
Abstract
Histamine is involved in the execution of an immune reaction. Receptors for histamine, of which four different subtypes are known so far, are found on dendritic cells and on T cells. Via these receptors, histamine either indirectly or directly affects the activation of T cells. Data in the literature regarding the involved receptor subtypes and the mode of action of histamine on T cells are somewhat contradictory and depend on the type of cells analyzed, polarized T cells, or freshly prepared T cells within the context of the whole splenocyte population. Therefore, we analyzed the effect of histamine on murine T cells within splenocytes in a detailed manner. We stimulated freshly prepared splenocytes in the presence or absence of histamine with α-CD3 in vitro and analyzed the induced cytokine production. We show that histamine reduced the α-CD3-induced interferon-γ (IFN-γ) production of CD4+ cells via the histamine H2-receptor. Moreover, the effect of histamine on the α-CD3-induced IFN-γ production could be transferred within conditioned splenocyte supernatants induced by histamine (in the absence of α-CD3). Thus, the histamine effect is mediated by a soluble factor, which, however, is neither of the classical anti-inflammatory mediators, interleukin-10, or transforming growth factor-β.
Introduction
T
H1R and H2R are expressed ubiquitously, while the expression patterns of H3R and H4R are more restricted. With reference to their clinical relevance, H1R and H2R are found on vascular endothelial cells, smooth muscle cells, and in parietal cells of the stomach, but moreover also in the heart, in the central nervous system, and on several immune cells (Schubert and Peura 2008, Jutel and others 2009). The H3R is expressed mainly in cells of the nervous system (Bakker 2004, Masaki and Yoshimatsu 2010), while the H4R is expressed in bone marrow cells and in bone marrow-derived peripheral cells (Huang and Thurmond 2008).
Thus, dendritic cells and T cells express the histamine receptor subtypes H1R, H2R, and H4R, but not H3R. In addition to the functions just mentioned, several lines of evidence suggest a role for histamine in the regulation of T cell function, supported by the enhanced Th1 immune response in mice lacking endogenous histamine production (Jelinek and others 2007). In murine splenocytes, histamine reduces the induced expression of the lead Th1-type cytokine, interferon-γ (IFN-γ) (Osna and others 2001), probably mediated by the reduced synthesis of autocrine acting nitric oxide (Koncz and others 2007). However, using polarized Th1-cells, other authors show an accelerating effect of histamine on the induced IFN-γ production (Jutel and others 2001, Noubade and others 2007).
In the present study, we hypothesized that histamine acts, on the one hand, on pure polarized T cells in a direct manner; on the other hand, it acts on T cells in the context of splenocytes indirectly, transduced via histamine-responsive accessory cells. This hypothesis is supported by the notion that the observed effect of histamine on polarized T cells is mediated by the H1R (Jutel and others 2001, Noubade and others 2007), while in the splenocyte-based assay, it acts mainly via the H2R (Osna and others 2001). Using splenocytes from C57Bl/6 mice, in the present study, we demonstrate that histamine reduces the α-CD3-induced IFN-γ production H2R-dependently via induction of a soluble mediator, thus, in an indirect fashion. Both the histamine-responsive cells and the IFN-γ-producing cells are found in the CD4+ fraction of splenocytes. However, the histamine-induced regulatory factor is neither of the classical anti-inflammatory mediators, interleukin (IL)-10, or transforming growth factor-β (TGF-β).
Materials and Methods
Histamine receptor ligands
Histamine, mepyramine, and famotidine were obtained from Sigma-Aldrich (Taufkirchen), and thioperamide was purchased from Tocris (Bristol). JNJ7777120 (1-[(5-chloro-1H-indol-2-yl) carbonyl]-4-methylperazine) was kindly provided by Dr. Armin Buschauer, University of Regensburg, Germany. Histamine was solved in water, while the histamine receptor antagonists were solved in dimethyl sulfoxide (Sigma-Aldrich). For all substances, stock solutions of 10 mM concentration were prepared.
