
Abstract
Patients with chronic granulomatous disease (CGD) cannot produce reactive oxygen species (ROS) due to a genetic defect in the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase system. Dysregulation of the
Introduction
C
While the central role of ROS as direct antimicrobial effectors is still under discussion, ROS are also linked to key intracellular signaling events, such as the activation of transcription factors (Sen and Packer 1996; Sun and Oberley 1996), among which nuclear factor kappa-light-chain-enhancer of activated B cells (NFκβ) (Blackwell and others 1996; Rusyn and others 1998), induction of mitogenesis (Irani and others 1997; Joneson and Bar-Sagi 1998), and activation of proteases (Reeves and others 2002). ROS can also act as a substrate or cofactor for the catabolic indoleamine 2,3-dioxygenase enzyme (IDO) (Shimizu and others 1978; Hayaishi 1996; Thomas and Stocker 1999; Dang and others 2000). IDO is the rate-limiting enzyme in the tryptophan metabolism (Stone and Darlington 2002). Activation of IDO results in decreased levels of
The tryptophan metabolism has also been linked to Th17 responses. In a collagen-induced arthritis mice model,
A robust Th17 response is necessary for an adequate defense against Candida (Huang and others 2004). Th17 cells are characterized by the production of IL-17A (Park and others 2005), which is important for attracting neutrophils to the tissues (Kolls and Linden 2004). The importance of IL-17A in antifungal host defense is reflected by the fact that IL-17A receptor antagonist-deficient mice have an increased susceptibility to disseminated candidiasis (Huang and others 2004). Furthermore, in a murine oropharyngeal candidiasis model, IL-17A- or IL-23p19-deficient animals showed increased severity of oropharyngeal candidiasis (Conti and others 2009). Also humans with chronic mucocuteneous candidiasis (CMC) and hyper-IgE syndrome (HIES) have decreased Th17 responses that are believed to be responsible for their susceptibility to fungal infections (Grimbacher and others 1999; Eyerich and others 2008; Ma and others 2008; Milner and others 2008; Minegishi and others 2009; van de Veerdonk and others 2011).
CGD is the primary immunodeficiency with the highest incidence of invasive fungal infections (Antachopoulos 2010), mainly due to Aspergillus spp. (lifetime incidence 25%–40%). Candida is commonly encountered in cases of fungal meningitis, fungemia, and lymphadenitis (Winkelstein and others 2000; van den Berg and others 2009; Antachopoulos 2010). It is currently not clear why patients with CGD are specifically susceptible to fungal infections, although infections with bacterial pathogens such as Staphylococcus aureus are also often encountered. The absence of ROS has been shown not to be the direct explanation of the increased susceptibility (Henriet and others 2011).
In the current study, the tryptophan metabolism and IL-17A response in human CGD patients were investigated. We demonstrate that CGD patients, in contrast to the p47phox-deficient mice, are able to convert
Materials and Methods
Patients and healthy volunteers
Blood was collected from healthy volunteers who did not suffer from infectious or inflammatory diseases, and from 8 CGD patients, harboring homozygous mutations in the NCF1 gene (p47-phox) or CYBB gene (p91-phox). After informed consent was given, blood was collected by venipuncture into 10 mL ethylenediaminetetraacetic acid tubes (367525; BD). The clinical characteristics of all patients are presented in Table 1.
Reagents
The following study materials were used: recombinant IFN-γ (Boehringer Ingelheim BV);
Microorganisms
C. albicans ATCC MYA-3573 (UC 820) (Lehrer and Cline 1969) was grown overnight in Sabouraud broth at 37°C; cells were harvested by centrifugation, washed twice, and resuspended in culture medium. C. albicans was heat killed for 1 h at 100°C; S. aureus (ATCC 25923) was heat killed for 30 min at 100°C; heat-killed conidia and hyphae of Aspergillus fumigatus (V4507) and Aspergillus nidulans (V44-46) were prepared as described previously (Chai and others 2009; Henriet and others 2011).
Human mononuclear cells and neutrophils
Peripheral blood mononuclear cells (PBMCs) and neutrophils were isolated from peripheral blood using density gradient centrifugation (800 g) of diluted blood (1 part blood to 1 part pyrogen-free saline) over Ficoll-Paque (Pharmacia Biotech) (Netea and others 2006). PBMCs were harvested, washed twice in phosphate buffered saline (PBS), and suspended in culture medium. To remove the erythrocytes from the neutrophils, the pellet was shocked at least twice with an ice-cold lysing reagent (2.75 g NH4Cl + 0.25 g KHCO3) for 15 min. Subsequently, cells were washed in PBS and resuspended in culture medium. The PBMCs and neutrophils were counted in a hemocytometer, and their concentration was adjusted to 5×106 cells/mL.
