
Abstract
The roles of inflammation-associated genes in the pathogenesis of endocervical polyps remain unclear. We thus compared the expression levels of 509 inflammation-associated genes between endocervical polyp tissues and endocervical canal membrane tissues using a gene microarray. Sixteen inflammation-related genes were differentially expressed in endocervical polyps compared with those of normal endocervical canal membrane tissues. Expression of 8 of these 16 genes was further validated biochemically. The protein expression levels of IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, and IL-11 were significantly higher in endocervical polyps than those in endocervical canal tissues, while expression of TGF-β1 and IL-10 was significantly lower (P<0.05). In addition, endocervical polyp tissues expressed IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, IL-11, TGF-β1, and IL-10 mainly in the cytoplasm of the inflammatory cells and, to a lesser extent, in the acinus of the serous gland. Endocervical polyp is a polygenic disease and aberrantly expressed genes may play roles in its pathogenesis.
Introduction
C
Inflammatory endocervical polyps are associated with chronic mucosal inflammation of the cervical membrane and the focal inflammation induces hyperplasia of the endocervical canal. The inflammatory endocervical polyp is populated with the surface epithelium containing columnar and/or squamous epithelium, microglandular cells, as well as activated macrophages/monocytes, eosinophils, neutrophils, and plasma cells. In contrast, normal endocervical tissue harbors few activated macrophages/monocytes, eosinophils, neutrophils, and plasma cells. The critical factors that stimulate the hyperplasia remain unclear, but infection and abnormal hormonal stimulation have been proposed. The innate immune system plays important roles in mucosal defense in cervical neoplasms (McCluggage 2004). Most previous studies investigated the pathogenesis of cervical polyps by examining a single or several factors, but not systematically investigating hundreds of genes. Clearly, the etiology of these polyps can involve complex interactions between numerous cell types and their biochemical mediators, which poses a considerable challenge to determine what factors are most important for pathogenesis.
DNA microarray technology now can quantitatively monitor the expression of numerous genes simultaneously (Hoffmann and others 2001). However, few studies of host responses to infection have been carried out for endocervical polyps with specific immunological DNA microarrays. In particular, no study has published the inflammation-related genes expressed in endocervical polyps as detected with DNA microarrays. One of our main interests is to dissect immune responses present in endocervical polyps. In our research, we have described a microarray that can be used to analyze the changes in gene expressions of tissues in endocervical polyps.
We hypothesized that inflammation-associated genes would be differentially expressed in tissues from endocervical polyps when compared with normal cervical mucosa. We chose normal cervical mucosa rather than blood as the control because we were interested in identifying those inflammation-related genes that were differentially expressed locally in the cells of the polyps. In this study, we compared gene expression in endocervical polyps with that of endocervical canal membrane tissue by using our gene chip with 509 inflammation-associated genes, which included proinflammatory factors, antiinflammatory factors, chemotactic factors, cytokines, and their receptors. Differentially expressed genes that were most likely relevant to cervical polyps included IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, IL-11, TGFβ1, and IL-10; they were chosen to validate the microarray analyses. Protein expression of these genes was further examined by using Western blot analysis and immunohistochemical staining.
Materials and Methods
Specimens
All patients signed a written informed consent for use of their samples. Specimens were collected from patients undergoing endocervical polyp hysteroscopic resections in the Department of Gynecology and Obstetrics of the First Affiliate Hospital of Harbin Medical University, Harbin, China. Endocervical polyp tissues were obtained from 8 patients aged 23 to 65 years old (average age 49 years). Control tissues used for DNA microarray assays were normal cervical canal mucous membranes from 8 patients aged 24 to 52 years (average age 42 years). All samples were immediately placed in separate centrifuge tubes on dry ice and stored at −80°C until used for microarray studies. The 8 cases with endocervical polyps included 1 subject with recurrent polyps, 2 with multiple polyps, and the remaining 5 isolated polyps. Polyp sizes ranged from (1.0 cm×0.8 cm×0.5 cm) to (5 cm×4 cm×2.5 cm).