Preparation of splenocytes
Female C57Bl/6 mice at 8 weeks of age were obtained from Elevage Janvier (Le Genest-Saint-Isle). They were housed and fed in the animal facility of the Hannover Medical School. Spleens were removed after killing mice by an overdose of a mixture of xylazin (70 mg/kg bodyweight; Bayer) and ketamine (230 mg/kg bodyweight; Albrecht) followed by cervical dislocation. Animal procedures were performed according to §4 of the German Animal Welfare Protection Act after a previous report to the State Government of Lower Saxony. Spleens obtained from IL-10-deficient (IL-10−/−) C57Bl/6 mice were a generous gift from Dr. André Bleich, Central Animal Laboratory, Hannover Medical School. In order to generate single-cell suspensions, spleens were homogenized in complete medium (RPMI 1640 medium supplemented with 5% [v/v] fetal calf serum (FCS), 2 mM L-glutamate, 50 μM 2-mercaptoethanol, 100 U/mL penicillin, and 100 μg/mL streptomycin; all from PAA). After five min incubation on ice, sedimented fragments were discarded, and the remaining cells were transferred into a new vial, pelleted, and re-suspended in ACK solution (155 mM NH4CL, 10 mM KHCO3; Sigma-Aldrich) in order to eliminate red blood cells. After incubation, cells were washed twice by centrifugation and re-suspension in complete medium. Where indicated, total splenocytes were separated into CD4+ and CD4− cells by magnetic cell separation (Mouse CD4+ T cell isolation kit II and AutoMACS; Miltenyi Biotech), essentially as described by the manufacturer. Success of separation was analyzed by flow cytometry using antibodies directed against CD4 and CD8 molecules (Miltenyi Biotech). Finally, the number of cells in suspension was determined, and complete medium was added to the cell suspension to obtain a final density of 5×106 cells/mL.
In vitro stimulation of splenocytes (direct histamine effect)
96-well microtiter plates (w/o tissue culture treatment; Greiner Bio-One) were coated by adding 100 μL/well phosphate-buffered saline (PBS) containing 5 μg/mL α-CD3 antibody (clone 145-2C11) and incubation overnight at 4°C. Wells were washed twice with PBS (Sigma-Aldrich) and then filled with 100 μL/well of complete medium with or without histamine (Sigma-Aldrich) and/or other ligands. Finally, 5×105 (100 μL) splenocytes were added to each well to obtain a total volume of 200 μL/well and incubated at 37°C for 20 h. Where indicated, splenocytes were preincubated for 15 min with histamine receptor ligands.
Detection of an indirect histamine effect
In order to produce histamine-induced conditioned medium, splenocytes were stimulated overnight with histamine alone at a density of 2.5×106/mL in a total volume of 10 mL in 10 cm petri dishes (Nunc). The next day, freshly prepared splenocytes were incubated in 10 cm petri dishes (w/o cell culture treatment; Greiner Bio-One) with 10 μM famotidine (in order to prevent direct effects of histamine) for 30 min at 37°C, pelleted, and re-suspended in the cell-free conditioned medium obtained from the overnight culture supplemented with 10 μM famotidine. In some experiments, the conditioned medium was additionally supplemented with neutralizing aTGF-β antibody (R&D systems). For in vitro stimulation, these suspensions were plated on 96-well microtiter plates coated with α-CD3 antibodies and incubated as just described.
Determination of IFN-γ concentrations
After incubation, cell-free supernatants were harvested from the α-CD3 antibody-coated plates and kept on ice or, for longer periods, frozen at −80°C until analysis. IFN-γ concentrations in the obtained supernatants were determined by sandwich-enzyme-linked immunosorbent assay (ELISA) using specific antibodies (Thermo Scientific).
RNA extraction and reverse transcriptase-polymerase chain reaction analysis
RNA was extracted from 5×107 splenocytes using the NucleoSpin RNA II (Macherey-Nagel). Total RNA was reverse transcribed into cDNA using oligo dT primers and RevertAid Reverse Transcriptase (Fermentas). Histamine receptor-specific cDNA sequences were amplified using specific primers for mH1R (S: 5′-GAC CTT GGT GGA TCG ACA GT-3′; AS: 5′-TGT CTG GAA TGT GAG CGA AG-3′), mH2R (S: 5′-GCA AGC CAC AAC TCT CAC AA-3′; AS: 5′-GCT CAG GCT CAG GAG ACA AC-3′), mH3R (S: 5′-ATG CCA TTG CAC AGG TAT G-3′; AS: 5′-TTG GGA TAC CAT CTT CAT GC-3′), and mH4R (S: 5′-GAA TCA GCT GCA TCT CGT CA-3′; AS: 5′-GTG ACC TGG CTA GCT TCC TG-3′). Amplicons were analyzed by agarose gel-electrophoresis and visualized using ethidium-bromide staining.
Statistical analysis
Statistical analyses were performed by calculating means±SD. Analysis of significance was performed using one-way analysis of variance with Bonferroni post test (GraphPad Prism 5.0; San Diego). P values of <0.05 (*), <0.01 (**), and <0.005 (***) were considered significant.