About 5×105 PBMCs or neutrophils in a total volume of 200 μL per well were incubated at 37°C in round-bottomed, 96-well plates (Greiner) with the different stimuli, as indicated in the figure legends. After 24 h, 48 h, or 7 days of incubation, supernatants were collected and stored at −20°C until assayed. When cells were cultured for 7 days, this was done in the presence of 10% human pooled serum.
Enzyme-linked immunosorbent assay
The concentration of IFN-γ, IL-6, IL-17A, and TNF-α was measured in cell culture supernatants using enzyme-linked immunosorbent assay (ELISA) (IL-17A and TNF-α: R&D Systems; IFN-γ and IL-6: Sanquin), according to the instructions of the manufacturer.
Real-time polymerase chain reaction
One million freshly isolated PBMCs or neutrophils were incubated with the various stimuli. After 24 h of incubation at 37°C, total ribonucleic acid (RNA) was extracted in 400 μL of TRIzol reagent (Invitrogen). Isolated RNA was being reverse transcribed into complementary DNA using oligo(dT) primers and MMLV reverse transcriptase. Polymerase chain reaction (PCR) was performed using a 7300 realtime PCR system (Applied Biosystems). The primer sequences for human IDO are as follows: 5-GGT-CAT-GGA-GAT-GTC-CGT-AA-3 (forward) and 5-ACC-AAT-AGA-GAG-ACC-AGG-AAG-AA-3 (reverse). β2M was used as a reference gene, for which the primers were 5-ATG-AGT-ATG-CCT-GCC-GTG-TG-3 (forward) and 5-CCA-AAT-GCG-GCA-TCT-TCA-AAC-3 (reverse). PCR conditions were as follows: 2 min at 50°C and 10 min at 95°C, followed by 40 cycles of PCR at 95°C for 15 seconds, and 60°C for 1 min. Data are expressed as fold increase compared with the unstimulated sample.
High-performance liquid chromatography
To compare the amount of
Statistical analysis
Experiments were performed in duplicate, and supernatants were pooled. The differences between groups of 3 or more subjects were analyzed using the Mann–Whitney U test or the Wilcoxon signed rank test, for unpaired and paired data, respectively. The level of significance between groups was set at P<0.05 (*) and P<0.01 (**). Data are presented as mean±standard error of the mean.
Results
CGD patients have an increased production of proinflammatory cytokines in vitro
The cytokine profile of CGD patients upon stimulation with several stimuli was investigated. PBMCs of CGD patients produced more IL-6 and TNF-α upon stimulation with the specific TLR4 ligand LPS (Fig. 1). Similarly, heat-killed C. albicans induced more IL-6 and TNF-α production in PBMCs isolated from CGD patients (Fig. 1). In line with this, stimulation with live A. nidulans and A. fumigatus conidia, 2 commonly encountered species in CGD patients, resulted in a higher cytokine production by PBMCs of CGD patients compared with healthy controls (Fig. 1). Heat-killed Aspergillus conidia and hyphae showed to be poor stimulators of the proinflammatory cytokines.
