A similar effect was exerted by exogenously administered mouse r-IL-10, which augmented EAO in mice (Kaneko and others 2003). It is worth noting that both cytokines used the same type of cytokine receptors. In conclusion, it seems that IFN-τ and estrogens in the EAO exerted convergent adverse effects on the histological picture of the disease.
However, the course of the immune response was differentially influenced by the combined effect of IFN-τ and estrogen in contrast to each of the treatments administered alone.
We observed that the DTH was increased in the case of both E2 and IFN-τ administration, but surprisingly, combined delivery led to no effects. It seems that E2 can abolish the effect of IFN-τ and vice versa. Similar antagonistic activity was observed in the case of anti-TGC IgG1 antibodies; however, E2 alone did not change the level of this isotype. In turn, in TGC-immunized animals in comparison to nonimmunized mice, the IgG2a concentration was higher only after the administration of E2 or IFN-τ; however, we could observe a significant antagonistic influence of E2 in relation to IFN-τ. All these findings point to the participation of exogenous E2 and IFN-τ in the regulation of high-level immune phenomena during EAO. Moreover, we observed an interaction between E2 and IFN-τ, which is demonstrated by the elimination of IFN-τ activity by E2. In this context, the influence of the studied substances on the autoimmune response supports our hypothesis that type I IFNs and E2 can interact during an autoimmune disorder, but we cannot speculate whether this phenomenon is general or limited to selected autoimmune disorders.
Further analysis of the humoral response may reveal the character of Th1/Th2 balance. It is known that IFN-γ stimulates the production of IgG2a and suppresses the production of IgG1 (Snapper and Paul 1987; Stevens and others 1998), whereas interleukin-4 (IL-4) is involved in IgG1 class switching and suppression of IgG2a (Bossie and others 1987; Snapper and Paul 1987; Stevens and others 1998). IgG2a expression can also be upregulated by Th1-related cytokine IL-27 (Yoshimoto and others 2004). In our experiments, IFN-τ administration lowered TGC-specific IgG1 concentration and increased IgG2a production; thus, we can suspect that IFN-τ promotes a Th1-type response in EAO induced with DC-aided immunization with TGC. This observation differs from the results observed in other autoimmune disorders: orally administered IFN-τ seems to support the Th2-type immune response in the model of EAE (Soos and others 1997, 2002) and diabetes (Sobel and others 2008), mainly by stimulation of IL-10 production. Domination of Th1-related cytokines after immunization with TGCs alone has been reported by others (Itoh and others 1998; Tokunaga and others 2008). However, in the murine EAO model, Th2-related cytokines, such as IL-10, can also take part in pathogenesis (Kaneko and others 2003; Tokunaga and others 2008). The expression of IL-10 mRNA was also observed in testicular DCs in inflamed but not healthy testes in rats (Rival and others 2007) and in T cell receptor (TCR)-γδ+ and TCR-αβ+ T cells in murine EAO induced by injecting Listeria monocytogenes into the testes (Matsuzaki and others 1997; Mukasa and others 1998). Thus, if stimulation of IL-10 production after oral administration of IFN-τ is a common phenomenon, our results seem to be convergent with the effects of this cytokine in EAE and diabetes.
In contrast to IFN-τ, E2 does not influence antibody production significantly, but abolishes the effect of IFN-τ. Changes in the production of different classes of antibodies are consistent with the results of DTH, because this reaction is also connected with a Th1 response. Estrogen orally administered can exert proinflammatory properties in CIA (Hosoda and others 2004) or increase inflammatory markers in women treated with oral estrogen therapy (Hemelaar and others 2006; Shifren and others 2008). Thus, we can speculate that E2 can regulate the cytokine network in such a way that, on the one hand, we can observe an intensification of inflammatory Th1-related DTH reaction and, on the other hand, observe that E2 reduces the effect of IFN-τ. The background of mixed E2 activity requires further research.