Results
Histamine reduces α-CD3-induced IFN-γ production in murine spleen cells
In an initial set of experiments, we analyzed the effect of histamine on murine splenocytes. While in unstimulated splenocytes IFN-γ was not detectable (not shown), 20 h incubation in the presence of immobilized α-CD3 resulted in the release of substantial amounts of IFN-γ (Fig. 1). The concentrations of IFN-γ in supernatants of stimulated splenocytes differed between individual experiments, ranging from 10,000 to 20,000 pg/mL. Simultaneously added histamine concentration-dependently reduced the α-CD3-induced IFN-γ release of splenocytes (Fig. 1). The maximal effect of histamine was reached at a concentration of 10 μM, resulting in a roughly 50% reduction of IFN-γ release.
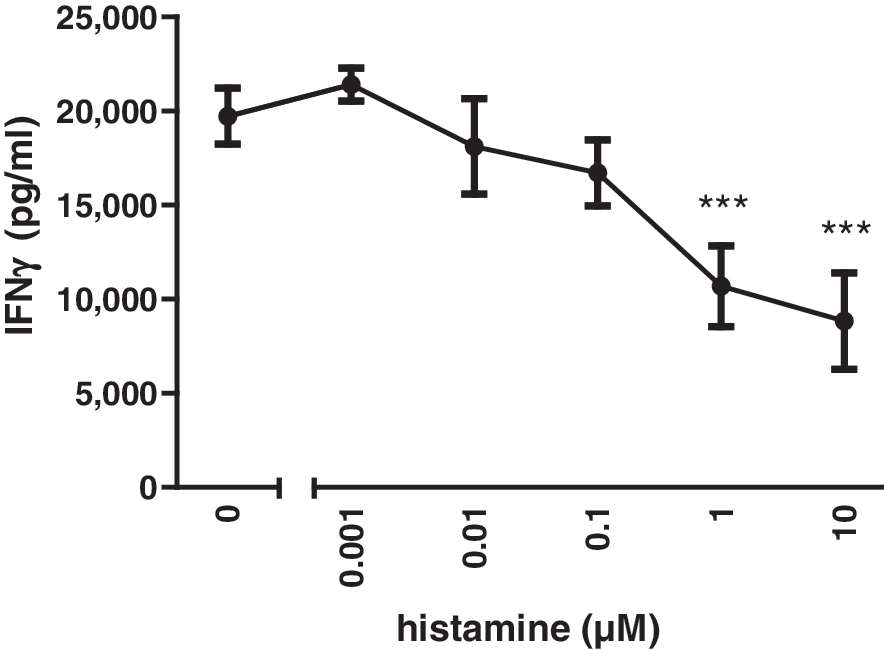
Histamine treatment reduces α-CD3-induced IFN-γ production in murine splenocytes. Freshly prepared C57Bl/6 splenocytes were incubated in the presence of plate-bound α-CD3 and increasing concentrations of histamine, as indicated on the x-axis. After 20 h of incubation, IFN-γ concentrations in the supernatants were determined by ELISA. Shown is one representative experiment out of five with similar results. (***P<0.005 vs. 0 μM histamine). IFN-γ, interferon-γ; ELISA, enzyme-linked immunosorbent assay.
The H2R mediates the effect of histamine on induced IFN-γ production
In freshly isolated splenocytes, mRNA for all four histamine receptors could be detected. However, the expression of the H3R was very low (Fig. 2A), and there is no evidence for the functional expression of H3R in immune cells (Thurmond and others 2008). In order to determine which of the four histamine receptors is/are responsible for the observed effect of histamine on the α-CD3-induced release of IFN-γ, we added selective histamine receptor antagonists during the stimulation of splenocytes. The histamine receptor antagonists mepyramine (H1R), famotidine (H2R), thioperamide (H3R+H4R), and JNJ 7777120 (H4R) themselves had no effect on the α-CD3-induced IFN-γ production (Fig. 2B). The histamine-induced reduction of the α-CD3-induced IFN-γ production was not affected by mepyramine, thioperamide, or JNJ7777120. However, the H2R-selective antagonist famotidine abolished the histamine effect in a concentration-dependent manner, reaching complete neutralization at a concentration of 10 μM (Fig. 2B).