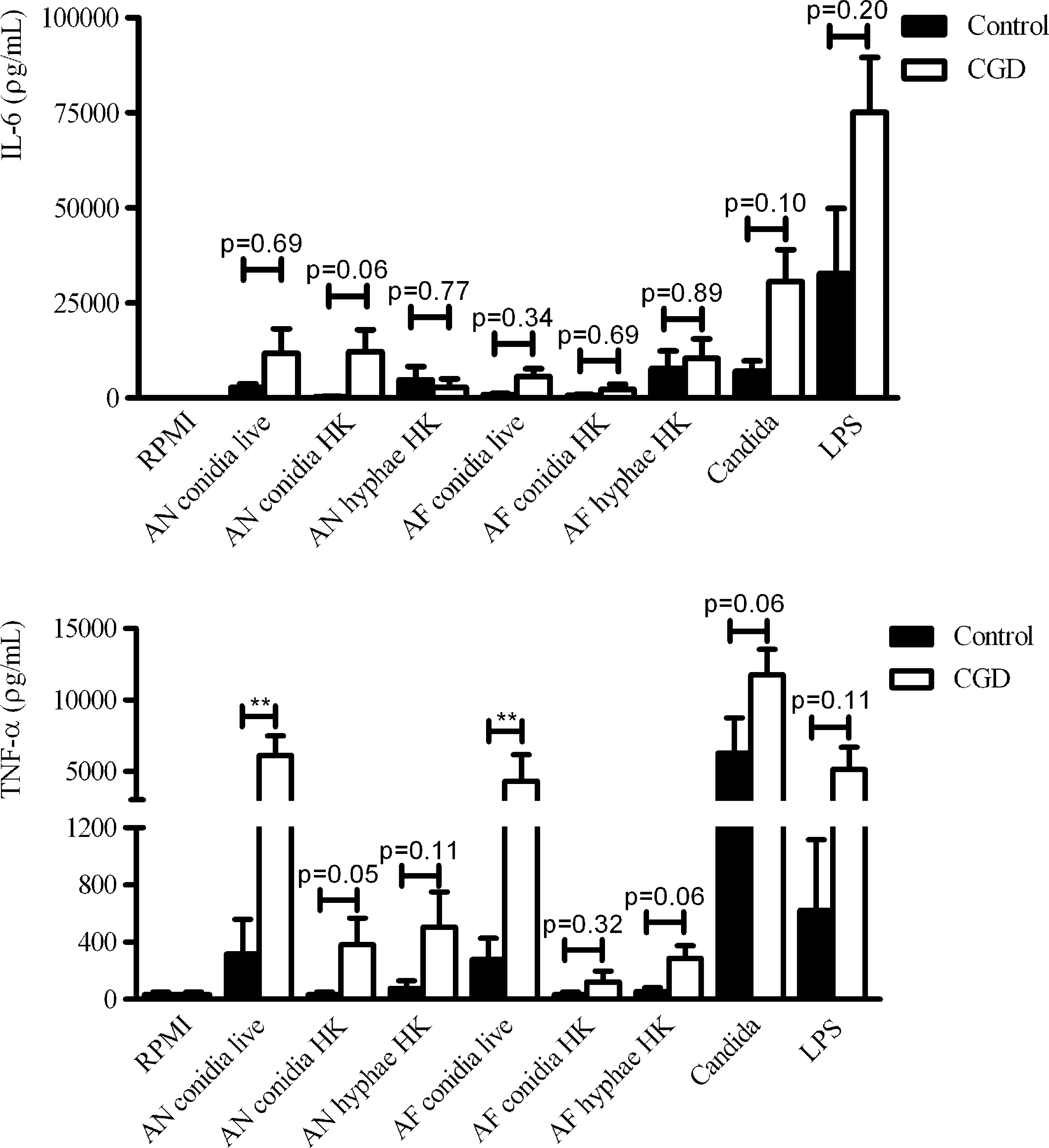
CGD patients show increased production of proinflammatory cytokines upon fungal stimulation in vitro. PBMCs of healthy controls and CGD patients were stimulated for 24 h with Roswell Park Memorial Institute (RPMI) Medium, live and heat-killed (HK) Aspergillus nidulans (AN) and Aspergillus fumigatus (AF) conidia or hyphae, heat-killed Candida albicans (1×106/mL), or LPS (10 ng/mL). IL-6 and TNF-α were measured in cell culture supernatants using ELISA. Data represent 5 CGD patients and 5 healthy controls from 3 different experiments. Data were analyzed using the Mann–Whitney U test (**P<0.01). Bars represent mean + SEM. CGD, chronic granulomatous disease; PBMCs, peripheral blood mononuclear cells; LPS, lipopolysaccharide; TNF-α, tumor necrosis factor-α; IL-6, interleukin-6; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of the mean.
