
Abstract
Natural killer (NK) cells are important in the ocular surface innate response against viral and bacterial infection. Major-histocompatibility-complex class I-related chain A (MICA) antigens are ligands of natural killer group 2D, an activating or coactivating receptor expressed on NK cells. Recent studies demonstrated that interferon-gamma (IFN-γ) could modulate MICA expression in tumor cells. However, little is known about MICA expression and regulation in human corneal epithelium. Our study assessed whether the proinflammatory cytokine, IFN-γ, affects MICA expression in human corneal epithelium. We identified low levels of surface MICA expression in corneal epithelium using flow cytometry. IFN-γ promoted surface MICA expression in corneal epithelium and increased soluble MICA levels in a dose-dependent manner. IFN-γ also enhanced NK cell-mediated cytotoxicity against the corneal epithelium. Anti-MICA antibodies could further block this process. In summary, we describe a novel IFN-γ function in the regulation of the innate response in ocular surfaces.
Introduction
As a ligand of NKG2D, MHC class I chain-related A molecules (MICA) share structural homology with classical human leukocyte antigen molecules, yet do not interact with β2 microglobulin, nor bind peptides, and are not expressed on normal circulating lymphocytes. In addition, MICA antigens could elicit a very powerful immune response by activating the effector immune cells (González and others 2006). MICA interaction with its receptor potentially plays a central role in innate immunity. While MICA ligand levels might also determine the strength of such immune responses, the signals and mechanisms regulating MICA expression still need to be elucidated.
Specific factors involved in the regulation of MICA expression have now been indentified, although their precise mechanism of these factors has yet to be elucidated. Cell surface expression of MICA, which is under the control of a heat shock promoter, could be induced by cellular stress (Cerwenka and Lanier 2003). Up-regulation of MICA has also been reported to occur upon deoxyribonucleic acid (DNA) damage, oxidative stress, and treatment with retinoic acid or the histone deacetylase inhibitor sodium valproate (Mistry and O'Callaghan 2007). However, relatively little is known about whether MICA expression and regulation is involved in corneal innate immunity.
Most investigations into ophthalmic applications of interferon (IFN) have dealt with viral infection, mainly HSV (Sundmacher and others 1987). However, the roles played by IFN-gamma (IFN-γ) in viral ocular infections are not fully established. Previous studies found that IFN-γ effectively induces MHC class I and II molecules' expression in retinal pigment epithelial cells and other stressed cells (Zavazava and others 1991; Propper and others 2003). Several publications have also recently focused on the effect of IFN-γ on MICA expression in tumor cells. Schwinn and others (2009) suggested that the down-regulation of MICA expression by IFN-γ may facilitate the escape of MHC class I-negative melanoma cells from NKG2D-mediated killing by NK cells. Similar results were obtained by Zhang and others (2008), who reported that IFN-γ decreased surface MICA protein expression in human cervical carcinoma and erythroleukemia K562 cell lines. Thus, the correlation between IFN-γ and MICA expression in human corneal epithelium also needs to be further investigated.
Our study describes the detection of MICA expression at low levels on the surface of human cultured corneal epithelium. Treatment of corneal epithelial cells with IFN-γ could have up-regulated the surface and soluble expression of MICA in a dose-dependent manner, which enhanced an effective MICA-dependent killing of the corneal epithelium by allogeniec NK cells. Thus, we speculated that IFN-γ might contribute to the innate responses of the ocular surface by up-regulating MICA expression in corneal epithelium and activating NK cells.
Materials and Methods
Antibodies
Primary
Anti-human CD56 and CD3 microbeads (Miltenyl Biotech), anti-human MICA monoclonal antibodies (R&D Systems), isotype control mouse IgG (BD Biosciences), and mouse anti-human cytokeratin 3/12 (Santa Cruz Biotechnology) were purchased.
Secondary
Affinity-purified secondary antibodies and species-absorbed conjugates for dual-labeling were purchased from Sigma-Aldrich.
