
Abstract
A hallmark of acute classical swine fever is the high interferon (IFN)-α levels found in the serum early after infection, followed by an inflammatory cytokine storm. Plasmacytoid dendritic cells (pDCs) represent the only known cell type that produces IFN-α upon classical swine fever virus (CSFV) infection in vitro. In primary target cells of the virus the viral protein Npro inhibits the induction of type I IFN via the degradation of IRF3. We hypothesized that the early systemic pDC-derived IFN-α response sensitizes immune cells for enhanced responsiveness and augment cytokine responses after CSFV infection through the upregulation of IRF7. Therefore, bone marrow-derived granulocyte macrophage-colony stimulating factor (GM-CSF)-induced DCs, were pretreated with IFN-β or conditioned medium from CSFV-activated enriched pDC, and expression of the pro-inflammatory cytokines interleukin (IL)-1β, IL-6, and IFN-α was assessed after infection with wild-type CSFV and with an Npro mutant [Npro(D136N)] unable to interact with IRF3 and IRF7. While type I IFN treatment sensitized the DCs for enhanced IFN and cytokine responses after stimulation with influenza virus, lipopolysaccharide or poly(I):poly(C), this was not observed for CSFV. In contrast, the Npro(D136N) mutant CSFV induced elevated IFN-α responses in type I IFN-pretreated GM-CSF DCs. These results indicate that CSFV has evolved to prevent type I IFN sensitization in infected cells through the action of the Npro.
Introduction
C
Nevertheless, the underlying mechanism that leads to the observed immunopathological events or the cellular sources of these cytokines are not clear. Particularly puzzling is the discrepancy between in vitro and in vivo observations. In vitro, primary target cells of CSFV such as macrophages, myeloid dendritic cells, and endothelial cells show only weak or no pro-inflammatory cytokine response and no type I IFN expression after CSFV infection (Knoetig and others 2002; Carrasco and others 2004; Ruggli and others 2005). Also, CSFV-infected monocyte- or bone marrow-derived DCs show no signs of maturation or activation in terms of MHC or CD80/86 up-regulation or cytokine expression in vitro (Carrasco and others 2004). The lack of innate immune system activation by CSFV in vitro is, at least in part, caused by the ability of Npro to prevent IRF3-mediated activation, through induction of proteosomal degradation of the transcription factor (Ruggli and others 2003; Bauhofer and others 2005; Bauhofer and others 2007). In apparent contradiction, during acute CSF high IFN-α, tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 levels are found in the serum and cytokine-expressing cells are found in lymphoid tissue (Sanchez-Cordon and others 2002; Summerfield and others 2006; Jamin and others 2008). These responses appear to relate to the virulence of the CSFV strain as well as to the severity of the disease and were suggested to contribute more to the disease development than to protection with virulent strains of CSFV (Ruggli and others 2009).
In vitro, the only cell type that was shown to produce IFN-α and TNF-α in response to CSFV infection are plasmacytoid dendritic cells (pDCs) (Balmelli and others 2005), a subset of DCs specialized in the production of type I IFN (Liu 2005). It was proposed that pDCs can respond to CSFV due to their unique constitutive expression of IRF7 and independence on IRF3 (McCullough and others 2009). However, the fact that pDCs are a very rare cell type questioned whether they are solely responsible for the massive production of IFN-α and inflammatory cytokines during acute CSF. For influenza virus infections a comparable discrepancy between nonresponsiveness of target cells in vitro and effective innate immune responses induced in vivo are observed. Nevertheless, while the NS1 of influenza virus efficiently blocks activation of conventional DCs, this can be overcome by exposure of the DCs to low doses of IFN-β prior to Influenza A virus (IAV) infection (Osterlund and others 2005; Phipps-Yonas and others 2008). This priming of DCs with type I IFN resulted in influenza virus-induced IFN-α and TNF-α secretion as well as DC activation, and is possibly the basis for a wider distribution of cell types involved in innate immune responses during infection.
Based on this evidence and on the fact that the early systemic IFN-α could sensitize non-pDC cells for enhanced IFN-α/β and inflammatory cytokine production upon CSFV infection, we hypothesized that similar mechanisms for innate immune cell activation could play a role during CSFV infections in vivo. Considering that pDCs would provide a source of type I IFN, we addressed whether CSFV-induced cytokines and type I IFN in conventional dendritic cells or macrophages could be promoted by type I IFN priming through the induction of IRF7 expression.
