
Abstract
Currently, products containing interferon beta (IFNβ) are injected either intramuscularly or subcutaneously. To avoid the necessity of injection, we developed a novel monomeric Fc fusion protein of IFNβ (IFNβFc) that is absorbed via an immunoglobulin transport system present in the upper and central airways upon administration of the drug as an inhaled aerosol. The systemic absorption of IFNβFc through the lung in non-human primates, at deposited doses of 1, 3, and 10 μg/kg, was compared to the absorption of a single 3 μg/kg dose of IFNβ-1a (Avonex®) subcutaneously administered. IFNβFc was well absorbed through the lung, displaying dose proportional increases in serum concentrations, and was biologically active, as shown by increases in plasma neopterin levels. The circulating half-life of IFNβFc was ∼3 times longer (∼30 h) than that of IFNβ-1a, (8–9 h). At approximately equimolar doses of IFNβFc (10 μg/kg) and IFNβ-1a (3 μg/kg), the stimulation of neopterin over background levels was approximately equivalent, demonstrating that the longer half-life of IFNβFc compensated for the lower relative specific antiviral activity of IFNβFc measured in vitro. In conclusion, IFNβFc was efficiently absorbed after pulmonary delivery in non-human primates, retained its biological activity, and may offer a convenient alternative to injectable IFNβ.
Introduction
I
FcRn is also present in the lung, in bronchial epithelial cells (Bitonti and others 2004; Sakagami and others 2006). FcRn acts as a transporter from the lumen of the upper and central airways to the blood allowing pulmonary delivery of Fc-containing proteins. Pulmonary delivery of a long acting IFNβ could be an important improvement in the treatment of MS. Pulmonary administration of IFNβ-1a was previously tested using intratracheal instillation (Martin and others 2002). No toxicity was observed but the use of instillation is not practical in human subjects, therefore aerosol formulations and appropriate inhalation devices are necessary.
The cynomolgus monkey is one of the few animals in which human IFNβ-1a has been demonstrated to be biologically active (Mager and others 2003). Systemic administration of IFNβ-1a to monkeys results in an increase in serum neopterin concentrations (Pepinsky and others 2001). Neopterin, predominantly produced by the macrophage, is used as a biomarker to monitor IFN biologic activity.
In this study, we administered IFNβFc by pulmonary delivery in non-human primates using a commercially available aerosol generator. We used CHOK1SV cells to produce IFNβFc monomer fusion protein with a 15 amino acid linker of 3 repeating GGGGS units (the “GS15” linker) between IFNβ-1a and the Fc fragment. After purification, the effectiveness of pulmonary delivery of the IFNβFc in monkeys was studied. We monitored IFNβFc pharmacokinetics and the biomarker neopterin by ELISA in serum and in plasma, respectively, from the treated animals. Our results showed that, IFNβFc after pulmonary delivery in non-human primates is as efficient as subcutaneously injected IFNβ-1a; at equimolar doses similar increases in neopterin levels in plasma were observed. The combination of pulmonary delivery and increase of the half-life of IFNβ could be a major advance in treatment of MS patients by facilitating the administration and reducing the frequency of administration.
Materials and Methods
IFNβFc expression
Two Fc fusion protein of IFNβ (IFNβFc) constructs were made. IFNβFc DNA sequence was cloned into pEE 12.4/6.4 vector from Lonza GS Gene Expression System between BSiwI and Rsr II restriction sites. In the first construct, IFNβFc was linked to the Fc fragment of immunoglobulin G1 with a “GS15” linker (GGGGS×3), and in the second construct, IFNβ-1a was directly fused to Fc. For both constructs, CHOK1SV cells were transfected following recommendations from Lonza. Briefly, 1×107 cells were transfected by electroporation in the presence of 40 μg of PvuI-linearized DNA and then cloned by limiting dilution in 96-well microtiter plates. Clones were selected using 50 μM of the GS inhibitor methionine sulfoximine.
Single clones were isolated and IFNβFc expression levels were evaluated by ELISA of culture medium. Positive clones were subcloned and screened for IFNβ expression levels and their ability to grow. Culture medium from the best clone of each construct was used to purify IFNβFc.