Cell culture
The Ethics Committee of Shanghai Eye, Ear, Nose and Throat Hospital approved this study. Corneal epithelium was obtained from the eyes of adult patients provided by Shanghai Eye Bank and cultured as described previously (Djalilian and others 2008). Briefly, corneoscleral rims from the donor corneas were obtained as soon as the central corneal button had been used for the penetrating keratoplasty. The tissue was rinsed with sodium chloride for 3 times and then treated with 1.3 unit/mL dispase II in defined keratinocyto serum-free medium (K-SFM) at 4°C for 16 h. The corneal epithelial sheets were peeled off and digested with 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA) at 37°C for 10 min to be rendered into single cells, which were then seeded on the 3T3 fibroblast feeder layers. To prepare growth-arrested feeder layers, subconfluent 3T3 cells were incubated with 10 μg/mL mitomycin C (Sigma-Aldrich) for 2 h at 37°C and then trypsinized. The mitomycin C–treated cells were subcultured in Dulbecco's modified Eagle's medium (DMEM; Hyclone, Thermo, Fisher Scientific, Inc.), supplemented with 10% fetal bovine serum at a density of 2×104 cells/cm2. Human corneal epithelial cells used in the whole study were obtained from 5 corneoscleral rims and cultured to passage 1 for all experiments. Corneal epithelial cells were maintained in the medium comprising 75% K-SFM and 25% DMEM. For each experiment except the enzyme-linked immunosorbent assay (ELISA), we first incubated cultured dish in 0.05% trypsin-EDTA, then gently shook and aspirated off 3T3 feeder cells, and washed with phosphate-buffered saline (PBS) for 3 times to remove feeder cells. In some experiments, cells were treated for 48 h with 100 U/mL or 1,000 U/mL IFN-γ (Sigma-Aldrich).
Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized venous blood obtained from normal healthy volunteer donors by Ficoll-Hypaque density gradient centrifugation (Amersham Pharmacia Biotech). NK cells were isolated from PBMC by magnetic cell sorting (Miltenyi Biotec).
Reverse transcription–polymerase chain reaction
Total ribonucleic acid (RNA) was extracted from cells using TRIzol reagent (Invitrogen) according to the manufacturer's protocol, and reverse transcription–polymerase chain reaction (RT-PCR) was conducted as previously described (Kim and others 2007). Complimentary DNA was PCR amplified with the following specific primers: MICA, forward 5′-ATCTTTGAGCCACGACAC-3′ and reverse 5′-CTTCTTACAACAACGGACA TA-3′. Glyceraldehyde 3-phosphate dehydrogenase was used as a control.
Flow cytometry
MICA expression in corneal epithelium was detected using flow cytometry. Anti-human MICA antibodies at the appropriate concentrations were added to 1×105 cells for 30 min at 4°C, washed twice with ice-cold PBS, and stained with secondary antibodies. Samples were analyzed with a flow cytometer (Beckman Coulter, Inc.).
Enzyme-linked immunosorbent assay
Soluble MICA (sMICA) was detected in cell culture supernatants by a human MICA ELISA kit (ProSpec-Tany; TechnoGene) according to the manufacturer's protocol.
Cytotoxicity assay
The cytolytic activity of NK cells against human corneal epithelium was assessed using an lactate dehydrogenase cytotoxicity detection kit (Roche Biochemicals) according to the manufacturer's instructions. Corneal epithelial cells treated with or without IFN-γ for 48 h were centrifuged and washed 3 times with PBS, harvested by scraping, and then mixed with NK cells in triplicate wells of 96-well U-bottom plates for 4 h at 37°C. Effector (E) to target (T) ratios (E:T) were 3:1 and 1:1. Plates were subsequently centrifuged at 250 g for 10 min, and 100 μL of the cell-free supernatants was collected and incubated with the kit reaction mixture. For blocking studies, target cells were preincubated for 30 min at 4°C with 10 μg/mL of anti-MICA antibody. Assays were stopped after incubation for 4 h at 37°C. Absorbances at 490 nm were recorded and the percent cytotoxicity for each effector/target cell ratio was calculated using the following formula: cytotoxicity=(experimental−effector spontaneous−target spontaneous)/(target maximum−target spontaneous) ×100%.
Statistical analysis
All data are representative of experiments performed at least 3 times. Differences in surface and soluble MICA at different IFN-γ concentrations were assessed by analysis of variance test. The Bonferroni method was also used to correct for multiple testing. A 95% confidence interval (P<0.05) was considered to be significant.
