
Abstract
The interferon (IFN)-inducing capacity of influenza virus plays a significant role in the efficacy of candidate live attenuated influenza vaccines (LAIVs). IFN is induced by a subpopulation of noninfectious biologically active particles (niBAPs) that can be defined and quantified as IFN-inducing particles (IFPs). When chicken embryonic cells were infected with increasing multiplicities of IFP (m
Introduction
Assays developed much earlier for scoring biological activities other than infectivity in populations of different viruses (Marcus 1959, 1982, 1986; Marcus and Sekellick 1985) were applied recently to influenza virus. They identified and quantified heretofore unrecognized subpopulations of noninfectious virus that were, nonetheless, biologically active as scored by well-defined phenotypes: (i) cell-killing particles (Ngunjiri and others 2008); (ii) IFN-inducing particles (IFPs) (Marcus and others 2005); and (iii) interfering induction-suppressing particles (ISPs) (Marcus and others 2005). Each of these influenza virus subpopulations is subsumed in the population measured as hemagglutinating particles. Collectively, all but plaque-forming particles and the hemagglutinating particles without measurable biological activity are considered noninfectious biologically active particles (niBAPs).
Because of the emerging role of the IFN system in influenza virus pathogenesis and the development of LAIVs, this study focuses on the IFN-inducing capacity of influenza virus populations. Analysis of IFN-induction dose (multiplicity)–response (IFN yield) curves (Marcus 1986; Marcus and others 2005, 2010) generated by the IFP subpopulation in influenza virus populations allows (i) determination of the maximum (quantum) yield that IFP can induce in a given cell system; (ii) quantification of the number of IFP in the virus population; and (iii) calculation of the efficiency of the IFP in terms of inducing IFN (Marcus and others 2005, 2010). The most commonly observed IFN-induction dose–response curve fits well a theoretical curve based on a Poisson distribution of IFP among the cell population in which every cell that receives ≥1 IFP produces a full yield of IFN. The maximum plateau (quantum) yield of IFN is reached when all of the cells have received 1 or more IFP. In the course of determining the capacity of influenza virus populations to induce IFN, we noted that some stocks of influenza virus consistently generated IFN dose–response curves that produced less IFN than expected at higher m
Materials and Methods
Viruses
Both rgA/TK/OR/71-SEPRL (H7N3) and its variant rgA/TK/OR/71-delNS1[1–124] (H7N3) were generated via a reverse genetics system (Wang and others 2008) and kindly provided by Chang-Won Lee (Ohio State University, Wooster, OH). A deletion of 10 nucleotides in the NS gene of the rgA/TK/OR/71-delNS1[1–124] virus produces a truncated NS1 product lacking the C-terminal effector domain (Wang and others 2008). The 2 viruses are otherwise isogenic for the remaining 7 gene segments. The rgA/TK/OR/71-delNS1[1–124] virus displays a propensity to produce IFP, while the virus with the intact NS gene produced high titers of ISP. Virus stocks were prepared in 10-day-old embryonated chicken eggs from specific-pathogen-free flocks (Charles River SPAFAS, Storrs, CT) by infecting with seed virus at near end-point dilution and incubating for 48 h at 34°C. Allantoic fluid containing virus was harvested from the eggs and stored at −80°C. The rgA/TK/OR/71-SEPRL (H7N3) virus contained a plaque-forming particle titer of 3.2×108/mL and an ISP titer of 4.0×109/mL. Due to the high ISP content, the IFP titer of this preparation could not be measured and was presumed negligible. The rgA/TK/OR/71-delNS1[1–124] (H7N3) virus possessed a plaque-forming particle titer of 2.0×108/mL and an IFP titer equal to 4.3×109/mL. The ISP titer of the rgA/TK/OR/71-delNS1[1–124] is below the limits of detection of our ISP assay (≤3.3×107/mL).
Cells and media
Monolayers of primary chicken embryo cells (CECs) were prepared from cell suspensions and plated at a final density of 1.0×107 CEC per 50 mm culture dish. Cell suspensions were prepared from 9-day-old embryos. The cell monolayers were maintained in NCI medium plus 6% calf serum and incubated undisturbed at 38.5°C (Sekellick and others 1986).
IFN-induction and induction-suppression assays
Monolayers of CEC were incubated for 9 days without a medium change (developmentally aged) for IFN inductions (Sekellick and Marcus 1986), and when infected with IFP they produced copious amounts of IFN. Briefly, a series of cell monolayers was infected with increasing numbers of virus particles, and the amount of IFN produced was collected at peak yields (24 h postinfection) (Sekellick and Marcus 1986; Marcus and others 2005). Biologically active, acid-stable IFN was quantified via a cytopathic effect-inhibition assay (Sekellick and Marcus 1986). IFP were calculated from the resulting dose–response curves as described in the text.