IFNβFc ELISA
Culture medium clone screening
Costar microtiter plates were coated for 1 h with 100 μL of 4.5 μg/mL anti-IFNβ rabbit antibody (Biogen Idec Inc.) and then blocked for 1 h with 300 μL per well of phosphate-buffered saline (PBS) (Invitrogen) containing 1% bovine serum albumin (Sigma). Culture medium, diluted 10-fold, was added to the wells and incubated for 1 h at room temperature. After washing with 300 μL of wash buffer: PBS 0.05% Tween 20, plates were incubated for 1 h with 100 μL of 0.17 ng/mL biotin-conjugated anti-IFNβ mouse monoclonal antibody (Biogen Idec Inc.). The wells were then washed with wash buffer and incubated for 30 min with Streptavidin-horseradish peroxidase (HRP) (BD Pharmingen) (1:15,000 dilution) at room temperature. After a second wash, 100 μL of the enhanced K-blue 3,3′,5,5′-tetramethylbenzamidine (TMB) (Neogen) was added for 15 min, and then 100 μL 0.1 M H2SO4 was added to stop the colorimetric reaction. Absorbance of the wells was measured at 450 nm.
IFNβFc preparation and characterization
Purification of IFNβFc
Culture medium from transfected CHOK1SV cells was passed through a 0.2 μm filter and then loaded on a dye affinity chromatography column (GE Healthcare 5-mL HiTrap Blue Sepharose HP) equilibrated with 50 mM HEPES, 100 mM NaCl, pH 7.0. The column was washed with 50 mM HEPES, 100 mM NaCl, pH 7.0, and then with 50 mM HEPES, 200 mM NaCl, 6% propylene glycol, pH 7.0, and IFNβFc was eluted with 50 mM HEPES, 650 mM NaCl, and 36% propylene glycol, pH 7.0. To separate the monomeric and dimeric forms of IFNβFc, the protein was further purified using a size exclusion chromatography (SEC) column (GE Healthcare Superdex 200) equilibrated with 50 mM HEPES, 500 mM NaCl, 1% Trehalose, and 0.005% Tween-20, pH 7.5 at 2°C–8°C. IFNβFc was eluted from the column at 20 cm/h with 50 mM HEPES, 500 mM NaCl, 1% Trehalose, and 0.005% Tween-20, pH 7.5 and the individual elution fractions were evaluated for IFNβFc content using analytical SEC.
Analytical SEC
A TSK G3000SWXL (7.8 mm×30 cm, 5 μm) column from Tosoh Bioscience (Part No. 08541) was used for SEC analysis of IFNβFc. The column was equilibrated with 400 mM sodium perchlorate, 25 mM sodium phosphate (pH 6.5) for 60 min at 1.0 mL/min at room temperature. IFNβFc samples (100 μL) were injected and the column was isocratically run with the same mobile phase for 18 min. Eluted protein was detected by UV absorbance at 214 nm.
Binding of soluble human FcRn to immobilized IFNβFc or Human IgG1 by Biacore
Interaction between either IFNβFc or human IgG1 (Sigma Catalog No. I5154), and soluble human FcRn was measured by Surface Plasmon Resonance using steady-state affinity analysis on a Biacore T100. IFNβFc or Human IgG1 was immobilized on CM5 sensor chips at ∼500–1,000 RU using a 10 or 20 μg/mL solution of protein in 10 mM sodium acetate coupling buffer (Catalog No. BR-1003-50), pH 4.5 using the standard amine kit from Biacore (Catalog No. BR-1000-50). Soluble hFcRn was injected at concentrations ranging from 0.05 to 100 μM at room temperature in pH 6.0 sodium phosphate binding buffer for 10 min at a flow rate of 25 μL/min and dissociation was observed for 10 min. The binding responses recorded near the end of the association phase were plotted as a function of concentration. The equilibrium KD was derived from nonlinear regression analysis of the data from the binding curve (Req versus Concentration). The experimental binding results were fitted to a 1:1 binding model using the Biaevaluation software (BIAevaluation version 4.1; Biacore Instruments).