Material and Methods
Generation of bone marrow hematopoietic cells-derived DCs using granulocyte macrophage-colony stimulating factor
Generation of granulocyte macrophage-colony stimulating factor (GM-CSF)-induced, bone marrow hematopoietic cells (BMHC)-derived porcine DCs was preformed as described previously (Carrasco and others 2001). Briefly, BMHCs collected from a sternum were seeded in T150 flasks at a density of 1×106 cells/mL in 40 mL RBMI 1,640 medium (Invitrogen) supplemented with 10% fetal bovine serum (FBS; Biowest), 100 U/mL penicillin, 100 μg/mL streptomycin (Invitrogen), GlutaMAX™-I (Invitrogen), and 100 U/mL recombinant porcine GM-CSF (Summerfield and others 2003). Cells were fed every 3 to 4 days with GM-CSF. After 14 days of culture, the nonadherent cells, the majority of them displaying DC morphology, were collected.
Isolation and culture of enriched pDCs
Peripheral blood mononuclear cells (PBMCs) were isolated from the blood of specific pathogen-free pigs using Ficoll-Paque (1.077g/L, Amersham Pharmacia Biotech AG) density-gradient centrifugation. pDCs were enriched by CD172a+ sorting using the Miltenyi magnetic-activated cell sorting separation system with LD columns (Miltenyi Biotec GmBH, Bergish Gladbach, Germany) and the anti-CD172a mAb clone 74-22-15A as described previously (Guzylack-Piriou and others 2004). CD172a+ PBMC, composed of monocytes, conventional blood DC, and pDCs (Summerfield and McCullough 2009), were plated at 4×105 cells/well in a U-bottom 96-well plates in Dulbecco's modified Eagle's medium+GlutaMAX I (DMEM; Invitrogen) supplemented with FBS and 0.02 mM β-mercaptoethanol (Invitrogen).
Viruses
CSFV strain vA187-1 wild-type (WT) (Ruggli and others 1996) and the Npro mutant CSFV vA187-D136N [Npro(D136N)] carrying a D-to-N substitution at position 136 of the polyprotein (Ruggli and others 2009) were grown in the porcine aortic endothelial cell line PEDSV.15 (kindly obtained from Dr. Jörg Seebach, Division of Immunology and Allergology, Department of Internal Medicine, University Hospital Geneva, Switzerland). Virus titers were determined on PK-15 cells by endpoint dilution and expressed as 50% tissue culture infectious doses (TCID50)/mL. The IAV NIBRG23 carrying the HA (without polybasic cleavage site) and NA from H5N1 A/turkey/Turkey/1/2005 and the remaining genes from H1N1 A/PR/8/34 (kindly obtained from Jim Robertson, National Institute for Biological Standards and Control, UK) was rescued by reverse genetics. The virus was grown in embryonated 10-day-old chicken eggs as described (Ocana-Macchi and others 2009). The IAV titers were determined by end-point titration on MDCK cells in the presence of 1 μg/mL L-1-tosylamid-2-phenylethyl chloromethyl ketone trypsin (Sigma-Alderich). About 72 h postinfection (p.i.) the cells were washed and stained with crystal violet. The TCID50/mL was calculated according to the Reed-Muench formula (WHO Manual on Animal Influenza Diagnosis and Surveillance).
Infection and treatment of DCs
In some cultures, GM-CSF DCs were pretreated with different doses of recombinant porcine IFN-β (produced in house as described) (Husser and others 2011) in suspension at a density of 2×106 cells/mL in RPMI medium without FBS. Thereafter, the IFN-β-containing medium was removed, fresh medium was added, and the cells were seeded in 48-well plates at a density of 5×105 cells/well in 500 μL. For stimulation the cells were infected at the indicated multiplicity of infection (MOI) with WT or Npro(D136N) mutant CSFV or with IAV, or treated with 10 μg/mL polyinosinic polycytidylic acid (pIC) (Sigma Chemicals), 10 μg/mL LPS (Sigma), or 10μg/mL CpG DNA oligonucleotide D32 sequence (Guzylack-Piriou and others 2004) (Biosource). After 2 h of virus infection or stimulation, FBS was added to a final concentration of 10% (v/v) if not indicated otherwise.