ISP are measured via a similar assay in which monolayers of 9-day aged CEC are simultaneously exposed to a known IFN-inducing virus, UV-irradiated avian reovirus (7,500 ergs/mm2) (Winship and Marcus 1980), and increasing amounts of the suspected IFN induction-suppressing virus. This procedure has been described previously (Marcus and Sekellick 1985; Marcus and others 2005). Briefly, a control trial in which every cell in a CEC monolayer is infected by only the inducing virus (m
IFP
=5.0) is used to establish a maximum IFN yield. Simultaneous exposure to increasing multiplicities of the ISP-containing preparation results in a decline in IFN yield from that maximum. According to the Poisson distribution, the volume of ISP-containing virus that reduces the maximum IFN yield to 37% contains on average m
Results
Theoretical IFN induction dose (multiplicity)–response (IFN yield) curves
Figure 1 displays families of theoretical dose–response curves that IFP might generate in cell populations. They fall into 2 general types, termed (A) r≥1, r≥2, r≥3,…, and (B) r=1, r=2, r=3,…, where the class of cells that produce IFN is equivalent to the fraction of cells infected with r IFP at a given m
P
(r) is the probability of the cells in a given population attaching r IFP when the average multiplicity of infection is m
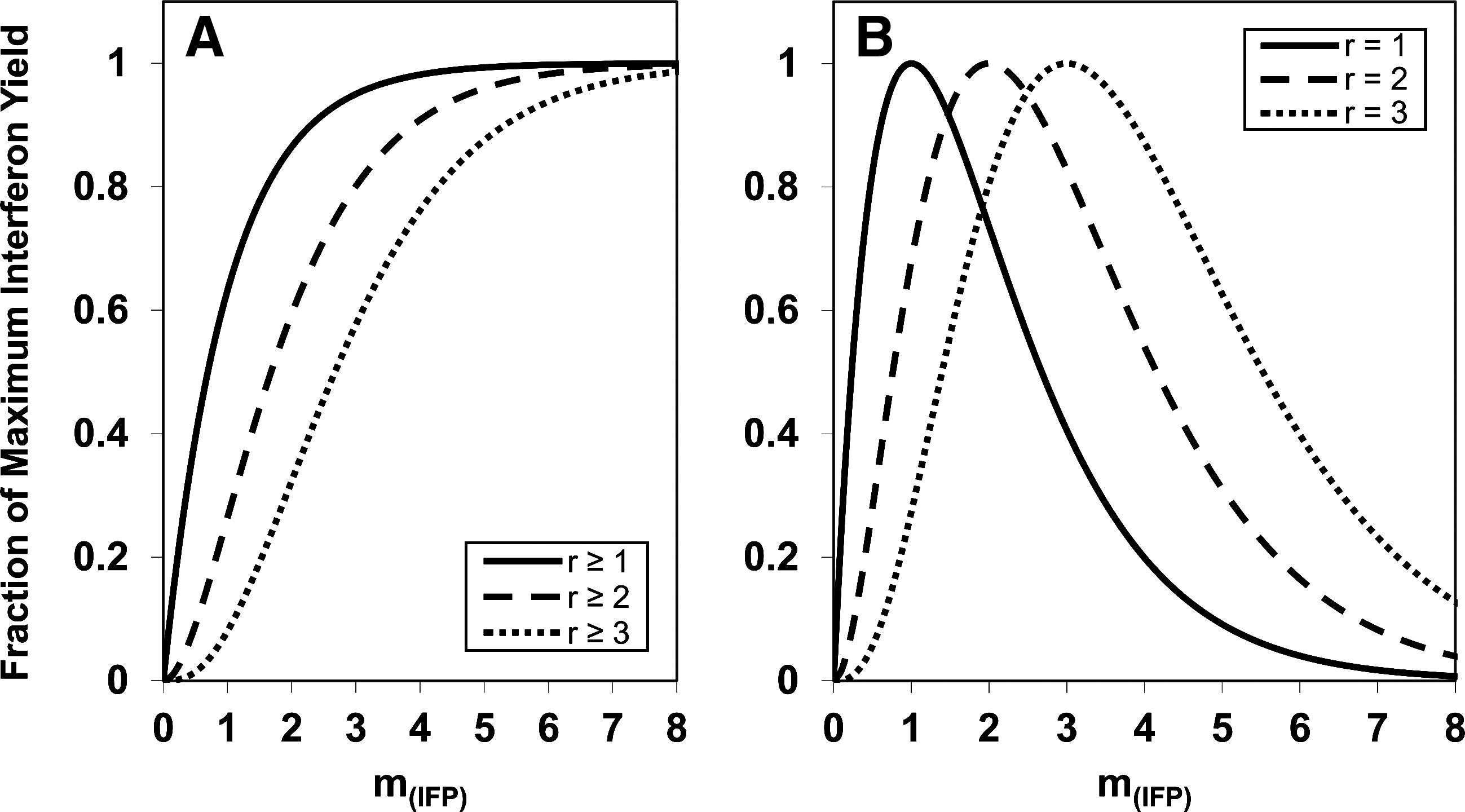
IFN-induction dose–response curves.