Pharmacokinetics and pharmacodynamics
Experiments were conducted on 2.5–3.5 kg male and female captive-bred cynomolgus monkeys at New Iberia Research Center (New Iberia, LA). Before the study, neopterin levels in plasma from each monkey were measured and only those animals with concentrations of neopterin<3 ng/mL were chosen. Monkeys were divided into 4 groups of 3 monkeys each. Animals in Groups 1–3 received a single dose of 1, 3, or 10 μg/kg IFNβFc respectively, by pulmonary administration. Monkeys in Group 4 subcutaneously received 3 μg/kg of IFNβ-1a (Avonex). A control group of monkeys administered vehicle only was not used because in previously published work (Martin and others 2002) intracheal instillation of vehicle caused no change in neopterin levels. Blood was collected before dosing and at 1, 4, 8, 24, 72, 96, 120, 144, and 168 h postdose. Serum and plasma were prepared and analyzed for IFNβ levels and neopterin concentrations respectively.
Before dosing, the animals were sedated with either ketamine for subcutaneous injections or with a mix of ketamine and diazepam for pulmonary administration. Animals receiving IFNβFc by the pulmonary route were intubated with a 3–5 mm cuffed endotracheal tube and the cuff was inflated. IFNβFc was administered as an aerosol, generated using an Aeroneb Pro® nebulizer (Aerogen) in-line with a Bird Mark IV respirator. The respirator was used to control respiratory rate, ∼30–32 breaths per min, and depth of respiration (shallow breaths). IFNβFc in 50 mM HEPES (pH 7.5), 500 mM NaCl, 1% trehalose, and 0.02% Tween 20 was loaded into the nebulizer and the nebulizer run for ∼5–7 min to administer a total volume of 3 mL. IFNβFc specific activity was tested before and after nebulization under the same working conditions, and no significant loss of activity was observed. Doses were calculated from the volume of aerosolized IFNβFc solution, the concentration of the IFNβFc solution and the body weight of the animal. Previous work showed that the lung deposition was ∼15% of the nebulizer dose.
Pharmacokinetic and Pharmacodynamic parameters were calculated using WinNonlin Professional version 5.3.
Biologic assays
IFNβ ELISA
IFNβ concentrations in serum were measured using an ELISA kit from Invitrogen (catNo. KAC1201). The kit utilizes the one-step sandwich method. Briefly, microtiter plates were coated with the affinity-purified polyclonal antibody to IFNβ. IFNβ binds to the antibody-coated plate. The HRP-labeled secondary anti-IFNβ monclonal antibody also binds to form an antibody-antigen complex where the IFNβ is sandwiched between the primary antibody immobilized on the well and the enzyme-labeled secondary antibody. Finally, an enzyme reaction system is activated by adding a color developer. The IFNβ bound is quantified by spectrophotometric measurement of the chromogenic product. Samples were diluted 1:5, 1:10, or 1:20 in sample buffer contained in the kit.
Antiviral activity assays
The activity of IFNβFc was determined by testing the protein samples for antiviral activity using a cytopathic effect assay (Alam and others 1997). In this assay, human lung carcinoma A549 cells (ATCC, CCL-185: 1.6×104 cells/well) were plated into 96-well plates. After 5 h, serial dilutions of IFNβFc test samples were added, and 18 h later the cells were challenged with EMC virus. Dilutions of an IFNβ international standard (WHO international standard 00/572, NIBSC: 10, 5.7, 3.3, 1.9, 1, 0.6, 0.3, and 0.2 IU/mL) were assayed on every plate. Viable cells were quantified 28 h after viral challenge by addition of sulforhodamine B dye and measurement of absorbance at 490 nm. Standard curves generated on each plate were used to determine the IFNβ activity in each test sample.
Neopterin ELISA
Biological responses to IFNβFc in cynomolgus monkeys were assessed by measuring the plasma neopterin levels using a commercial neopterin ELISA kit [Immuno-Biological Laboratories (IBL), Inc.]. In this assay, an unknown amount of neopterin in the plasma and a fixed amount of HRP-conjugated neopterin compete for binding sites on a mouse monoclonal antibody against neopterin. Both the antigen and the monoclonal antibody complex bind to the wells of the microtiter strips coated with goat anti-mouse antibody as recommended by IBL. Briefly, a fixed volume of plasma, HRP-conjugated neopterin, and mouse monoclonal antibody were added into each well and incubated in the dark for 90 min at room temperature. The incubation mixture was then decanted and plates were washed 3 times with wash buffer. TMB substrate solution was then added to each well and the plates were incubated in the dark for 15 min. The substrate reaction was stopped with 1N HCP. The plates were optically measured by reading the absorbance at 450 nm.