Detection of intracellular viral proteins by flow cytometry
For labeling of intracellular viral proteins, cells were fixed and permeamilized with Fix & Perm permeabilization kit (ADG Bio Research GmbH) according to the manufacturer's instructions. The CSFV protein NS3 was stained with the monoclonal antibody (mAb) C16 (kindly provided by Dr. Irene Greiser-Wilke, Veterinary School Hannover, Germany), and the NP protein of NIBRG23 was detected with the mAb clone HB65 (ATCC). Binding of the primary antibodies were revealed using phycoerythrin-conjugated anti-mouse IgG F(ab’) 2 fragments (Dako).
mRNA isolation and real-time reverse transcription–polymerase chain reaction
Samples of 5×105 DCs treated as indicated were pelleted, and total RNA was isolated using TRIZOL (Invitrogen) according to manufacturer's protocol and treated with RNase-free DNase I (Ambion). The real-time reverse transcription–polymerase chain reaction (real-time RT PCR) was performed with the SuperScript™ III Platinum®One-Step qRT-PCR System (Invitrogen) using the ABI PRISM 7,700 sequence Detector system (Applied Biosystems). The relative expression of IRF3 and IRF7 mRNA was calculated by the ΔCt method and the amount of IRF-3 or IRF-7 mRNA relative to 18S mRNA was expressed as 2−Ct (Livak and Schmittgen 2001). The following primers and probes were used: 18S forward primer 5′-CGC CGC TAG AGG TGA AAT TC-3′; 18S reverse primer 5′-GGC AAA TGC TTT CGC TCT G-3′; 18S probe 5′-TGG ACC GGC GCA AGA CGG A-3′; IRF-3 forward primer 5′-GGACCTGCACATTTCCAACAG-3′; IRF-3 reverse primer 5′-AGCGGCCTAGAAATCCATGTC-3′; IRF-3 probe 5′-CCTCACCTCCGACCAGTACAAGGCCT-3′; IRF-7 forward primer 5′-CTGCGATGGCTGGATGAA-3′; IRF-7 reverse primer 5′-TAAAGATGCGCGAGTCGGA-3′; IRF-7 probe 5′-CCGCGTGCCCTGGAAGCACTT-3′ (Alves and others 2009).
Detection of cytokines in the cell culture supernatant
Secreted IFN-α was quantified by enzyme-linked immunosorbent assay (ELISA) using the mAbs K9 and F17 (kindly provided by Dr. B. Charley, INRA, Jouy-en-Josas, France) as described previously (Guzylack-Piriou and others 2006). IL-6 and IL-1β in the cell culture supernatants were quantified using the porcine IL-6 DuoSet® ELISA and the porcine IL-1β/IL-1F2 DuoSet® ELISA, respectively (both from R&D Systems).
Statistical analysis
P values were calculated by unpaired t-test in SigmaPlot® for normally distributed data. When normality test failed, a Mann–Whitney Rank Sum test was performed. Differences were considered significant at a P value of <0.05. Error bars represent the standard deviations of triplicates in one experiment and were calculated with Excel (Microsoft).
Results
Pretreatment of GM-CSF DCs with various doses of IFN-β does not lead to enhanced IFN-α in response to CSFV
We first explored the dose and time dependency of IRF7 and IRF3 induction after IFN-β treatment of GM-CSF DCs. Western blot was not sensitive enough to detect endogenous IRF7 protein in porcine GM-CSF DCs (data not shown). Therefore we employed real-time RT-PCR to assess IRF7 and IRF3 mRNA induction in GM-CSF DCs that were pretreated with 50, 100 and 1,000 U/mL IFN-β for 2, 6 and 16 h (Fig. 1A). Two hours after IFN-β treatment IRF7 mRNA was strongly up-regulated at all 3 applied IFN-β concentrations, and the expression increased only slightly with longer incubation or higher IFN-β doses. As expected, IRF3 mRNA levels remained unchanged.