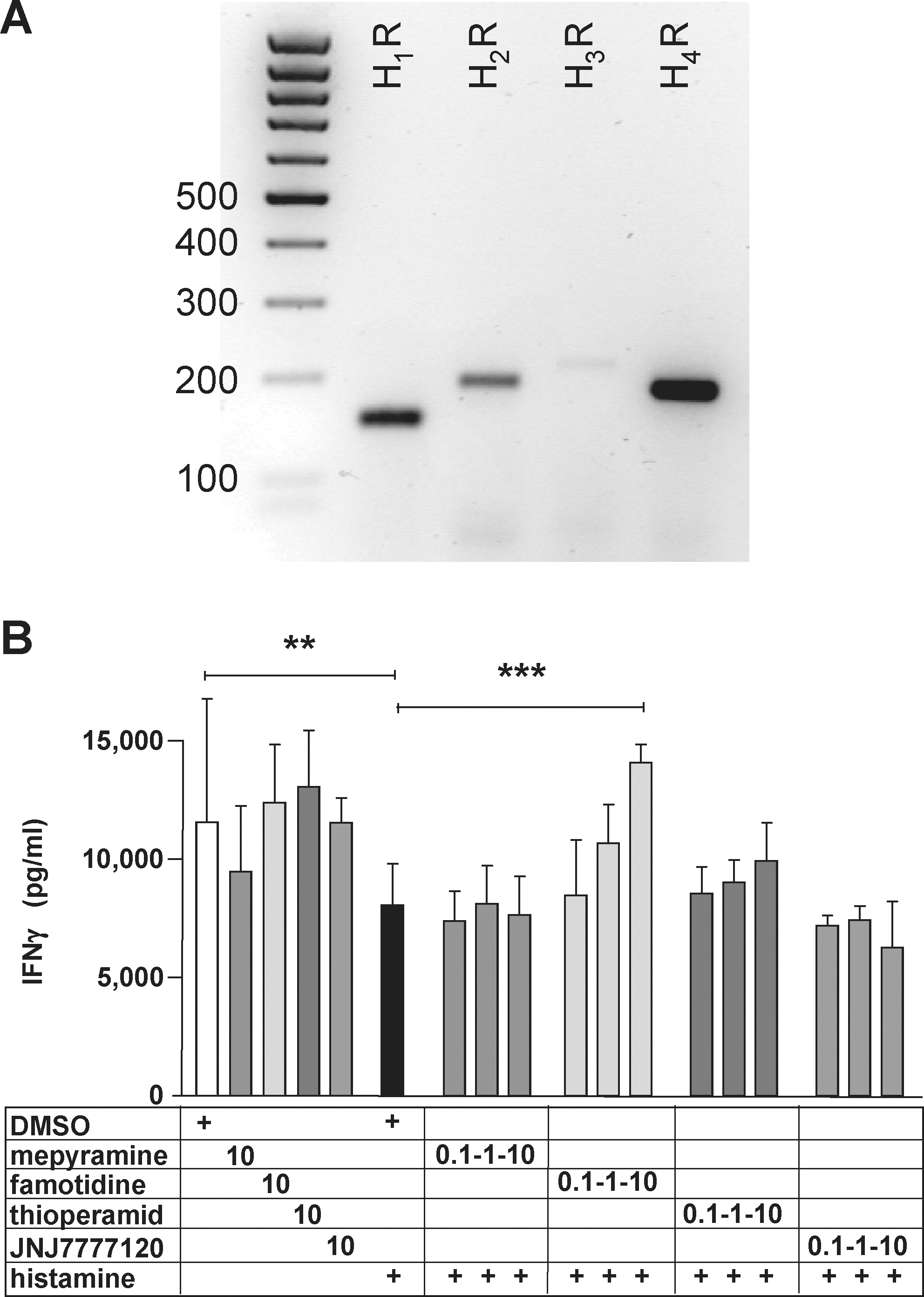
The effect of histamine on α-CD3-induced IFN-γ production in murine splenocytes is mediated via the H2R.
Histamine affects the α-CD3-induced release of IFN-γ in CD4+ cells
Splenocytes are composed of multiple cellular populations, mainly B and T cells. We analyzed whether it is the CD4+ T cell subpopulation that exhibits responsiveness to histamine. Splenocytes were either left untouched or were separated into CD4+ and CD4− populations (Fig. 3A) before in vitro stimulation with immobilized α-CD3 in the absence or presence of histamine. The quantification of released IFN-γ (Fig. 3B) revealed that histamine was effective in unseparated splenocytes and in the CD4+ subpopulation, while in the CD4− population, it had no effect.