Results
Low expression of surface MICA in cultured human corneal epithelial cells
Flow cytometric analyses and ELISA were used to determine whether primary cultures of human corneal epithelium cells express MICA. Corneal epithelial cells were obtained via mechanical and enzymatic digestion (trypsin) and cultured to enrich the different cell populations. As MICA detection has been demonstrated to be trypsin-sensitive (Saikali and others 2007), adult human cells were cultured until the first passage to determine their physiological characteristics. At this time, cells had reached desirable numbers and were also expressing cytokeratin12, which is the biomarker for corneal epithelium (Fig. 1A, 1B). Surface MICA expression in corneal epithelium was identified by flow cytometry (Fig. 1C), whereas sMICA was not detected using ELISA. MICA mRNA levels were also detected by RT-PCR analysis (Fig. 1D). This further prompted us to consider whether IFN-γ has an effect on corneal epithelium.
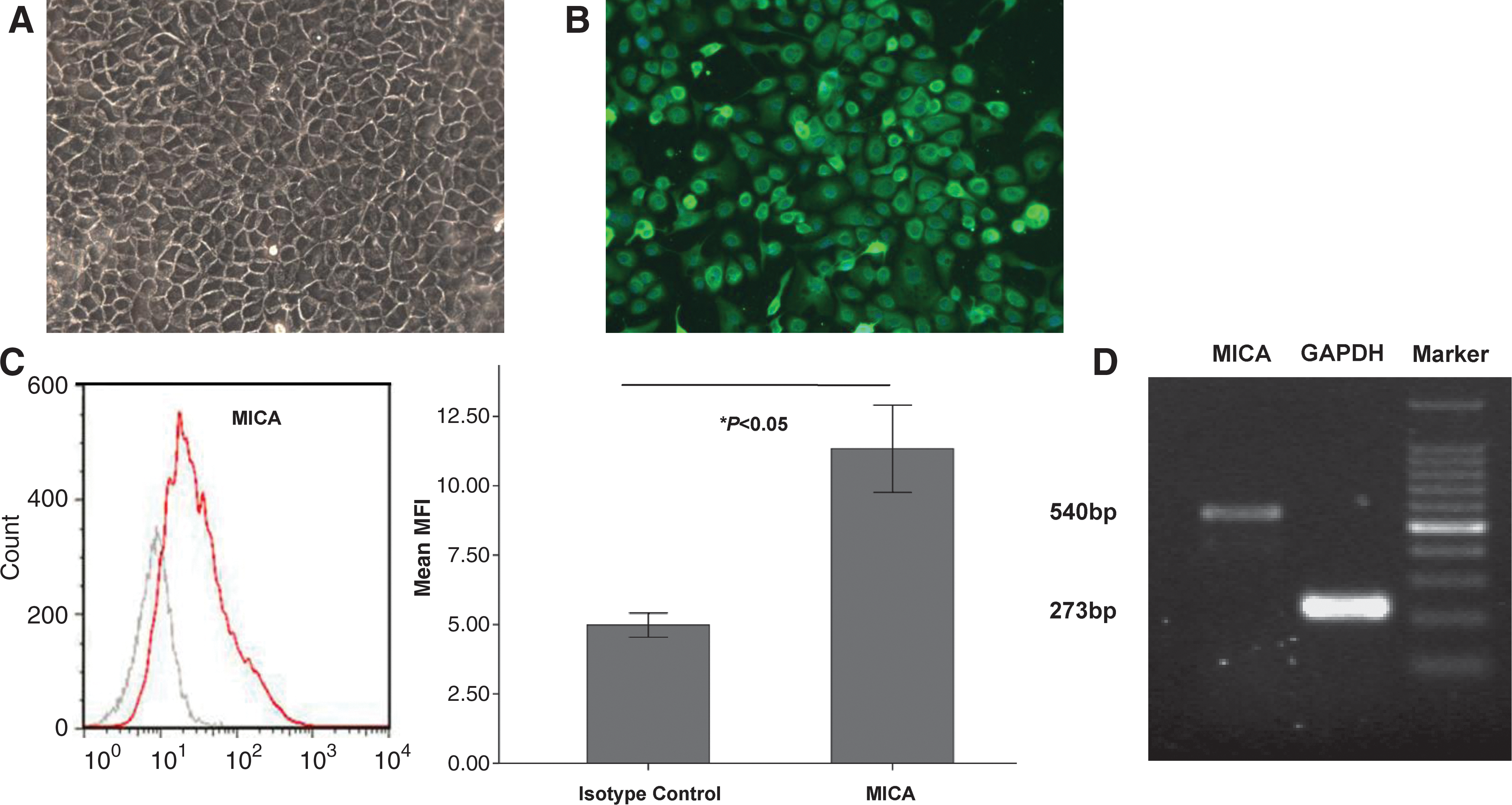
Expression of major-histocompatibility-complex class I-related chain A (MICA) on human corneal epithelium in vitro.