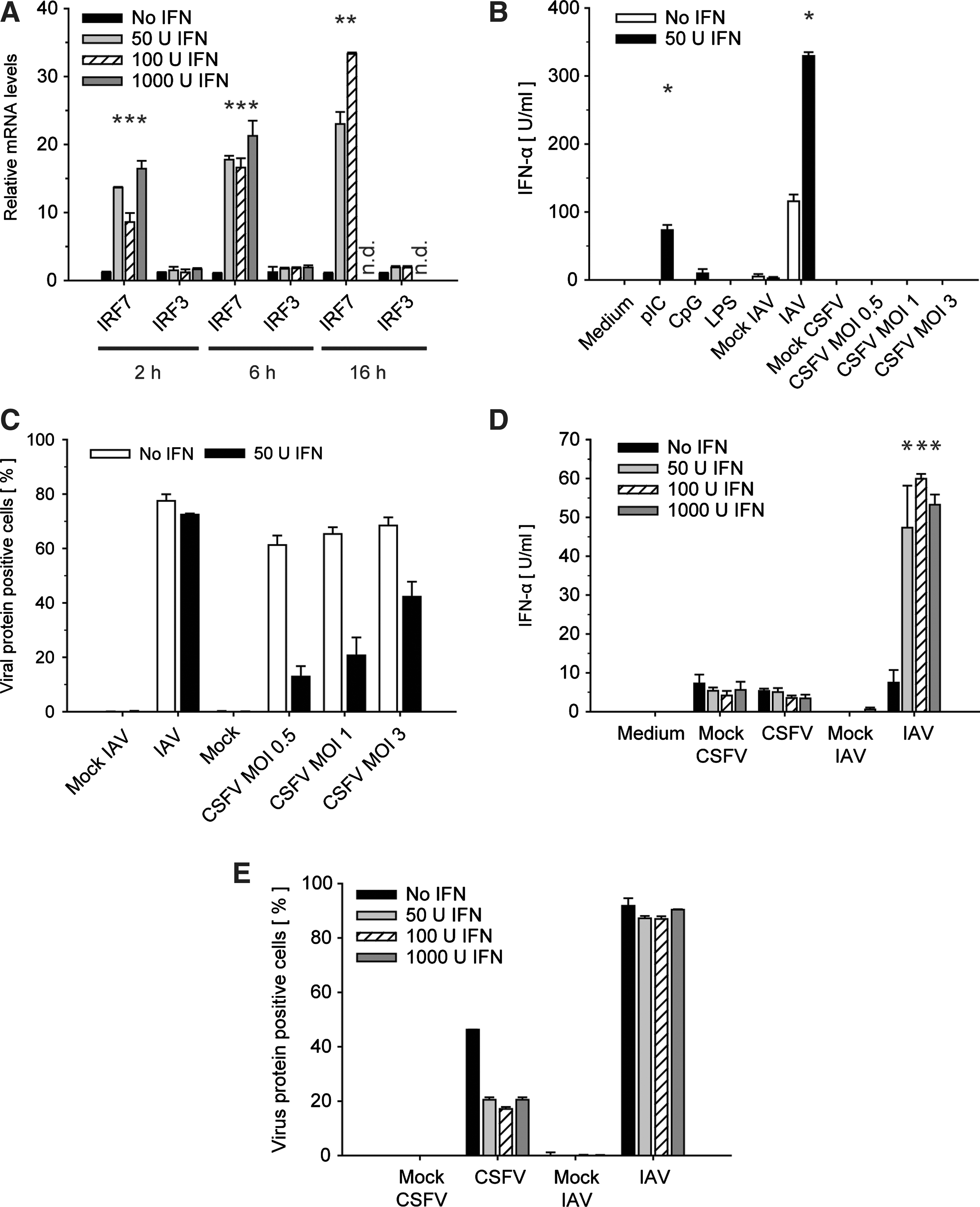
Effect of pretreatment with IFN-β on IFR7 and IRF3 mRNA up-regulation, virus-induced IFN-α response, and viral replication. GM-CSF DCs were left untreated or stimulated with 50, 100, or 1,000 U/mL IFN-β for 2 h if not otherwise indicated.
After having shown that IRF7, but not IRF3, was strongly induced in IFN-β-treated GM-CSF DCs, we examined the virus-induced IFN-α responses in GM-CSF DCs pretreated with 50 U/mL IFN-β for 2 h. IAV for which it was reported that type I IFN priming enhances virus-induced cytokine expression (Phipps-Yonas and others 2008) served as a positive control. CpG treatment was employed to confirm the absence of pDCs that are the only porcine cells known to respond to both CpG and CSFV by IFN-α secretion (Summerfield and McCullough 2009). While unprimed cells secreted no or low levels of IFN-α after IAV infection, IFN-β pretreatment for 2 h rendered the GM-CSF DCs clearly responsive to IAV in terms of IFN-α secretion (Fig. 1B). Furthermore, IFN-β pretreatment primed GM-CSF DCs for enhanced IFN-α expression after stimulation with pIC. In contrast, CSFV WT did not trigger any IFN-α response in GM-CSF DCs regardless of the viral dose employed and whether the cells were pretreated with IFN-β or not (Fig. 1B).