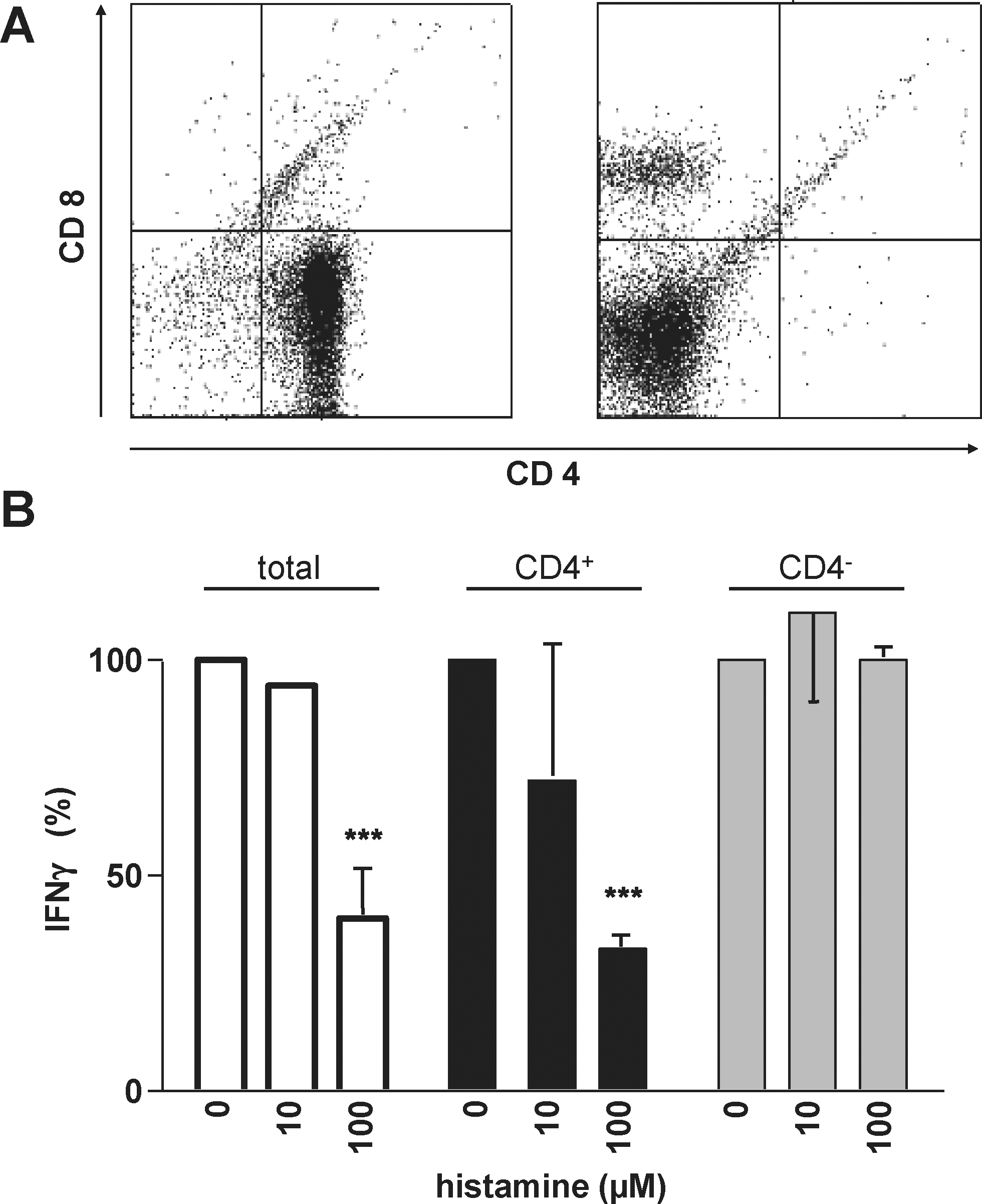
The CD4+ cell fraction out of murine splenocytes accounts for the effect of histamine on α-CD3-induced IFN-γ production. Freshly prepared C57Bl/6 splenocytes were separated into the CD4+ and the CD4− cell fractions.
The histamine effect can be transferred within conditioned medium
We next examined whether histamine directly acts on those cells that are induced by α-CD3 to release IFN-γ. For this purpose, splenocytes were incubated overnight in the absence or presence of histamine without further additions such as α-CD3. Cell-free conditioned media were prepared thereof, which did not contain detectable amounts of IFN-γ. Freshly isolated splenocytes were re-suspended in these conditioned media and stimulated with α-CD3. This incubation was performed in the presence of 10 μM famotidine in order to eliminate direct effects of histamine, which is transferred within the conditioned media (Fig. 4A). The histamine-induced conditioned media reduced the α-CD3-induced IFN-γ release as a function of the concentration of histamine used to generate the conditioned media (Fig. 4B). Such an inhibitory effect of the conditioned medium was also observed when famotidine was omitted during the second incubation (not shown). Thus, the inhibitory effect that is transferred in the conditioned medium is not mediated directly via the H2R, and, therefore, histamine, as observed in the initial experimental setting.