Results
IFNβFc synthesis and characterization
The C-terminus of IFNβ-1a was fused to the Fc fragment of IgG1 using either a 15 amino acid linker of 3 repeating GGGGS units (the “GS15” linker) or no linker sequence. Single Fc fragments were co-expressed with the IFNβFc. In this expression system, the IFNβFc can bind either to another IFNβFc, to form a homodimer, or to a single Fc fragment, to form a heterodimer. We have designated the IFNβFc linked to one Fc as IFNβFc monomer and the homodimer of 2 IFNβFc components as IFNβFc dimer (Fig. 1A). By SEC, IFNβFc monomer was separated from the IFNβFc dimer and from the Fc dimer (Fig. 1B). IFNβFc monomer represented ∼46% of the protein loaded onto the preparative SEC column and the pooled column fractions containing IFNβFc monomer (99.4% pure) comprised ∼8% of the total area under the chromatogram. Therefore, recovery of IFNβFc monomer in this purification step is ∼17%.
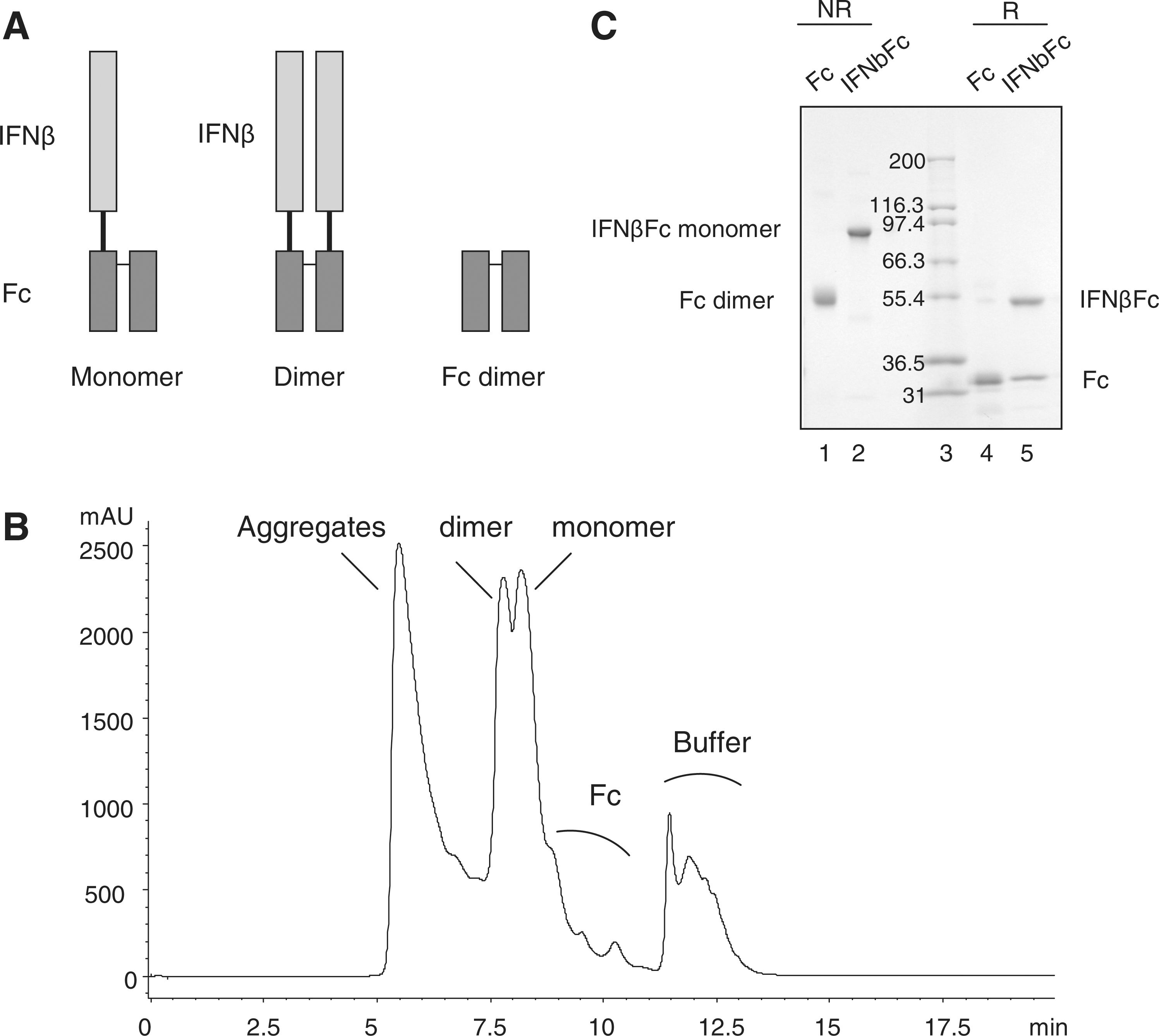
Analysis of IFNβFc by SDS-PAGE and SEC.