In order to determine a potential impact of the virus sensitivity to type I IFN, we monitored in parallel the capacities of CSFV and IAV to infect IFN-β-primed GM-CSF DCs as compared to untreated cells. While IAV infection was not affected by preexposure to IFN-β, the infection levels of CSFV were clearly reduced (Fig. 1C).
Next we increased the IFN-β doses to examine whether a higher concentration of IFN-β would trigger a CSFV-induced IFN-α response in GM-CSF DCs. Relating to the IRF7 levels shown in Fig. 1A, all doses of IFN-β had a similar biological effect on IAV-induced IFN-α expression. Importantly, priming with up to 1,000 U/mL IFN-β for 2 h did not lead to IFN-α secretion upon CSFV infection (Fig. 1D). Interestingly, also the antiviral effect measured in terms of the ability to prevent CSFV NS3 or IAV NP expression was similar with all doses. While CSFV infection was reduced by ∼60%, IAV infection was remarkably resistant to IFN-β even with a dose of 1,000 U IFN-β/mL (Fig. 1E).
Impact of IFN-β priming duration
Priming with different doses of IFN-β for 2 h did not result in augmented IFN-α response to CSFV infection. Therefore, we continued our investigations by modulating the duration of exposure to IFN-β. GM-CSF DCs were cultured in the presence of IFN-β for 6 or 16 h, and subsequently stimulated with pIC or CSFV. Significantly enhanced IFN-α expression was still observed when GM-CSF DCs were primed for 6 h but not for 16 h before stimulation with pIC (Fig. 2 A). Also, under these conditions CSFV was not able to induce consistent and significant amounts of IFN-α. It is important to note that we were also unable to prime other cell types such as PK-15 cells, monocyte-derived DCs, and monocyte-derived macrophages for type IFN responses after infection with CSFV (data not shown).
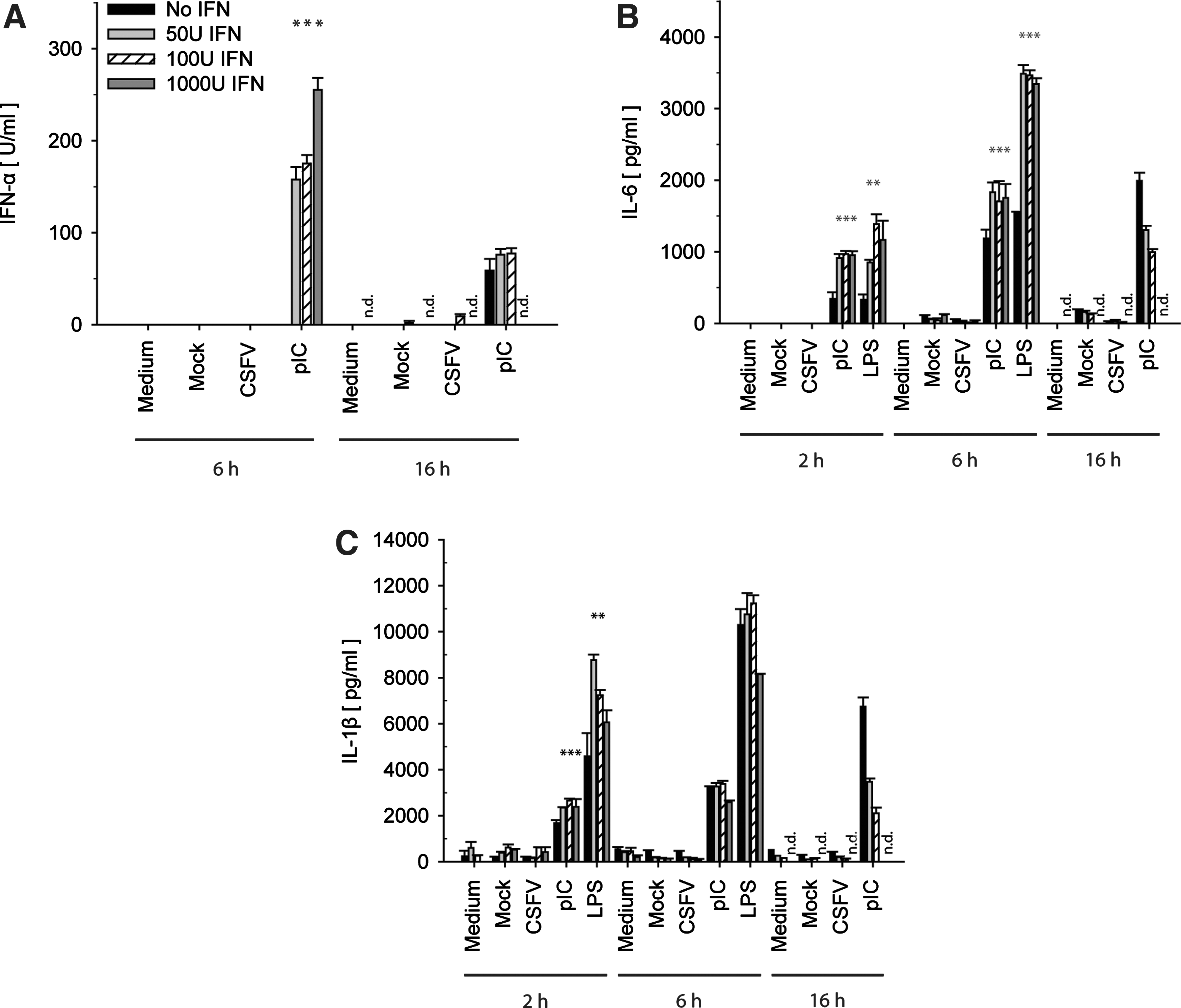
IFN-β dose- and time-dependent IFN-α, IL-6, and IL-1β responses after virus infection or PRR stimulation. GM-CSF DCs were incubated with 50, 100, or 1,000 U/mL of IFN-β for 2, 6, and 16 h. Thereafter, the cells were washed and stimulated with LPS (10 μg/mL), pIC (10 μg/mL), CSFV (MOI 3 TCID50/cell), or IAV (MOI of 0.5 TCID50/cell). The mock controls were as described in Figure 1. After 24 h, IFN-α