The changes in DTH and the level of TGC-specific immunoglobulins are accompanied by fluctuations in the number of splenocytes. The weight of the spleen was unaffected after the administration of E2 and this is consistent with other reports (Dhaher and others 2001; Li and McMurray 2006). The total number of spleen cells was mainly changed in nonimmunized DC-treated mice. Both E2 and IFN-τ significantly lowered average spleen cellularity. In TGC-treated mice, such treatment had a minimal effect on this parameter. The effect of IFN-τ is similar to the effect of IFN-α, because IFN-α reduces the number of leukocytes in both the spleen and the bone marrow of mice (Rosenthal and others 1989) and inhibits the generation of leukocytes in humans (Facchetti and others 1997).
In immunized mice, IFN-τ administration leads to an increase in the number of CD11c+MHC II+and F4/80+MHC II− cells and E2 abolishes these effects. On the other hand, E2 lowers the number of F4/80+MHC II+ cells and IFN antagonizes the activity of E2. It seems that the changes in DTH reaction and antibody production in the case of IFN-τ can be partially attributed to the increase in the number of APCs, and an antagonistic effect of E2 is visible here as well. Further, the changes in the number of different populations of APCs may be a reflection of rebuilding the APC compartment in peripheral immune organs, which can influence the nature of the immune response.
The second part of our hypothesis concerns the influence of IFN and E2 at the level of ER-α in APCs. Similar to the number of splenic APCs, the effect of E2 and IFN-τ is different in DC-treated and TGC-treated animals, and in some cases, it is completely opposite. Only IFN-τ enhances the expression of ER-α and E2 abolishes this effect in F4/80-MHC II+ cells, whereas in F4/80+MHC II+ cells and CD11c+MHC II+ cells, IFN-τ administration does not increase the ER-α level; however, E2 decreases ER-α expression in comparison with mice treated with IFN-τ. This observation supports our assumption that type I IFN administration can regulate the level of ER-α, but the effect is dependent on the target cell population. The differences between immunized and nonimmunized animals point to the possibility that mediators released during inflammation can modify the influence of IFN-τ. Our experimental design does not allow us to assess the connection between ER-α level and its influence on immune mechanisms. Regression analysis (data not shown) does not indicate a direct influence of ER-α level on the overall immune response parameters. We can only speculate that changes in ER-α level can simulate the higher or lower concentration of estrogen, because the level of estrogen–receptor complex is dependent on the concentration of both estrogen and the receptor. It is worth of note that in TGC-treated and estrogen-administered animals, in which serum estrogen concentration was significantly higher than in estrogen-untreated groups, the level of ERα in APCs was significantly lower when compared with the IFN-τ–treated groups.
The estrogen–receptor complex can further interact with other signaling pathways, such as the pro-inflammatory transcription factor nuclear factor-kappa B and mitogen-activated protein kinase pathways (Suzuki and others 2007b). However, to define the association between ER-α content and function of APCs, more detailed studies are needed.
In the last in vitro experiments, we tried to assess whether E2 or IFN-τ can directly regulate the ER-α level in BM-DCs and BM-Mφ and, on the other hand, whether E2 can influence expression of type I IFN-dependent genes. The lack of significant direct effects in the in vitro model leads us to the conclusion that the observed in vivo phenomena of a modulatory influence of IFN-τ and E2 and their common interactions may be attributed to inflammatory milieu or the indirect activity of both these immunomodulators on other cellular and molecular targets. Collectively, the results in this article strongly support the hypothesis that IFN-τ and E2 can interact in the context of the autoimmune phenomena. This interaction is visible at the level of overall markers of immune response (specific cellular and humoral response), in spleen APC compartment composition, and at the level of intracellular phenomena in APCs (ERα receptor protein level). This interaction can be potentially exploited in therapy, as co-administration of IFN and E2 can lead to different effects in comparison to the activity of each substance alone.
Footnotes
Acknowledgments
This study was supported by a grant from the Polish Ministry of Sciences and Higher Education (No. N30800931/3409). The authors are very grateful to Prof. Fuller Bazer and Dr. Joann Fleming from Texas A&M University, for providing ovine rIFN-τ, and Dr. Jacek Bania from Wrocław University of Environmental and Life Sciences, for providing the cell line production of GM-CSF.
Author Disclosure Statement
No competing financial interests exist for any of the authors of this manuscript.