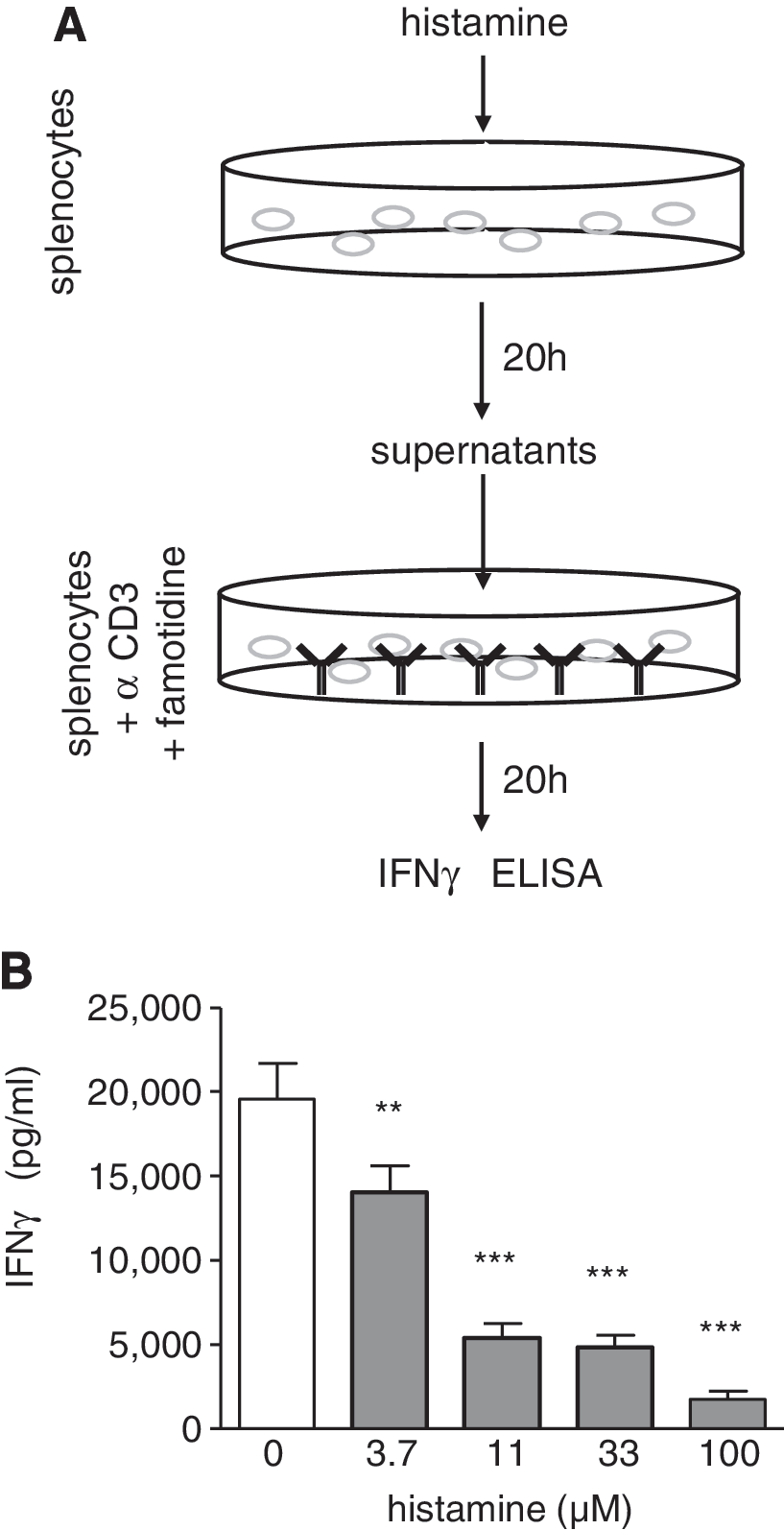
The effect of histamine on α-CD3-induced IFN-γ production can be transferred within histamine-induced conditioned supernatants.
Neither IL-10 nor TGF-β are mediators conveying the histamine effect
The two major anti-inflammatory mediators, IL-10 and TGF-β, were tested for their ability to transfer the histamine effect on α-CD3-induced IFN-γ release. Splenocytes from IL-10-deficient mice generated histamine-induced conditioned media, which as effectively as that from wild-type cells reduced the α-CD3-induced IFN-γ release of wild-type splenocytes (Fig. 5A). In addition, by direct addition of histamine during the α-CD3 antibody stimulation, histamine reduced the IFN-γ release from IL-10−/− splenocytes (not shown). Addition of recombinant TGF-β -1, -2, or -3 during the α-CD3 stimulation of splenocytes effectively reduced the release of IFN-γ, thus mimicking the effect of histamine. However, histamine did not enhance the mRNA expression of TGF-β -1, -2, and -3 (not shown). Consequently, neutralizing α-TGF-β antibodies added to the histamine-induced conditioned media did not inhibit the heir capacity to reduce the α-CD3-induced IFN-γ release in the transfer assay (Fig. 5B).