The acrylamide gel in Fig. 1C shows Fc alone and the IFNβFc monomer in reduced and nonreduced conditions. Under nonreduced conditions, the species identified are an Fc dimer of 50 kDa and an IFNβFc monomer of 74 kDa (IFNβFc 49 kDa plus one Fc 25 kDa). Under reduced conditions, the 2 proteins are separated from each other. Fc migrated at 25 kDa and IFNβFc monomer at 49 kDa. Monomer and dimer Fc fusion protein antiviral activities were compared in a cytopathic effect assay and the monomer form was shown to be ∼4 times more active (Bitonti and Dumont 2006). Therefore, only the IFNβFc monomer was used for subsequent experiments.
IFNβFc monomer was tested for its ability to inhibit the cytopathic effect of EMC virus on A549 cells. Using an international standard as a reference, the specific activity of IFNβFc with the GS15 linker was 3.94×107±0.62 IU/mg (n=5). In the same test, Avonex drug product was 2.8×108±0.65 IU/mg (n=5). On a molar basis, the addition of the Fc fragment reduced the specific activity by 2.3-fold.
To evaluate whether the linker would interfere with binding to the Fc receptor, FcRn, the affinities of 2 IFNβFc constructs, one without and one with the GS15 linker, were determined by Biacore. The Biacore results showed no difference of affinity for human FcRn between the 2 IFNβFc proteins (Kd=7 μM for both).
Pharmacokinetics and pharmacodynamics in monkeys
Serum IFNβ level
Pharmacokinetic parameters increased approximately linearly with dose after pulmonary delivery, with a Cmax of 1.15±0.73, 3.9±0.65, and 17.92±6.13 ng/mL for the 1, 3, and 10 μg/kg doses, respectively (Fig. 2 and Table 1). Following subcutaneous administration of 3 μg/kg Avonex, the maximum IFNβ serum concentration was 3.66±0.92 ng/mL, comparable to the value attained after 3 μg/mL of IFNβFc. After equimolar doses, [IFNβFc at 10 μg/kg (0.135 nmol/kg) and Avonex at 3 μg/kg (0.125 nmol/kg)] the Cmax and area under the curve (AUC) were 4.9 and 4.4-fold greater, respectively, for IFNβFc than IFNβ (Table 1). The addition of the Fc fragment increased the serum half-life by about 3-fold: 9.9±1.4 h for Avonex to 27±5.7 h for IFNβFc.