For experimental verification using immunohistochemistry and Western blot analysis, additional cervical polyp tissues and normal endocervical canal mucous membrane tissues were obtained from inpatients of the Department of Gynecology and Obstetrics of the First Affiliated Hospital of Harbin Medical University. Endocervical polyp tissues were obtained from 50 patients aged 18 to 78 years (average age 41). Normal cervical canal mucous membrane tissues were obtained from 38 patients aged 20 to 75 years (average age 35 years) to avoid any potential contamination of the normal tissues with a few cells from the cervical polyps. Fresh tissues (about 1 cm3) were frozen immediately in liquid nitrogen after resection. Among the 50 cases with cervical polyps, 3 had recurrent polyps, 5 had multiple polyps, and the remaining 37 had single polyps. The polyp sizes ranged from (1.0 cm×1.0 cm×0.5 cm) to (6 cm×5 cm×3 cm).
All patients with endocervical polyp had not received hormonal therapy within one month before the procedures. The inflammatory polyps were identified by pathological diagnosis.
Microarray analysis
Total RNAs were isolated from 8 pooled endocervical polyps using an RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. Pooled polyp RNA samples (100 ng) were reverse transcribed and used to generate Cy5-UTP–labeled cRNA using a low RNA input linear amplification kit (Agilent Technologies, Inc.). Likewise, RNA samples extracted from 8 pooled normal endocervical canal membrane tissues were used to synthesize cRNA that was labeled with Cy3-UTP. Labeled cRNAs from endocervical polyps and control samples were mixed and analyzed for gene expression on 3 microarray chips (509 cytokine-related genes) purchased from CapitalBio.
Hybridization was performed in a SciGene 4000 HybOven (SciGene Corp.) at 65°C for 18 h in a rotary chamber at 10 rpm. Slides were washed according to the manufacturer's instruction (Agilent Technologies, Inc.) and scanned with a dual-confocal continuous microarray scanner (GenePix 4000B; Molecular Devices). GenePix Pro version 6.1 was used for image acquisition. Microarray data were also imported into Spotfire Decision Site for functional genomics (Spotfire Inc.), filtered, and subjected to statistical analysis.
Western blot analysis
Frozen tissues were ground individually with liquid nitrogen, and proteins were extracted. Protein concentrations were determined using the bicinchoninic acid method (BCA kit). Total protein (50 μg) was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane (Amersham Life Science). After blocking with 5% skim milk in 1×TBST [0.01 M Tris (pH 7.6), 0.1 M NaCl, and 0.1% Tween-20], the membrane was incubated with the indicated primary antibodies diluted at 1:2,000 (antihuman IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, IL-11, TGF-β1, and IL-10; all from Abcam), respectively. After addition of the HRP-conjugated secondary antibody, protein bands were developed by using the Enhanced Chemiluminescence (ECL Plus) system (Hongbo Bioscience). Four replicates were analyzed for each protein for each of the 50 endocervical polyps and 38 endocervical canal membrane tissues with β-actin as a loading control (Santa Cruz Biotechnology). The Western blot results were scanned and the signal relative to β-actin was quantified using ImageJ.
Immunohistochemistry
Microarray-identified candidate proteins in endocervical polyps were selected for analysis with well-characterized specific antibodies. The samples were fixed in 4% paraformaldehyde at 4°C overnight and then rinsed with water to remove excess paraformaldehyde. Samples were dehydrated in 50% ethanol for 2 h, followed by 70% ethanol for 2 h, and then stored at room temperature in 70% ethanol. After rehydration in water for 2 h, samples were dehydrated in a graded alcohol series and then embedded in paraffin for histological analysis. To unmask antigens, 5-μm sections were boiled in Target Retrieval solution (Dako) in a pressure cooker. The slides were incubated overnight at 4°C with polyclonal antibodies against IL-12p40, IL-17, IFN-γ, TNF-α, CCR2, IL-11, TGF-β1, and IL-10 (Abcam) in replicates of 4 for each polyp and normal cervical samples. Each antibody was diluted at 1:100 in 0.1 M of phosphate-buffered saline (PBS) solution (pH 7.4). PBS solution without the primary antibodies was used for negative controls. Immunohistochemical staining was performed with a streptavidin peroxidase kit (Hong Bo Company). Antigens were visualized with diaminobenzidine. Subsequently, tissues were counterstained with hematoxylin and eosin, dehydrated, and mounted. Positive staining for IL-12p40, IL-17, IFN-γ, TNF-α, CCR2, IL-11, TGF-β1, and IL-10 appeared as brown color. Negative controls showed no staining. Protein expression in endocervical polyps and cervical canal membranes was examined. Cell counts from photomicrography were quantified (cells per mm of basement membrane or mm3 of tissue) using Image J software and the quantification was blinded to the sample identification.