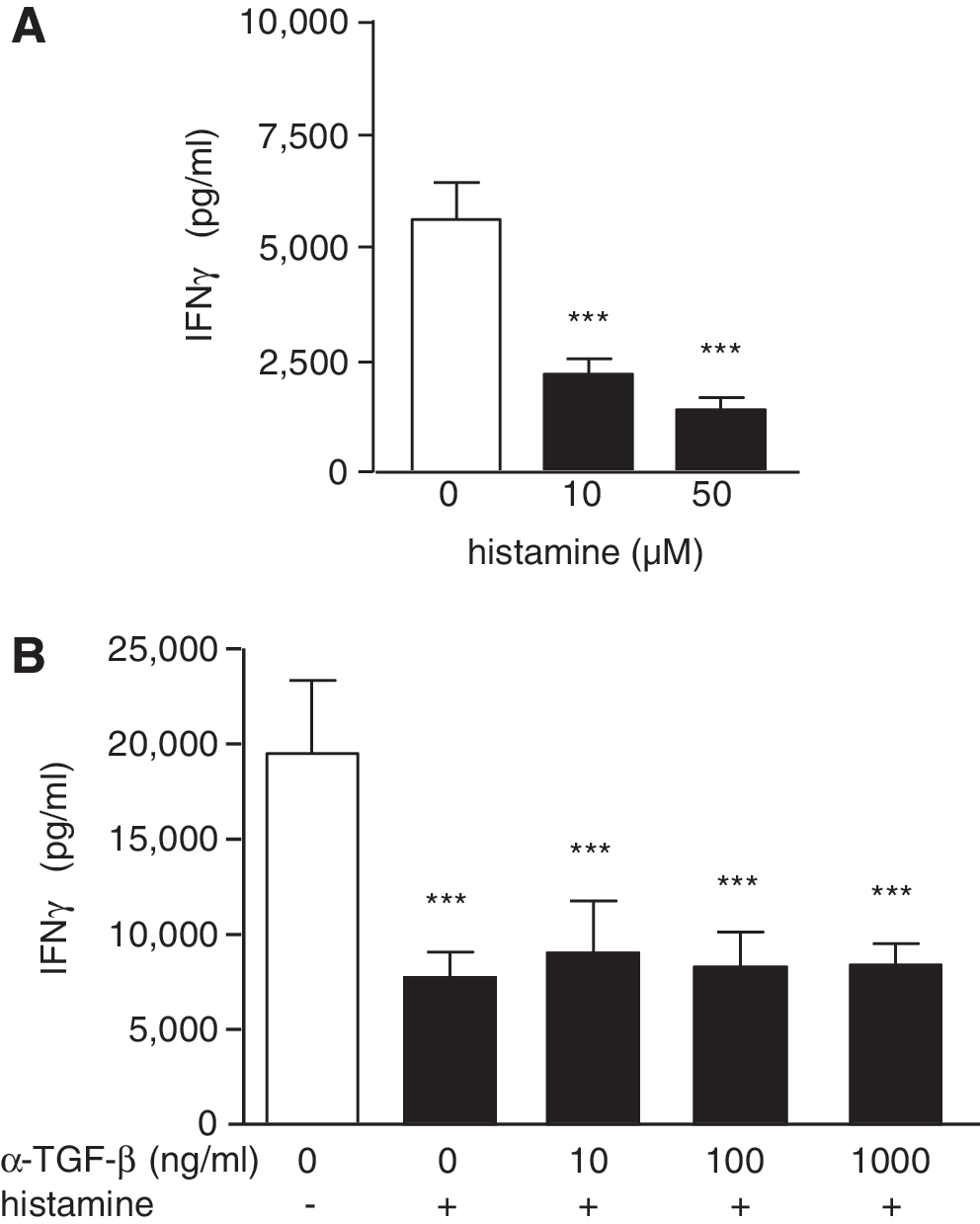
The effect of histamine on α-CD3-induced IFN-γ production is independent of IL-10 and of TGF-β.