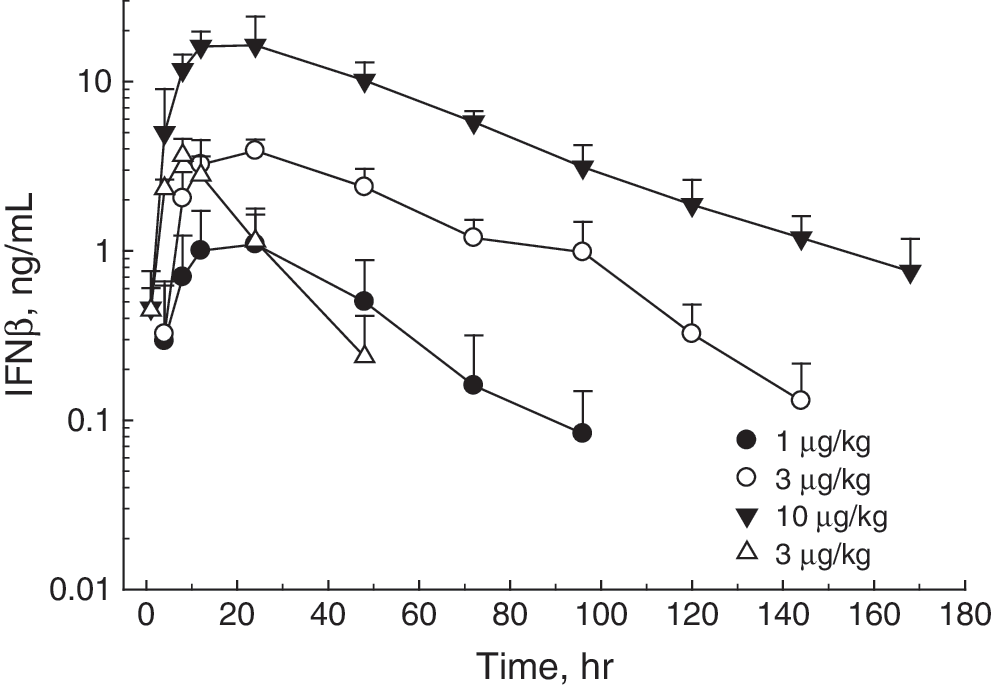
Pharmacokinetics of IFNβ subcutaneously injected and IFNβFc administered via inhalation in non-human primates. Blood from animals was collected before dosing and at 1, 4, 8, 24, 72, 96, 120, 144, and 168 h postdose. Serum IFN levels were measured by ELISA. Data points are means of values from 3 monkeys±the standard deviation. IFNβ subcutaneous 3 μg/kg (Δ); IFNβFc pulmonary at 1 μg/kg (●); 3 μg/kg (○); 10 μg/kg (▾).
t max are expressed in median with range shown in parentheses.
IFNβFc, Fc fusion protein of interferon beta; s.c., subcutaneous.
Plasma neopterin levels
Cynomolgus monkeys are responsive to human IFNβ (Mager and others 2003). In vivo biological activity can be monitored by measuring the biomarker, neopterin. Plasma neopterin levels (Fig. 3) increased after IFNβ subcutaneous injection and also after pulmonary delivery of IFNβFc. IFNβFc administered via pulmonary delivery was absorbed slower than IFNβ-1a administered via subcutaneous injection, resulting in a later Tmax (median at 48 h versus median at 24 h) (Table 2). At the 1 μg/kg dose of IFNβFc, the plasma levels of neopterin considerably varied between animals. At 3 and 10 μg/kg, the plasma concentrations were more consistent but the area under the effect curve (EAUC0-168h) values showed significant variation, with standard deviations of 21% to 55% (Table 2). Nevertheless, a dose-response effect was observed from 1 to 10 μg/kg in EAUC0-168h and neopterin maximal effect concentration (Emax), but the neopterin increase with dose was in a less than dose-proportional manner. With a 3-fold increase in IFNβFc dose from 1 to 3 μg/kg, EAUC0-168h of neopterin increased only by 1.6-fold with no variation in the Emax. With a further 3-fold dose increase from 3 to 10 μg/kg, EAUC0-168h and Emax of neopterin increased by 1.9-fold (Table 2). The lack of higher neopterin response, after drug exposure increase, has been previously reported (Pepinsky and others 2001; Hu and others 2011), certainly due to the fact that the IFNβFc serum concentrations were much higher than the EC50: 0.38 ng/mL (IFNβFc serum concentration to achieve 50% of the Emax).