IFN-β priming and proinflammatory cytokines
In acute CSF, besides IFN-α, also elevated levels of pro-inflammatory cytokines are detected in the serum of CSFV-infected pigs (Sanchez-Cordon and others 2002; Summerfield and others 2006; Jamin and others 2008). Therefore, we analyzed whether IFN-β pretreatment of GM-CSF DCs followed by CSFV infection would lead to enhanced expression of the IL-1β and IL-6 cytokines, as models for the pro-inflammatory response. Neither IL-1β nor IL-6 secretion was enhanced after exposure of GM-CSF DCs to various concentrations of IFN-β (50, 100, and 1,000 U/mL) for different time periods (2, 6, and 16 h) before CSFV infection (Fig. 2B,C). Yet, a priming effect of both cytokines was observed with LPS and pIC treatment confirming that GM-CSF DCs are able to display enhanced pro-inflammatory cytokine responses after IFN-β treatment. Compared with unprimed GM-CSF DCs, LPS- and pIC-induced IL-6 responses were clearly augmented, in particular with short priming periods (Fig. 2B). The strongest enhancement of IL-1β expression was observed after LPS treatment and only when priming was limited to 2 h of pretreatment (Fig. 2C). IAV did induce neither IL-6 nor IL-1β (data not shown).
CSFV-induced cytokine responses in GM-CSF DCs pretreated with conditioned medium derived from CSFV-stimulated pDCs
According to our hypothesis, pDC-derived IFN-α would act on cDCs to prime enhanced cytokine responses in vivo. Yet, in our in vitro studies we employed recombinant IFN-β to mimic a type I IFN-containing environment. In order to investigate potential effects of other type I IFN or other cytokines produced by pDC could have on GM-CSF DCs, we compared cytokine production of GM-CSF DCs that had been pretreated with either recombinant IFN-β or with conditioned medium derived from CSFV-activated enriched pDCs. In both cases, primed DCs responded with significantly higher IFN-α (Fig. 3A) or IL-6 expression (Fig. 3B) compared to unprimed DC when stimulated with IAV or LPS, respectively. In contrast, neither the conditioned medium containing IFN-α derived from CD172a+ PBMC composed of monocytes, conventional blood DC, and pDC nor recombinant IFN-β was able to prime the GM-CSF DC for CSFV-induced activation in terms of type I IFN or IL-6 responses.