Figure 1B shows a family of theoretical IFN-induction curves represented by types r=1, r=2, and r=3. These curves are characterized by a rapid decrease in the yield of IFN after the quantum yield is reached at m
Observed versus theoretical IFN-induction dose–response curves: an anomaly
Influenza virus stocks commonly generate IFN-induction curves that closely approximate type r≥1 (Marcus and others 2010) when developmentally aged primary chicken embryonic cells are hosts (Carver and Marcus 1967; Marcus and Sekellick 1977). Problematically, some stocks consistently display a decrease in IFN yield of varying magnitudes once an apparent plateau is reached, and therefore do not conform precisely to any of the theoretical curves described in Fig. 1. For example, in Fig. 2 a stock of rgA/TK/OR/71-delNS1[1–124] virus does not maintain a plateau yield of IFN at higher m

IFN-induction dose–response curves. Data points represent an average of 3 IFN-induction experiments using rgA/TK/OR/71-delNS1[1–124] virus. Cell monolayers of developmentally aged primary chicken embryonic cells were infected with increasing multiplicities of IFP as indicated on the abscissa, incubated for 24 h at 40.5°C, and the supernatant harvested and assayed for IFN (Sekellick and Marcus 1986). The IFN-induction dose–response curve as shown (dotted line) is not adequately described by the traditional theoretical type r≥1 dose–response curve (solid line) because of the marked decline in IFN yield after the maximum (plateau) is reached. Error bars represent the mean±SD. IFP, IFN-inducing particle; SD, standard deviation.
Hypothesis: ISPs are responsible for the less than expected yields of IFN observed at high m IFP
Insight into the discrepancy between expected and observed yields of IFN noted in Fig. 2 was gained by considering the presence of an ISP subpopulation in stocks of influenza with otherwise high titers of IFP. In this context, the suppressing activity of influenza virus ISP is dominant to IFN induction by IFP in coinfected cells (Marcus and others 2005), as is the case for ISP found and quantified in several families of viruses (Marcus and Sekellick 1985). We postulated that the observed decline in IFN yield at high m
The veracity of this postulate was tested based on 3 assumptions: (i) the virus population consisted of a mixture of discrete particles, IFP and ISP; (ii) infection of a cell by 1 or more IFP resulted in the production of a full yield of IFN, and; (iii) coinfection of cells by 1 or more ISP suppressed IFN induction completely in a cell otherwise programmed to produce it. A series of theoretical curves based on these assumptions were derived from the following equation:
where r
(
This calculation is different from the previous, which assumed that only IFP were present. This model proposes that influenza virus stocks may contain 2 discrete subpopulations of particles: one that induces IFN and one that suppresses its induction, and that the phenotype of the latter is dominant over the former (Marcus and Sekellick 1985; Marcus and others 2005). It is important to note that the probability of a cell receiving 1 or more IFP and no ISP is dependent upon the relative proportion of IFP to ISP in the preparation. The equation as written does not assume that the relative ratios of IFP and ISP are equal, but varies with the values of m

Experimental reconstruction of hybrid IFP:ISP dose–response curves. The solid line is theoretical and represents the yield of IFN expected from a pure subpopulation of IFP, producing a true type r≥1 curve. The dotted lines describe theoretical curves with IFP:ISP ratios as shown. Data points are from an average of 2 complete reconstruction experiments using independently mixed stocks containing the following IFP:ISP ratios—(triangle) 100 IFP:1 ISP; (square) 10 IFP:1 ISP; and (diamond) 1 IFP:1 ISP. Monolayers of developmentally aged CEC were infected with increasing doses of these premixed virus preparations and were incubated at 40.5°C. After 24 h the supernatants were harvested, processed, and assayed for type-I IFN content. Error bars represent the mean±SD. CEC, chicken embryo cell; ISP, IFN induction-suppressing particle.