Statistical analysis
Data were presented as means±standard deviations (SDs). Differences of gene expressions between endocervical polyps and endocervical canal tissues were tested by student's t-test. The significance level was set at 0.05. Statistical analyses were performed using SAS 9.1 (SAS Institute, Inc.).
Results
Hybridization signals on gene chips
RNAs extracted from endocervical polyps were used to generate cRNAs labeled with Cy5 whereas that of endocervical canal membrane tissues were labeled with Cy3. These labeled samples were hybridized to 3 microarray chips containing 509 inflammatory genes, as shown by the representative samples in Fig. 1. Differentially expressed genes were identified by the ratios of Cy5 staining (polyp tissue) to Cy3 (control tissue) signal (Fig. 2). Genes with Cy5/Cy3 ratios between 0.5 and 2.0 were depicted as closed circles in Fig. 2 and were not considered to be differentially expressed. In contrast, genes with Cy5/Cy3 ratios either<0.5 or>2.0 were considered to be differentially expressed and were depicted with open circles. A total of 16 genes met the criteria for differential expression on all 3 chips.

Representative results of microarray analysis. RNAs extracted from endocervical canal membranes (left panel) and endocervical polyps (right panel) were reverse transcribed to generate cRNAs that were labeled with Cy3-UTP and Cy5-UTP, respectively. These labeled samples were hybridized to microarray chips containing 509 inflammatory genes.

Scatter plot of relative RNA levels of inflammation-associated genes in cervical polyps versus normal cervical mucosa. Each circle depicts the mean value of the relative expression of one of the 509 genes in endocervical polyp tissue (Cy5) and normal cervical mucosa (Cy3) on the 3 microarray chips. Open circles: genes with Cy5/Cy3 ratios either<0.5 or>2.0. Closed circles: genes with Cy5/Cy3 ratios between 0.5 and 2.0.
Differential gene expression
Among the 16 genes that consistently showed differential expression, 6 genes had higher mRNA levels (fold-change>2.0) in endocervical polyps than in controls whereas 10 genes were downregulated (fold-change<0.5), as shown in Table 1. The expression of the 6 upregulated genes (IL-12p40, IL-17, IFN-γ, TNF-α, CCR2, and IL-11) was 2.5–4.6-fold higher in polyps than in control tissue. The 10 downregulated genes (TGF-β1, TNFRSF11A, CD86, IL-15, IL-22, IL-2, IFN-β1, IL-1β, IL-10, and CCL-15) have lower expression in polyps (0.32–0.44-fold) than in the control samples.
Consistently upregulated and downregulated genes of pooled cervical polyps compared with normal endocervical canal membrane tissues were identified in 3 microarray analyses.
Mean fold change of the 3 microarray analyses.
Western blot analysis
To validate the microarray data, 8 genes were chosen to further characterize their expression at the protein level. Protein levels of 6 upregulated genes (IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, and IL-11) and 2 downregulated genes (TGF-β1 and IL-10) were analyzed by Western blots. Significant differences were observed in the relative expression of these inflammation-related proteins between endocervical polyps and endocervical canal membranes (P<0.001) (Fig. 3A, B). Endocervical polyps expressed higher levels of IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, and IL-11 than control samples. In contrast, the relative protein expression of TGF-β1 and IL-10 was lower in endocervical polyps than that in control samples. The results from analyzing the protein levels of these 8 differentially expressed genes were consistent with their differentially expressed RNA levels in the endocervical polyps and control tissues.