Priming effect of conditioned medium derived from CSFV-infected enriched pDCs on IFN-α and IL-6 responses in GM-CSF DCs CD172a+ peripheral blood mononuclear cells (enriched pDCs) were mock (CSFV Mock) treated or infected with wild-type CSFV (CSFV WT) at an MOI of 2 TCID50/cell and 24 h postinfection (p.i.) cell culture supernatant were harvested and treated with 75 mJ UV to inactivate the virus. GM-CSF DCs were then incubated for 3 h with either pDC-derived medium diluted 1:2, 1:4, or 1:40, or with medium conditioned with 50 U/mL recombinant IFN-β, which was added after (IFN-β) or before UV treatment (UV IFN-β) or with medium only (No IFN). Thereafter, GM-CSF DCs were stimulated with LPS (10μg/mL), CSFV (MOI of 2 TCID50/cell), or IAV (MOI of 0,5 TCID50/cell), and 24 h p.i. IFN-α
IFN-α responses to Npro(D136N) mutant CSFV can be primed by IFN-β
A recent report published by our laboratory showed that the CSFV-encoded IFN antagonist Npro interacts not only with IRF3 but also with IRF7 (Fiebach and others 2011). Therefore, we asked whether the inability of CSFV to induce type I IFN or cytokine responses in IFN-β-primed GM-CSF DCs expressing IRF7 was related to the function of Npro. In order to address this, we employed the Npro(D136N) mutant CSFV that is unable to interact with IRF3 and IRF7 (Ruggli and others 2009; Fiebach and others 2011). Interestingly, the Npro(D136N) mutant virus was more sensitive to IFN-β pretreatment than the WT virus in terms of its ability to infect GM-CSF DCs, irrespectively of the MOI employed (Fig. 4A). Importantly, even though IFN-β priming reduced the infection of the Npro(D136N) mutant virus by at least 70%, the IFN-α levels were comparable in primed and nonprimed cells (Fig. 4B). To take the reduced infection caused by the IFN-β priming into account, we calculated the IFN-α levels per NS3-positive cell after infection with the Npro(D136N) mutant virus (Fig. 4C). The results show clearly that IFN-β priming promotes an elevated IFN-α secretion in NS3+ GM-CSF DCs only when the cells are infected with a CSFV mutant devoid of IRF7 binding. This was also confirmed by simultaneous infection and IFN-β treatment (Fig. 4D). Under these conditions, IFN-β enhanced also the absolute IFN-α level induced by the Npro(D136N) mutant virus. Again, the WT CSFV remained unable to induce IFN-α. Also, the simultaneous addition of IFN-β with IAV virus primed the GM-CSF DCs to respond by production of IFN-α, contrasting with the pIC-induced IFN-α secretion that required IFN-β pretreatment. Under these priming conditions, none of the viruses elicited an IL-6 or IL-1β response (data not shown).