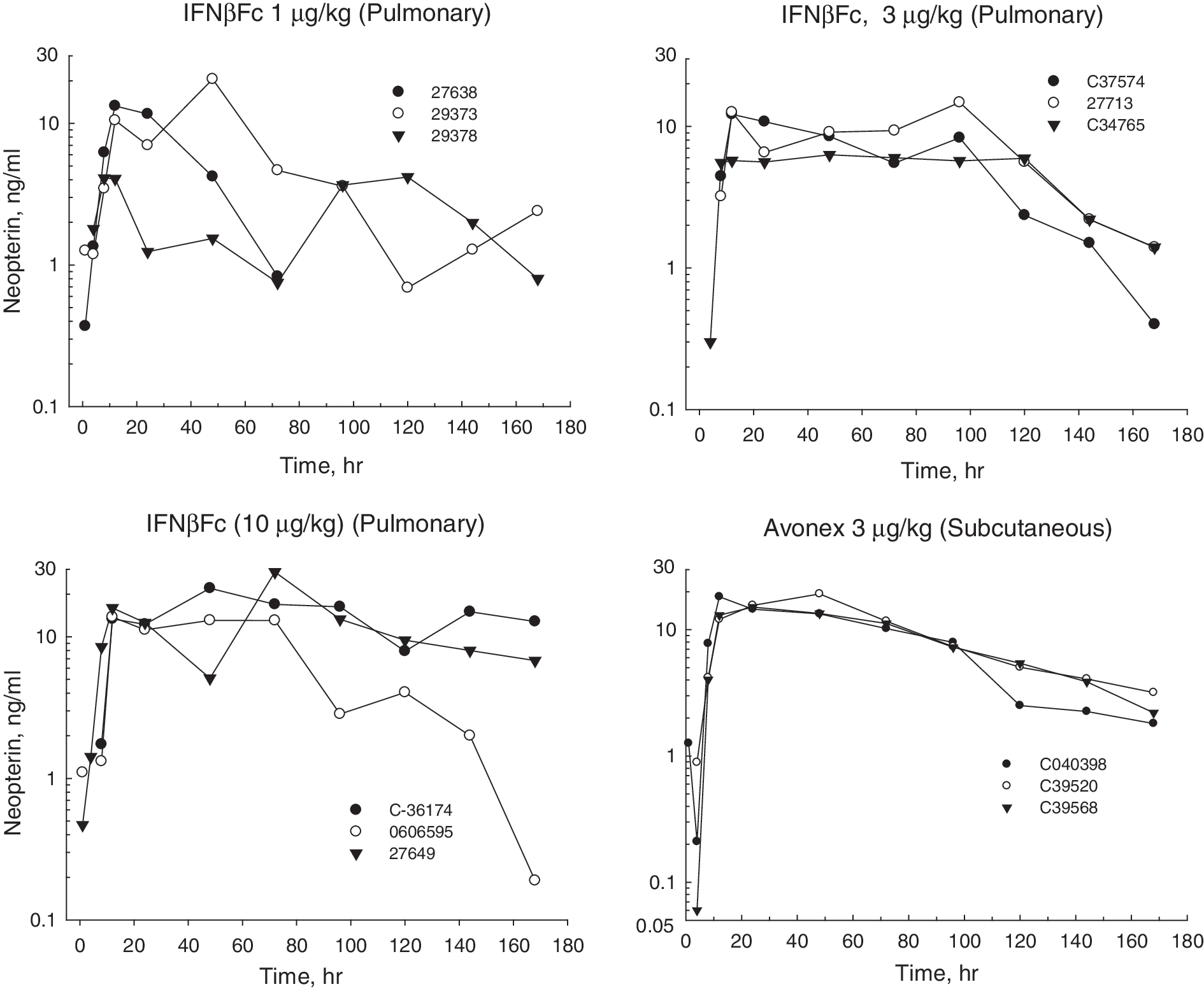
Changes in plasma neopterin concentrations in non-human primates after administration of IFNβFc by inhalation and IFNβ by subcutaneous injection. Neopterin concentrations were measured by ELISA. Each graph represents a group of 3 monkeys treated with either 1, 3, or 10 μg/kg IFNβFc or 3 μg/kg IFNβ.
Discussion
The results of these studies show that Fc fusion proteins can be delivered to the systemic circulation by binding to FcRn, an IgG receptor used as a transport pathway from the upper/central airways to the blood. It was of interest to test a new IFNβFc fusion protein with a GS15 (GGGGS×3) linker for pulmonary delivery in non-human primates. An IFNβFc fusion protein, with a random 8 amino acids linker between IFNβ and the Fc fragment of IgG1, was previously described (Bitonti and Dumont 2006). The new construct was designed with a longer linker to potentially decrease the proximity effect between IFNβ and Fc that could affect their respective functionalities, i.e., IFNβ efficacy and Fc binding to its receptor, FcRn. To evaluate the effect of the linker, we expressed 2 IFNβFc proteins with the same cellular expression and protein purification system; one without a linker and one with the GS15 linker. When the 2 IFNβFc proteins were compared for their affinities for FcRn, there was no apparent difference. Second, specific antiviral activities of the 2 proteins were compared. In the absence of the linker, the activity was approximately the same as that of the protein with the GS15 linker (data not show).