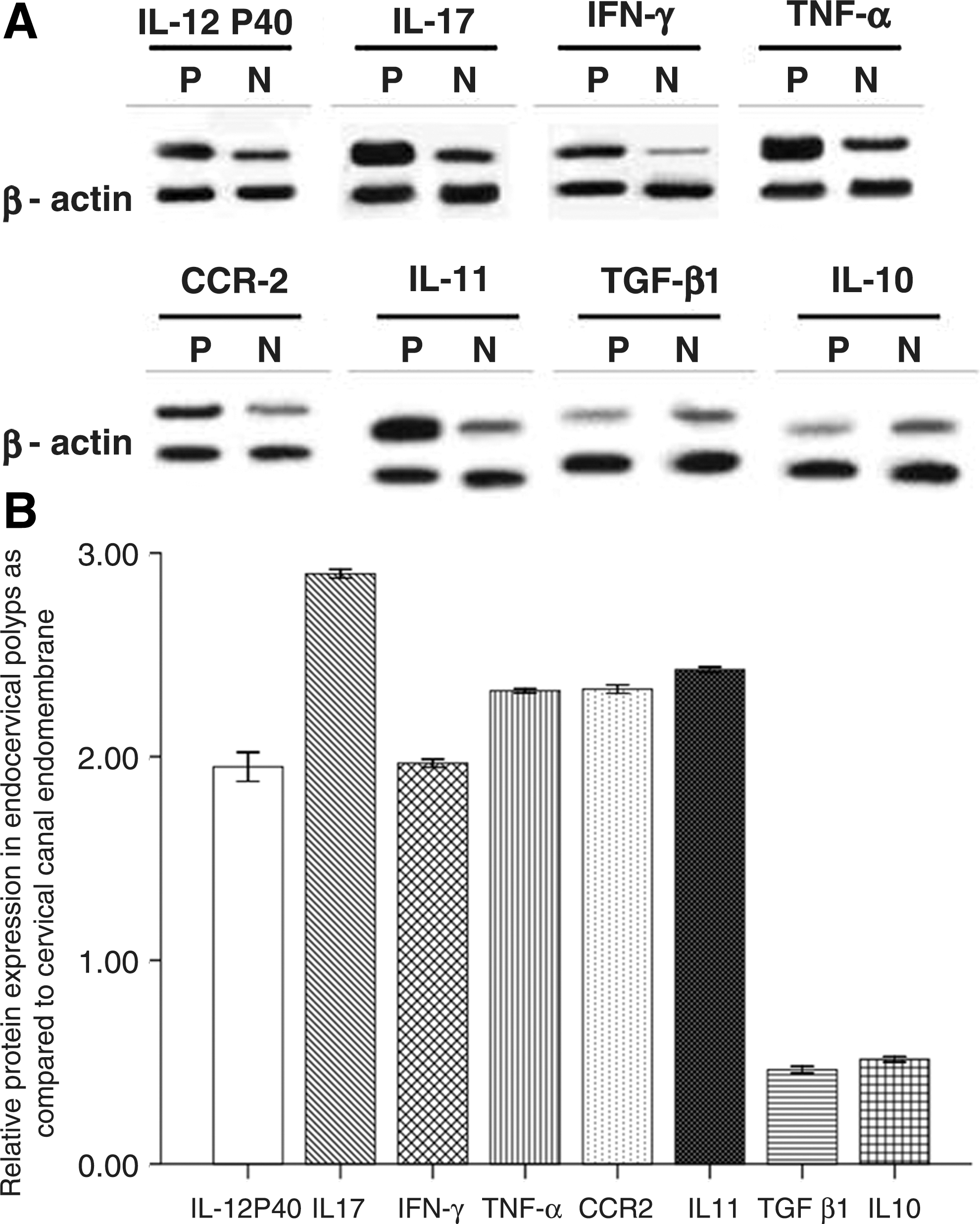
Relative protein expression of inflammatory response genes in endocervical polyps.
Immunohistochemical analysis
We next compared the expression of these 8 proteins in interstitial cells and acinar glands in endocervical polyps and endocervical canal membranes using immunohistochemical analysis (Figs. 4 and 5). In agreement with the microarray analysis and the Western blot analysis, the protein expression of the 6 upregulated genes and the 2 downregulated genes was significantly different in inflammatory cells of the endocervical polyps compared with the acini of the glands (P<0.001). Endocervical polyps had significantly more interstitial cells, and cells in the acinar glands expressed the proteins for the 6 upregulated genes (IL-12P40, IL-17, IFN-γ, TNF-α, CCR2, and IL-11) than the endocervical canal tissues did. Conversely, significantly fewer cells in endocervical polyps expressed TGF-β1 and IL-10 than those in endocervical canal membranes for both interstitial cells and the glands' acini.

Immunohistochemistry results for the expression of inflammatory proteins in endocervical canal membrane and polyps. Immunohistochemical staining was performed on samples from 50 endocervical polyps and 38 endocervical canal membrane tissues as described in the Materials and Methods section. For each sample and antibody combination, 5 microscopic fields were evaluated for 200 interstitial cells and 200 acinar gland cells.

The quantification results of immunohistochemistry results. Number of cells stained positive for indicated protein in the two different cell types. Cell counts were given as mean±SD. *P<0.001, significant difference between endocervical polyps and endocervical canal membrane.
Discussion
We found that 16 of 509 inflammation-associated genes were differentially regulated in endocervical polyps compared with control endocervical canal membrane tissues in our microarray analyses. These differentially expressed genes were mainly pro- or antiinflammatory factors and their receptors, adhesion molecules, leukocyte differential antigen, and inflammation signal transduction molecules.