Role of Npro in viral escape from the IFN-β-mediated IFN-α amplification loop. In
Discussion
In this study we examined IFN-α and cytokine responses of GM-CSF DCs upon CSFV infection in the context of a type I IFN containing environment. Type I IFN has broad antiviral and immunomodulatory effects. On one hand, it has beneficial effects in viral infections restricting viral dissemination and promoting adaptive antiviral effector immune responses. On the other hand, it is also known for promoting immunopathological events when released at high levels during longer periods (Theofilopoulos and others 2005). Accordingly, during CSFV infection a strong and long-lasting IFN-α response is detectable in the serum of infected pigs, and this has been suggested to relate to disease severity rather than to protective immune responses (Summerfield and others 2006; Summerfield 2011). Considering that CSFV efficiently prevents type I IFN induction in all analyzed target cells through the induction of viral Npro-mediated proteasomal degradation of IRF3, the question arose about the cellular source of the systemic type I IFN found in diseased pigs. So far pDCs are the only cell type known to respond to WT CSFV infection by IFN-α production in vivo and in vitro. For IFN-α induction this specialized cell type relies on constitutively expressed IRF7, a unique property of pDCs. In contrast, non-pDCs only employ IRF7 after its induction during the IFN-dependent autocrine-paracrine amplification loop (Marie and others 1998). Notably, IRF7 is the most potent transcription factor for the type I IFN system (Sato and others 2000). These facts led us to the sensitization hypothesis proposing that pDC-derived IFN-α would promote systemic IRF7 expression and enable non-pDCs to bypass the Npro-mediated IRF3 inhibition. This would result in IFN-α and/or pro-inflammatory cytokine expression in a wide range of cells. In principle, this scenario may represent a key event contributing to the cytokine storm observed in the acute phase of the disease and would also have been supported by the fact that type I IFN is an early cytokine detectable in the serum of pigs suffering from acute CSF. However, despite the increased expression of IRF7 transcripts after IFN-β exposure, preincubation of GM-CSF DCs with various concentrations of IFN-β or condition medium from CSFV-activated enriched pDC for different time periods did not result in enhanced IFN-α, Il-6, or IL-1β expression in response to CSFV WT infection. This was not due to the culture conditions, since enhanced IFN-α responses were found upon IAV infections in IFN-β pre-exposed GM-CSF DCs.
In contrast to the responses to WT CSFV, those to CSFV with a functionally defective Npro unable to interact with IRF3 and IRF7 (Fiebach and others 2011) were enhanced by IFN-β priming. This was observed either by calculating IFN-α units per NS3+ GM-CSF DC or when CSFV and IFN-β were added simultaneously. In this context, it was observed that CSFV was clearly more sensitive to IFN-β pretreatment when compared to IAV that showed a remarkable resistance to IFN-β, even at high concentrations. In addition, the Npro(D136N) mutant was even more sensitive to IFN-β pretreatment when compared to the parent virus. The reason for this enhanced sensitivity is unknown, but it could possibly relate to the antiviral activity induced by the mutant but not by the WT. Alternatively, this observation may point toward an interference of Npro with the type I IFN response pathway. We also observed lower percentages of NS3-expressing cells in BMHC-derived Flt3L-induced DCs, monocyte-derived DCs, and macrophages when they were treated with various doses of IFN-β before CSFV infection (data not shown). These results relate to the observation that pestiviruses are sensitive to the action of type I IFN only when the IFN is added before infection but has little or no antiviral effect is found when added after infection (Schweizer and others 2006; Suter and others 2011).
The present work emphasizes the important role of Npro in blocking innate immune responses. This viral protein not only prevents IRF3-dependent IFN responses but also acts on the autocrine–paracrine amplification loop most probably by preventing also IRF7-dependent IFN responses. This conclusion is based on the specific and potent interaction of Npro with IRF7, although the direct mechanism and consequences of this interaction remain to be elucidated (Fiebach and others 2011). Our conclusion is also in line with the observation that in pDCs, which are known to respond in an IRF3-independent, but IRF7-dependent manner (Honda and others 2005), WT CSFV infection leads to reduced IFN-α responses when compared to infections with Npro(D136N) mutant virus (Fiebach and others 2011). For the immunopathogenesis of CSF, we conclude that the aberrant type I IFN and inflammatory cytokines responses observed in vivo cannot be explained by type I IFN-induced sensitization of immune cells, as seen in other viral infections such as IAV (Osterlund and others 2005; Phipps-Yonas and others 2008). We currently have several explanations for the discrepancy of in vitro and in vivo results. First, the high tropism of CSFV for macrophages and DCs and its accumulation in lymphoid tissue results in overall high type I IFN and cytokine responses although at the cellular level the responses are low. Second, in vivo DCs and macrophages are exposed to a more complex (cytokine) environment resulting in a strong CSFV-induced activation. Despite this still puzzling situation, our data demonstrate that certain viruses have evolved that inhibit not only primary type I IFN responses mediated by IRF3 but also the IRF7-amplified secondary IFN responses in non-pDCs.
Footnotes
Acknowledgments
We are very grateful to Daniel Brechbühl for providing the sterna.
This project was supported by the Swiss National Science Foundation (project 310000-116800).
Author Disclosure Statement
No competing financial interests exist.