CGD patients have an intact L-tryptophan metabolism
Romani and others (2008) showed that p47phox-deficient mice have a hyperinflammatory phenotype due to a defect in the conversion of
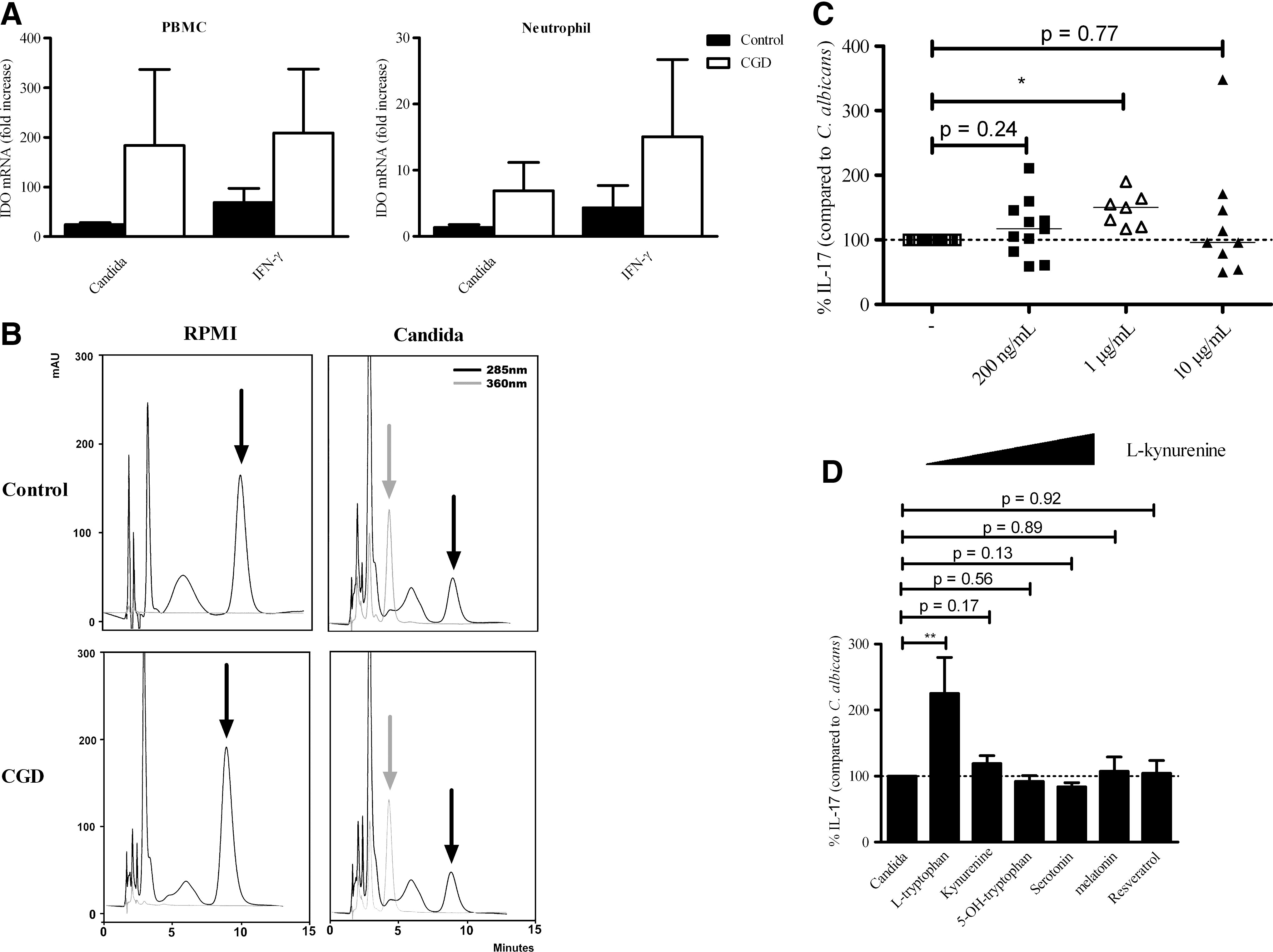
CGD patients do not have a defective
The tryptophan metabolite
CGD patients have a low IL-17A and high IFN-γ production upon stimulation with fungal pathogens
To investigate whether IL-17A production is affected in cells from CGD patients, PBMCs from CGD patients and healthy controls were stimulated for 7 days with αCD3αCD28-coated beads, LPS, different morphotypes of A. nidulans and A. fumigates, and heat-killed C. albicans. There was no difference in αCD3αCD28-induced IL-17 production between CGD patients and healthy controls (Fig. 3A). It is well established that especially in hyphal antifungal host defense, rapid and efficient recruitment of neutrophils is important (Bonnett and others 2006). Here we found that in response to A. nidulans hyphae and A. fumigates hyphae, the IL-17A production was relatively low in CGD patients although this was not significant (Fig. 3B). CGD patients produced more IFN-γ in response to all stimuli (Fig. 3B). PBMCs of CGD patients produced significantly less IL-17A upon stimulation with C. albicans (Fig. 3C).