Up-regulation of MICA by IFN-γ stimulation in corneal epithelial cells
To determine the influence of IFN-γ on MICA expression, corneal epithelial cells were incubated with the cytokine for 48 h, before the MICA expression pattern was analyzed. Flow cytometric analysis revealed that MICA expression was higher with the sequential IFN-γ exposure than without IFN-γ treatment (Fig. 2A). Moreover, up-regulation of surface MICA expression was dependent on the concentration of IFN-γ. A higher level of MICA expression was induced using 1,000 U/mL than that observed using 100 U/mL (Fig. 2A). MICA mRNA levels were also increased in a dose-dependent manner by IFN-γ stimulation, and were more significantly enhanced using 1,000 U/mL treatment (Fig. 2B).
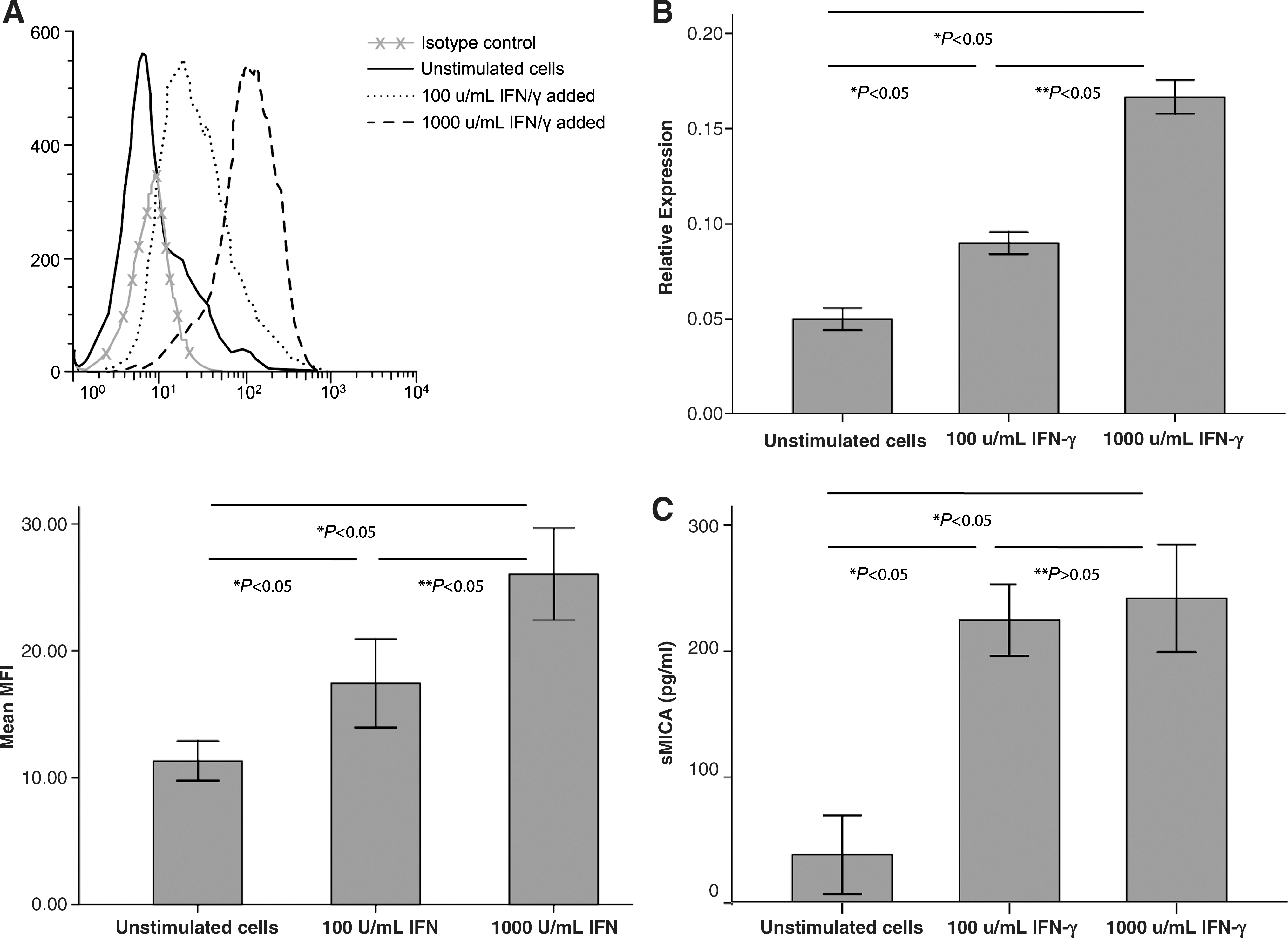
Effect of interferon-γ (IFN-γ) on the expression of MICA in cultured human corneal epithelium.
To further confirm the effect of IFN-γ on sMICA release, we detected sMICA concentration in culture supernatants of corneal epithelial cells. ELISA results indicated that sMICA released into the culture supernatants was dramatically increased after IFN-γ treatment for 48 h, whereas no significant difference was observed between the 2 concentration groups (Fig. 2C).
Collectively, these data demonstrated that IFN-γ promoted surface MICA expression in corneal epithelium in a dose-dependent manner and soluble MICA release into the supernatant by up-regulating MICA mRNA transcription.