Discussion
Histamine is a mediator molecule involved in physiological as well as in pathophysiological processes, such as the early phase of an acute allergic reaction, the transmission of neuronal signals, and the regulation of gastric acid release (Thurmond and others 2008). In these processes, preformed and intracellularly stored histamine is released from mast cells, neurons, and enterochromeaffine-like cells, respectively, in response to a specific signal (Thurmond and others 2008). Such release rapidly leads to very high local mediator concentrations. However, not only these classically known histamine-producing cells are able to generate histamine. Nearly all mammalian cells express the histamine-forming enzyme L-histidine-decarboxylase, thus, theoretically being able to generate the mediator. In contrast to the classically known histamine-producing cells, other cells, such as dendritic cells and T cells, are largely unable to store histamine and, therefore, immediately secrete the newly formed molecules (Ichikawa and others 2010). Thus, within the immune system, histamine may play a role not only in the effector phase in the periphery, but also in the priming phase in secondary lymphoid organs.
In order to analyze the effects of histamine on unpolarized T cells in the cellular context of a lymphoid organ, we used freshly isolated splenocytes throughout the entire study. In this cellular system, we were able to prove the observation that histamine reduces the α-CD3-induced IFN-γ release (Osna and others 2001). Interestingly, in a Th1-type cell clone (pGL10), histamine exerted a similar effect (Osna and others 2001), while using freshly polarized Th1 cells, the contrary, that is, the enhancement of the α-CD3-induced IFN-γ release by histamine, was observed (Jutel and others 2001). Reasons for this discrepancy may be the cellular systems used (Th1 cell clone vs. mixed Th1 cells) or their species of origin, which were mouse and human, respectively. However, using freshly polarized Th1 cells from murine spleen, we did not detect such an effect of histamine on the α-CD3-induced IFN-γ release (not shown), indicating that at least in this system, a second, inducible mediator from non-Th1 cells may be necessary.
In our study, the observed effect of histamine on the α-CD3-induced IFN-γ release was mediated via the H2R, as it was completely abolished by the H2R-selective antagonist famotidine (Mills and Wood 1989). This is in accordance with the observations of Osna and others (2001), who could inhibit the histamine effect with a different H2R-selective antagonist, cimetidine, and with another similar finding, however, in human cells (Carlsson and others 1985). In addition, Jutel and others (2001) showed in spleen cells from H2R-deficient mice after histamine-pretreatment and α-CD3 stimulation an enhanced IFN-γ production, as compared with cells obtained from wild-type mice.
In our cellular system, that is, freshly prepared splenocytes, the cells which both, are histamine-responsive and release IFN-γ are among CD4+ cells. However, the experiments so far do not necessarily determine that the IFN-γ-releasing T cells are directly responsive to histamine. Indeed, histamine stimulation of splenocytes (in the absence of α-CD3) induces an inhibitory capacity, which is transferable within the conditioned supernatants. This inhibitory capacity is not histamine itself, as the transfer experiments were performed in the presence of famotidine, which was shown initially to completely abrogate direct histamine effects. Thus, the histamine effect on the α-CD3-induced IFN-γ-release by T cell is at least in part indirect; the possibility of an additional direct effect of histamine (in the “direct” experimental setting), however, cannot be formally excluded. In summary, the histamine-responsive cells, which produce the soluble regulatory factor, express the H2R and are CD4+. These parameters may indicate regulatory T cells (Treg) being responsible for the histamine-induced reduction of α-CD3-induced IFN-γ release. Two of the main soluble mediators that are produced by Treg and are able to reduce cytokine production are IL-10 and TGF-β. These, however, could clearly be excluded to be the histamine-induced factor described in the present study.
Nitric oxide was also proposed to be a histamine-induced autocrine regulatory factor for the IFN-γ-production by Th1-cells (Koncz and others 2007). However, neither in our histamine-induced conditioned supernatants nor in the producing cells we detected nitric oxide (not shown). This, however, was not unexpected, as nitric oxide is a very short-lived molecule (Kelm and others 1991), and our conditioned supernatants are prepared 20 h after addition of the stimulus histamine.
In conclusion, in this study, we provide evidence for the existence of a soluble histamine-induced regulatory factor that suppresses α-CD3-induced IFN-γ-production in CD4+ T cells obtained from murine spleen. This factor is neither nitric oxide nor IL-10 nor TGF-β. Its identification is presently still under investigation.
Footnotes
Acknowledgments
The authors thank Dr. Klaus Resch (Institute for Pharmacology, Hannover Medical School) for stimulating discussions. The excellent technical assistance of Mrs. Anna Bauer and Mrs. Renate Schottmann is appreciated. This work was supported by a grant of the Deutsche Forschungsgemeinschaft (DFG, SFB 587) and by the EU COST action BMBS 0806.
Author Disclosure Statement
No competing financial interests exist.