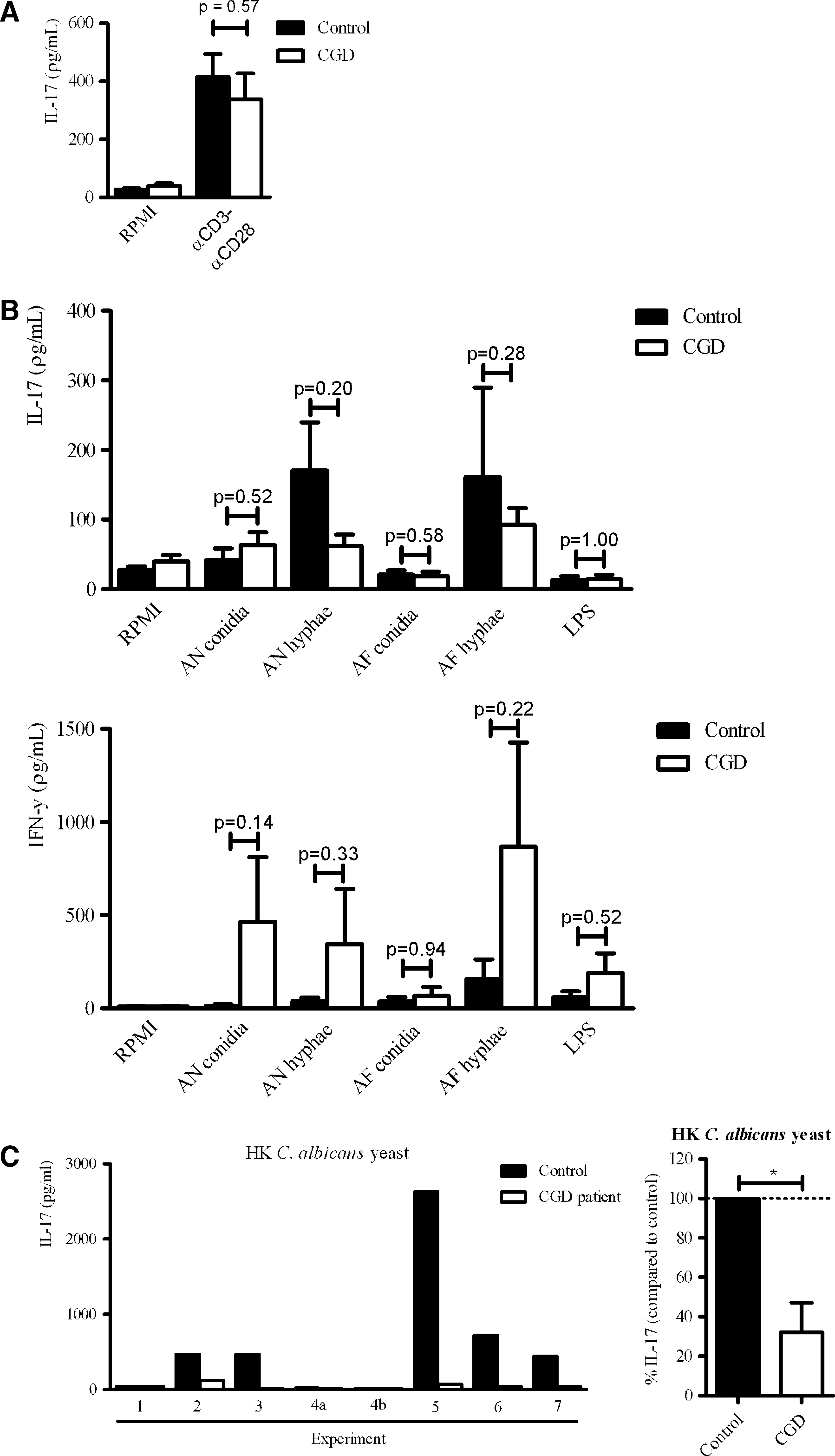
CGD patients have a low IL-17A and high IFN-γ production upon stimulation with fungal pathogens.