Comparison of expected and observed results in the IFN-induction dose–response curve with changes in the ratio IFP:ISP
To test the validity of this model a series of IFN-induction dose–response curves were generated experimentally from virus preparations reconstructed with known ratios of IFP:ISP. This was accomplished by appropriately mixing a stock of rgA/TK/OR/71-delNS1[1–124] that naturally contained a high titer of IFP (4.3×109/mL) with a stock of rgA/TK/OR/71-NS1[1–230], which contained a high titer of ISP (4.0×109/mL). The IFP:ISP ratio was adjusted by adding an appropriate volume of ISP to the IFP stock. The doses of the viruses used in the reconstruction experiments were corrected to account for these dilutions so that the number of IFP delivered to the monolayer were normalized. The data points shown in Fig. 3 represent the average of 2 experiments. Considering the complexity of the protocol and the biological system, the relatively good fit of the experimental curves over a large range of IFP:ISP ratios appears to validate the proposed model and supports the postulate that influenza virus populations contain IFP and ISP as 2 distinct subpopulations.
Discussion
The identification and quantification of niBAP of influenza viruses revealed large subpopulations of novel particles that expressed phenotypes with the potential to influence the pathogenesis of influenza virus and the course of the disease (Marcus and others 2005, 2009, 2010; Cauthen and others 2007; Ngunjiri and others 2008), and to modulate the efficacy of LAIV (Hai and others 2008; Wang and others 2008; Kochs and others 2009; Richt and García-Sastre 2009; Steel and others 2009). Foremost among them are subpopulations of IFP because of the IFN they induce (Marcus and others 2005). Furthermore, influenza viruses are quite sensitive to the action of IFN, although a small fraction of the infectious virus may be transiently resistant (Sekellick and others 2000). IFN also can function as a natural adjuvant (Tough and others 1996; Nagao and others 1998; Bracci and others 2005; Bracci and others 2006; Marcus and others 2007; Hai and others 2008; Wolff and Ludwig 2009), providing a link between the innate and adaptive immune systems (Tough and others 1996; Marcus and others 2010). Many delNS1 mutants expressing truncated NS1 proteins have proved to be effective as LAIV in several species (Wang and others 2008; Richt and García-Sastre 2009; Steel and others 2009; Wolff and Ludwig 2009) with the IFN-inducing capacity of the virus closely associated with its attenuation in the host (Hai and others 2008; Wang and others 2008; Kochs and others 2009; Richt and García-Sastre 2009; Steel and others 2009).
The induction of IFN and its expression by cells was subject to regulation by another niBAP common to influenza virus populations, ISP. ISP can be detected and quantified to demonstrate that a single ISP coinfecting a cell with IFP blocks IFN production (Marcus and others 2005). Figure 3 shows a type r≥1 IFN-induction dose–response curve generated by a pure population of IFP (solid line) and the deviations expected based on the presence of heretofore undetected small subpopulations of ISP (dotted lines), with the 2 kinds of distinct particles acting independently of each other as expected from a Poisson distribution. The good fit of the expected and observed IFN-induction dose–response curves generated from populations of virus reconstructed to contain different known ratios of IFP:ISP supports a model in which an IFP-infected cell otherwise programmed to produce IFN fails to do so when coinfected with 1 or more ISP (Fig. 3). Notably, both the magnitude and shape of the IFN induction dose–response curves were shown to be dependent on the content of ISP in the reconstructed mixtures of IFP and ISP. These results account for the anomalously lower than expected yields of IFN sometimes observed at high m
This represents the second example in which niBAP that express different phenotypes constitute distinct subpopulations relative to each other. Thus, defective-interfering particles do not interfere with the expression of noninfectious cell-killing (apoptosis-inducing) particles and vice versa (Marcus and others 2009, Fig. 5). This means that populations of influenza virus contain at least 4 subpopulations of niBAP (defective-interfering particles, noninfectious cell-killing particles, IFP, and ISP) that express distinct phenotypes each of which can be quantified in the presence of the others (Marcus and others 2005; Ngunjiri and others 2008; Marcus and others 2009).
Based on the important role that the IFN system plays in the generation of effective LAIV (Wang and others 2008; Richt and García-Sastre 2009; Marcus and others 2010), it would seem advantageous to design vaccines in which the generation of IFP is maximized and that of ISP minimized. However, in contrast to 2 ineffective LAIV that generated type r≥1 with relatively low yields of IFN, 2 candidate LAIV that were effective as vaccines consistently generated type r=2 IFN-induction curves that produced high peak yields of IFN, but a steep down-regulation of IFN production at high m
Footnotes
Acknowledgments
This study benefited from the services provided by the Animal Cell Culture Facility of the Biotechnology-Bioservices Center of the University of Connecticut, and primary chicken embryonic cells obtained from Charles River SPAFAS, Inc. (Storrs, CT). The research was supported by a donation to the University's Virus and Interferon Research Laboratory.
Author Disclosure Statement
No competing financial interests exist.