The IFNβFc with the GS15 linker selected as the lead candidate and was tested in a pulmonary delivery experiment in non-human primates to offer an alternative to injectable IFNβ.
Pharmacodymamic and pharmacokinetics parameters were determined for IFNβFc after dosing monkeys with either 1, 3, or 10 μg/kg (lung deposited dose) by pulmonary delivery. For comparison, another group of monkeys were subcutaneously dosed with 3 μg/kg Avonex.
Interestingly, the Cmax obtained after pulmonary administration of 3 μg/kg of IFNβFc, is similar to that obtained after subcutaneous injection of 3 μg/kg of Avonex. After injection of equivalent molar doses (3 μg/kg of Avonex and 10 μg/kg IFNβFc), the Cmax for IFNβFc increased by 4.9-fold and the AUC increased by 4.4-fold. The IFNβ serum half-life increased by about 3-fold. Thus, Fc technology not only enables pulmonary delivery, but also significantly increases the circulating half-life. The half-life for IFNβFc following pulmonary delivery to cynomolgus monkeys (27 h) was comparable to that obtained with 20 kDa PEGylated forms of IFNβ-1a following subcutaneous administration (22 h, Pepinsky and others 2001), subcutaneous and intramuscular administration (∼20 h, Xu and others 2011), and 40 kD PEGylated forms subcutaneous (up to 21H, Mager and other 2005) to rhesus monkeys.
In terms of biological activity, increases in plasma levels of the biomarker neopterin were similar when equimolar doses of IFNβFc (10 μg/kg) and IFNβ-1a (Avonex) (3 μg/kg) were administered by either pulmonary delivery or subcutaneous injection, respectively. A maximal neopterin response to exogenous IFNβ may have been achieved in this experiment. Between 3 and 10 μg/kg of IFNβFc, the neopterin concentration did not increase further, however the neopterin concentration remained elevated for a longer period of time. The same phenomenon was previously observed, with a lack of increase in the magnitude of the response in rhesus monkeys following administration of PEGylated IFNβ-1a due to the IFNβ-1a blood concentration being close to the Emax concentration (Pepinsky and others 2001; Xu and others 2011). In our studies, IFNβFc was administered at doses of 0.04, 0.12, and 0.4 MIU/kg, but, the measured levels of neopterin were higher. As we prescreened our monkeys for low basal levels of neopterin, we may have allowed selection of more sensitive monkeys. In any case, it appears that a maximal neopterin response can be achieved with 3 μg/kg of IFNβFc or Avonex. These increased neopterin levels were prolonged when the IFNβFc dose was increased to 10 μg/kg. At equimolar doses, the half-life of neopterin after IFNβFc inhalation was comparable to that observed after Avonex injection.
The impact of chronic IFNβ administration to the lung could be a concern. The lungs of monkeys were not examined in this study, but daily doses of intratracheal instillation of IFNβ at 60 μg (10 μg/kg) for one week did not induce any toxicity (Martin and others 2002). In our experiment, the 10 μg/kg deposition dose represented ∼30 μg total lung deposited dose per monkey. Because IFNβFc has an improved long half-life, the frequency of administration may be less than one week intervals at the same dose. Under these conditions, the lung exposure would be at least 7 times lower than that used in the toxicity studies of inhaled IFNβ-1a. While the methods of delivery, the doses of IFNβ, and the composition of the IFNβ are not identical in the experiments previously published by Martin and others and our experiment, the lack of toxicity in the previous work is at least suggestive that IFNβFc may be administered via the lung with some margin of safety.
Many Fc fusion proteins have been studied in humans and monkeys (Bitonti and Dumont 2006) and this technology is well established. Delivery to the upper/central airways is a convenient pathway for efficient systemic delivery via inhalation. This study shows that monomer Fc fusion technology can be successfully applied to IFNβFc, allowing therapeutic dosing by inhalation with similar efficacy to the doses of Avonex used for subcutaneous injections and allows also less frequent dosing because of the subsequent increase in half-life.
Footnotes
Author Disclosure Statement
Sebastien Vallee, Swapnil Rakhe, Thomas Reidy, Sandra Walker, Qi Lu, Paul Sakorafas, and Alan Bitonti are employed by Biogen Idec Hemophilia and Susan Low is a former employee of Biogen Idec Hemophilia.