Discussion
In the current study we demonstrate that human CGD patients produce markedly lower IL-17A concentrations upon stimulation with the fungal pathogen C. albicans compared with healthy controls, despite an overall higher proinflammatory cytokine production. CGD patients are able to convert
Th17 responses have recently been described as a new T helper subset (McGeachy and Cua 2008). In recent years it has become apparent that the Th17 response is essential for antifungal host defense. The recurrent mucosal Candida infections in patients with primary immunodeficiency syndromes such as HIES and CMC have been specifically linked to a deficiency in their Th17 response (Eyerich and others 2008; Milner and others 2008; van de Veerdonk and others 2011).
The role of the Th17 response in the host defense against Aspergillus is controversial. On the one hand, IL-17 deficiency is associated with a higher susceptibility to Aspergillus infection in mice (Werner and others 2009; Rivera and others 2011). On the other hand, studies suggest that a high IL-17 production is linked to a higher susceptibility to Aspergillus infection due to detrimental effects caused by a hyperinflammatory response (Zelante and others 2007; Romani and others 2008). These observations do not need to be contradictory, and they might support the concept that there is an optimal window for IL-17 response in the host defense against Aspergillus.
The decreased IL-17 production in response to fungal PAMPs was rather unanticipated. We have previously shown that CGD patients have relatively high IL-1β production in response to fungal stimuli (van de Veerdonk and others 2010). IL-1β is a driver of Th17 responses (Chung and others, 2009) and therefore we predicted a high Th17 response. Furthermore, it has been reported that T-cells from CGD patients have increased IL-17 and IFN-γ production in response to mitogenic stimulation, because NADPH oxidase–deficient macrophages cannot sufficiently induce Tregs that are needed to control proinflammatory T helper responses (Kraaij and others 2010). Interestingly we observed no difference in αCD3αCD28-induced IL-17 production between CGD patients and healthy controls, indicating that the decreased IL-17 production observed in CGD patients is only present in the presence of a specific pathogen. This might explain why the rest of the phenotype of CGD patients is not similar to HIES or CMC.
A difference in proportion of γδ T-cells in PBMCs from CGD patients compared with healthy controls could be an explanation for the observed differences in IL-17A production (Roark and others 2008). Therefore we have determined the relative contribution of the γδ T-cells to IL-17A production in response to Candida in healthy volunteers. We found that a very low amount of γδ T-cells was IL-17 positive compared with CD4 T-cells (data not shown). These data suggest that the large proportion of IL-17 in response to C. albicans is from the CD4 T-cell population. Since CD4 lymphopenia is not described in CGD patients, it is unlikely that differential T-cell counts are responsible for the low IL-17A production in CGD patients.
It is interesting to observe that both p47phox-deficient mice and patients with CGD are susceptible to Aspergillus infections, but the underlying mechanisms seem to differ. In mice it was suggested that the tryptophan metabolism was defective, which led to an increased level of IL-17 production that was detrimental for the mice. Here we confirm that in CGD patients the tryptophan metabolism is intact (De Ravin and others 2010), and moreover we observed that they had a lower IL-17 production in response to fungal stimulation. In addition,
In contrast to a low IL-17A production, we observed that the cells from CGD patients have an increased IFN-γ production compared with healthy controls in response to fungal stimuli. IFN-γ stimulates intracellular killing of C. albicans conidia by PMN in vitro (Djeu and others 1986; Kullberg and others 1993), and in a recent overview it has been reported that a large proportion of patients with defects in IL-12Rβ1 experience fungal infections (de Beaucoudrey and others 2010). However, both IFN-γ and IL-17 are required for an adequate antifungal host defense. Although there is no defect in IFN-γ production, the low IL-17A production might cause a sensitivity to fungal infections. CGD patients also have an increased susceptibility to S. aureus infection. Susceptibility to S. aureus has also been associated with a defective IL-17A response (van de Veerdonk and others 2009; Cho and others 2010), which is in line with the data presented here.
In conclusion, human CGD patients have a specific lower production of fungal pathogen-induced IL-17A production compared with healthy controls, while mitogenic stimulation of T-cells isolated from CGD patients does not result in an impaired IL-17 production. This might explain their inability to clear fungal infections. Furthermore, in contrast to a mouse CGD model, human CGD patients do not have a defect in their
Footnotes
Acknowledgments
M.G. Netea was supported by a Vici grant of the Netherlands Organization for Scientific Research (918.10.610). S.S.V. Henriet was supported by the ESPID-Wyeth Fellowship 2008–2010.
Author Disclosure Statement
No competing financial interests exist.