Endocervical polyps are common and are usually clinically benign growths. Although the pathogenesis remains largely unknown, 74.1% of cervical polyp patients had some form of chronic inflammation, such as cervical erosion or cervical hypertrophy (Yi and others 2009). Cervical polyps are frequently accompanied with endometrial hyperplasia, indicating that hyperestrinism plays a significant etiologic role in endocervical polyp formation. Long-term stimulation due to chronic cervicitis can lead to mucosal hyperplasia, which is closely associated with cervical polyps (Stamatellos and others 2007). The increased numbers of activated macrophages and monocytes, eosinophils, neutrophils, and plasma cells observed in endocervical polyps compared with those in uterine cervical mucosa suggest that inflammatory processes play important roles in the pathophysiology of endocervical polyps (Perez Novo and others 2010).
Genes that were significantly upregulated in endocervical polyps were IL-12P40, IL-17, IL-11, IFN-γ, TNF-α, and CCR2. These cytokines were often found at higher levels in some nasal polyps (Molet and others 2003; Saitoh and others 2010; Ba and others 2011; Shen and others 2011), but not all nasal polyps (Wang and others 2006; Shi and others 2009; Ba and others 2011). IL-12P40 increases neutrophil accumulation (Ma and others 2000), which has been observed in some polyps (Ba and others 2011). IL-11 and IL-17 levels were also higher in nasal polyps (Molet and others 2003; Wang and others 2006; Liu and others 2011) and they may influence collagen deposition and remodeling processes (Molet and others 2003). Whether a chronic infection in the cervical canal membrane may have initiated the inflammation or contributed to its continuity requires further study. The signaling pathway of MCP-1/CCR-2 affects migration and transmigration of monocytes and macrophages (Li and others 2008), modulates Th1 and Th2 homeostasis through various mediators, and appears to affect vascularization (Charo and Taubman 2004). We tentatively propose that CCR2 may participate in mononuclear cell infiltration into cervical canals during cervical polyp formation. Because chemotactic factors and their receptors can promote inflammatory cell migration to the membranes of endocervical canals, it is feasible that these cells may participate in angiogenesis of the membranes of endocervical canals.
In follow-up experiments with Western blots and immunochemistry, proteins from the 8 examined genes (6 upregulated and 2 downregulated) were differentially expressed in endocervical polyps versus the normal cervical canal membranes, in agreement with the RNA results of the microarray analyses. Our DNA microarray results also showed that IL-1β, IL-2, IL-10, IL-15, IL-22, CCL-15, IFN-β1, TGF-β1, and CD86 levels were lower in endocervical polyps than normal endocervical canal tissues. Reduced antiinflammatory mediators in endocervical polyps may skew the balance between antiinflammatory and proinflammatory factors, which could increase the susceptibility of the cervix to infection and polyp generation. For example, low IL-10 levels may lengthen the duration of an inflammatory response to an infection. We hypothesize that increasing IL-10 expression could reduce the pathogenic conditions in cervical polyp patients and may decrease the incidence of cervical polyps in susceptible women.
Surprisingly, IL-6 was not differentially expressed in endocervical polyps in this study whereas IL-6 was upregulated in most or all endocervical polyps examined previously (Peters and others 2010). These data suggest that the development of endocervical polyps may involve different signaling pathways.
One limitation of our study was the use of pooled samples. While the pooled samples provided an analysis of the cytokine profile of an average endocervical polyp, differences in expression of cytokine- and inflammatory-related genes between samples would have been reduced. The distinct patient sources for cervical polyp tissues and normal endocervical canal membrane tissues avoided the possibility of cross contamination although a genotypic difference between the groups may potentially affect the expression of the aforementioned genes. Another potential confounding factor is that we did not take into consideration possible effects of menstrual cycle stage on gene expression.
In summary, we obtained the expression profiles for inflammation-associated genes in endocervical polyps using gene chip technology and identified 16 differentially expressed genes. However, we did not identify any genes that appeared to be unique to endocervical polyps. More detailed analyses of these genes, their protein products, and potential inducing factors will provide clues to the pathogenesis, inflammation therapy, and potential new targeted therapies for endocervical polyps.
Footnotes
Acknowledgment
This work was supported by the Special Fund for Science and Technology for Youth, Heilongjiang (QC07C87).
Author Disclosure Statement
No competing financial interests exist.