IFN-γ-enhanced allogeneic NK cells mediated human corneal epithelium killing
By investigating the impact of IFN-γ on the lysis of corneal epithelial cells by NK cells, we found that killing was promoted by IFN-γ pretreatment at both E:T ratios (Fig. 3A). At the 3:1 ratio baseline, ∼10% of cells were lysed by NK cells. However, the number of killed cells increased to ∼15% at 100 U/mL and 22% at 1,000 U/mL of IFN-γ treatment. Meanwhile, increased NK cell-induced killing was observed in the 3:1 ratio group compared with the 1:1 ratio group, both in the isotype control and the anti-MHC I antibody group (P< 0.05) (Fig. 3B).
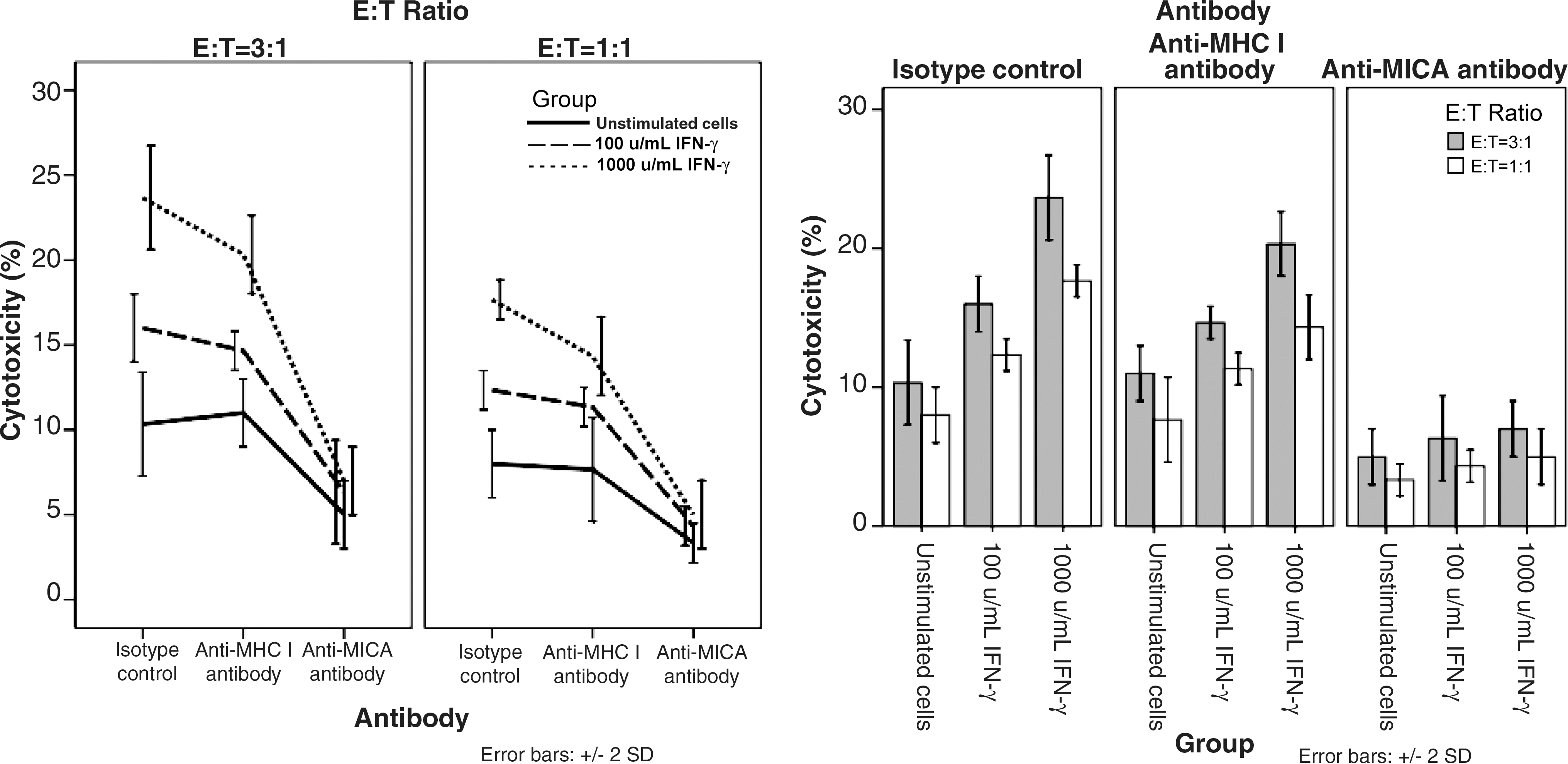
Cytotoxicity of natural killer (NK) cells to human corneal epithelium pretreated with IFN-γ. Corneal epithelial cells were treated with culture medium (unstimulated) or 100 U/mL or 1,000 U/mL IFN-γ for 48 h and used as targets cells. Target cells were washed 3 times with phosphate-buffered saline before mixed with NK cells. Assays were conducted in the presence of isotype control or anti-MICA antibody or anti-major histocompatibility complex (MHC) I antibody.
To ascertain whether NK cell recognition of the corneal epithelial cells was indeed mediated by MICA, and that the enhanced killing after IFN-γ treatment required the interaction between NK cells and MICA, antibody-blocking experiments were carried out. In the presence of the anti-MICA antibody, killing was strongly decreased in both E:T ratios. MICA blocking reduced the killing efficiency of the different groups to similar levels. This finding indicated that the enhanced killing of IFN-γ-treated cells was indeed caused by the interaction of NK cells with MICA in epithelial cells. To further support our hypothesis, we analyzed the blocking experiments with an anti-MHC class I molecule antibody, which did not affect the killing, confirming that the MICA blocking was not caused by steric hindrance.
Discussion
Previous to our study, relatively little was known about whether MICA expression and regulation are involved in corneal innate immunity. Although most investigations into ophthalmic applications of IFN have dealt with viral infection, mainly HSV, the roles of IFN-γ in viral ocular infections require further investigation.
Notably, our study revealed that MICA was weakly expressed on the surface of human corneal epithelium cells before the addition of IFN-γ, which was in contrast to both mRNA and ELISA results. This may be because MICA protein is stored in the cytoplasm of normal cells. Upon inflammation or stress, this protein is delivered to the surface of cells or secreted into intercellular matrix (González and others 2006). Previous study has shown that CMV could obstruct this process, in which MICA localization to or retrieval from the trans-Golgi network was blocked, thus abrogating surface expression (Wu and others 2003).
Conclusions drawn by previous studies about the relationship of IFN-γ and MICA expression seemed paradoxical. Schwinn and others (2009) and Zhang and others (2008) both reported that IFN-γ could decrease surface MICA protein expression in human tumor cells. However, Saikali and others (2007) found that cultured oligodendrocytes expressed detectable levels of MICA and MHC class I molecules, and had increased MICA expression after 2 days of IFN-γ plus tumor necrosis factor-α treatment. In our study, we demonstrated that IFN-γ treatment up-regulated MICA expression in cultured human corneal epithelium, which was dependent on the cytokine dose. More importantly, IFN-γ-treated cells were more susceptible to NK cell cytotoxicity. Antibody-blocking experiments revealed that the enhanced lysis of IFN-γ-treated cells was attributed to the interaction between NK cells and MICA on the surface of the corneal epithelial cells. This effect might be one of the mechanisms by which IFN-γ contributed to immune responses against viral infection in the cornea.
We also detected an increase of MICA expression at the transcriptional level after IFN-γ stimulation by RT-PCR analysis. Thus, we postulated that MICA expression up-regulation was a consequence of an increase in specific mRNA. To date, the transcriptional regulation mechanism of MICA is still poorly understood. Many transcription factors, such as specificity protein 1 and specificity protein 3, also play important roles in the regulation of MICA transcription (Venkataraman and others 2007). Other regulatory mechanisms, such as DNA methylation, may also be involved in its transcription. Further study is needed to clarify whether these regulatory mechanisms could be involved in corneal epithelium.
IFN-γ can further regulate MICA expression not only at the transcriptional level, but also at the post-translational level by promoting proteolytic cleavage. Zhang and others (2008) demonstrated that IFN-γ could up-regulate the expression of matrix metalloproteinase (MMP) and down-regulate the expression of the MMP inhibitor. Furthermore, IFN-γ stimulation increased the production of sMICA in the culture supernatants of tumor cells. Our data showed that IFN-γ was involved in the release of sMICA without reducing the cell surface expression of MICA. Thus, the results shed light on the possibility that IFN-γ modulates a different pattern of MICA expression in corneal epithelium.
IFN-γ is well recognized as an important regulator of immune responses. It has been found to regulate NK, CTL, and NKT cells that are critical to anti-virus immunity (Malmberg and others 2002; Dong and others 2005; Ota and others 2005). Tumor cells were lysed by NK cells in an MICA-dependent manner and IFN-γ strongly interfered with the efficiency of MICA-mediated cytolysis (Maccalli and others 2007). In the present study, we treated cultured corneal epithelium with IFN-γ before added to NK cells and demonstrated that IFN-γ up-regulated MICA expression in cells, thereby enhancing the cytotoxicity of NK cells. In contrast, Zhang and others (2008) and Bui and others (2006) showed that IFN-γ down-regulated MICA or H60 expression in tumor cells. Such different results might be due to the different cell lines used in each study. It is tempting to speculate that this type of IFN-γ response in corneal epithelium might be a regulatory mechanism to trigger the activation of the NKG2D receptor that is generally associated with enhanced cytotoxic activity of NK cells to virus-infected cells, thereby facilitating the immune system.
In conclusion, we propose for the first time that IFN-γ enhances MICA expression in human corneal epithelium, and consequently improves the cytoxicity to virus-infected cells. Cell surface expression of MICA should be tightly controlled in order to avoid the attack against normal corneal epithelial cells, which was crucial in maintaining immune privilege in the cornea. Moreover, MICA pathway might represent a promising mechanism for improving the immune responses against corneal infections.
Footnotes
Acknowledgments
This work was supported by grants from Scientific Research Program, Ministry of Health, China (2010–2012); National Natural Science Foundation of China (30872815; 81170817); Excellent Leading Scholars Program, Science and Technology Commission of Shanghai Municipality (10XD1401100); Outstanding Doctoral Research Program of Fudan University.
Author Disclosure Statement
No author has any conflict of interest.