
Abstract
Cardiopulmonary bypass (CPB) has been known to induce an inflammatory response that is influenced by various factors. Hypothermia is supposed to reduce inflammation after CPB. We developed an in vitro coculture model for CPB and compared the effects of hypothermia on the inflammatory response in the coculture model with results from a clinical prospective randomized trial. The coculture model consisted of endothelial cells and monocytes. Cells were stimulated with tumor necrosis factor (TNF)-α and exposed to deep hypothermia (20°C) or normothermia (37°C). In the clinical trial, 20 patients undergoing CPB for ventricular septum defect receive either normothermic (37°C) or mild hypothermic (32°C) CPB. We observed a significant interleukin (IL)-6 and IL-8 release in the coculture model 2 and 24 h after the experimental start. In the clinical trial, cytokines were significantly increased directly after weaning from CPB and remained elevated until 24 h. IL-8 and IL-6 secretions were similar in the hypothermic and normothermic group of the coculture model and the patients after 24 h. These results demonstrate that the inflammatory reaction observed in our coculture model is comparable with the cytokine increase in the blood of children undergoing CPB. Our coculture model could be useful for studies on the mechanisms of CPB-induced inflammation.
Introduction
C
To validate this coculture model based on IL secretion, we performed a prospective clinical study and randomized 20 infants to either hypothermic (32°C) or normothermic (37°C) CPB during their surgical ventricular septum defect (VSD) repair. We compared the temperature- and time-dependent cytokine release of the coculture model with the results on the inflammatory response from the clinical prospective study.
Materials and Methods
Cell culture
Human umbilical vein endothelial cells and monocytes (THP-1 cell line) cells
Umbilical cords were provided by the Department of Obstetrics, Charité-Universitätsmedizin Berlin, Germany. Informed written consent was obtained from all donors. The study was approved by the ethics committee (EA2/076/06) and conformed to the Declaration of Helsinki. Human umbilical vein endothelial cells (HUVEC) were prepared as previously described (Diestel and others 2008, 2009). Briefly, HUVEC were cultured in endothelial cell growth medium (Biochrom) in culture flasks coated with 0.2% gelatin. A confluent monolayer was passaged with trysin/EDTA (Biochrom). Experiments were performed with HUVEC passages 0–4 from at least 3 different umbilical cords.
The THP-1 monocyte cell line was obtained from the American Type Culture Collection. The cells were maintained in RPMI with 10% fetal calf serum, 1%
An endothelial cell monolayer was seeded and cultivated for 24 h under normothermic conditions. At the experimental start, the same amount of THP-1 cells was added to the endothelial monolayer.
Dynamic cooling and rewarming protocol of the coculture
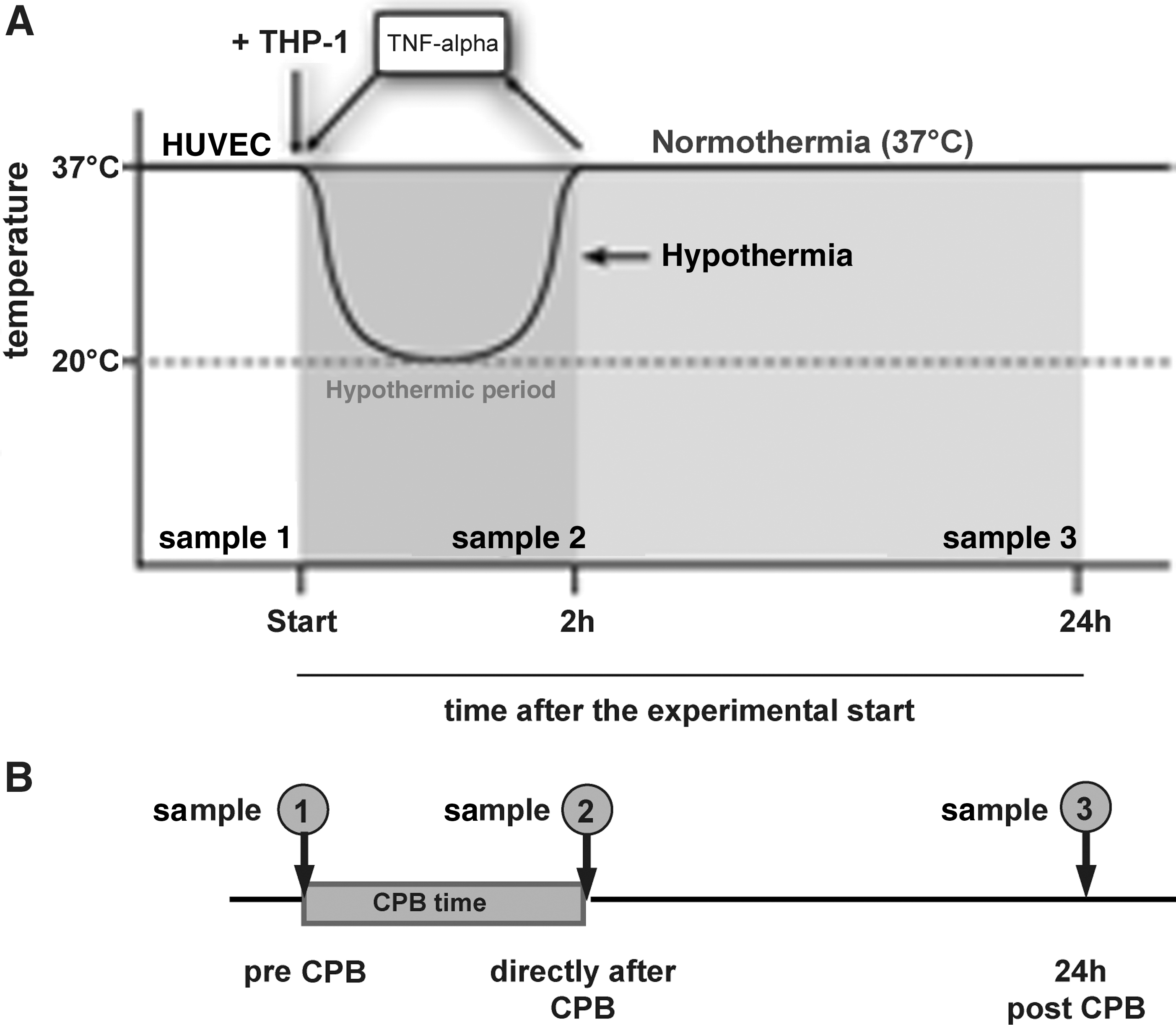
The cells were incubated under temperature conditions according to a time–temperature protocol simulating hypothermia exposure during pediatric cardiac surgery, as previously described (Diestel and others 2008, 2009). At the experimental start, hypothermic exposure was implemented in an incubator (Mytron) with an atmosphere of 5% CO2 at 100% humidity. Samples were collected at 0 h (sample 1), 2 h (sample 2), and 24 h (sample 3) after the experimental start. To simulate the inflammation after CPB, the cells were treated with 500 U/mL TNF-α (Biochrom). A normothermic control was maintained at 37°C throughout the duration of the experiments as well as unstimulated cells (Fig. 1A).
Immunocytochemistry
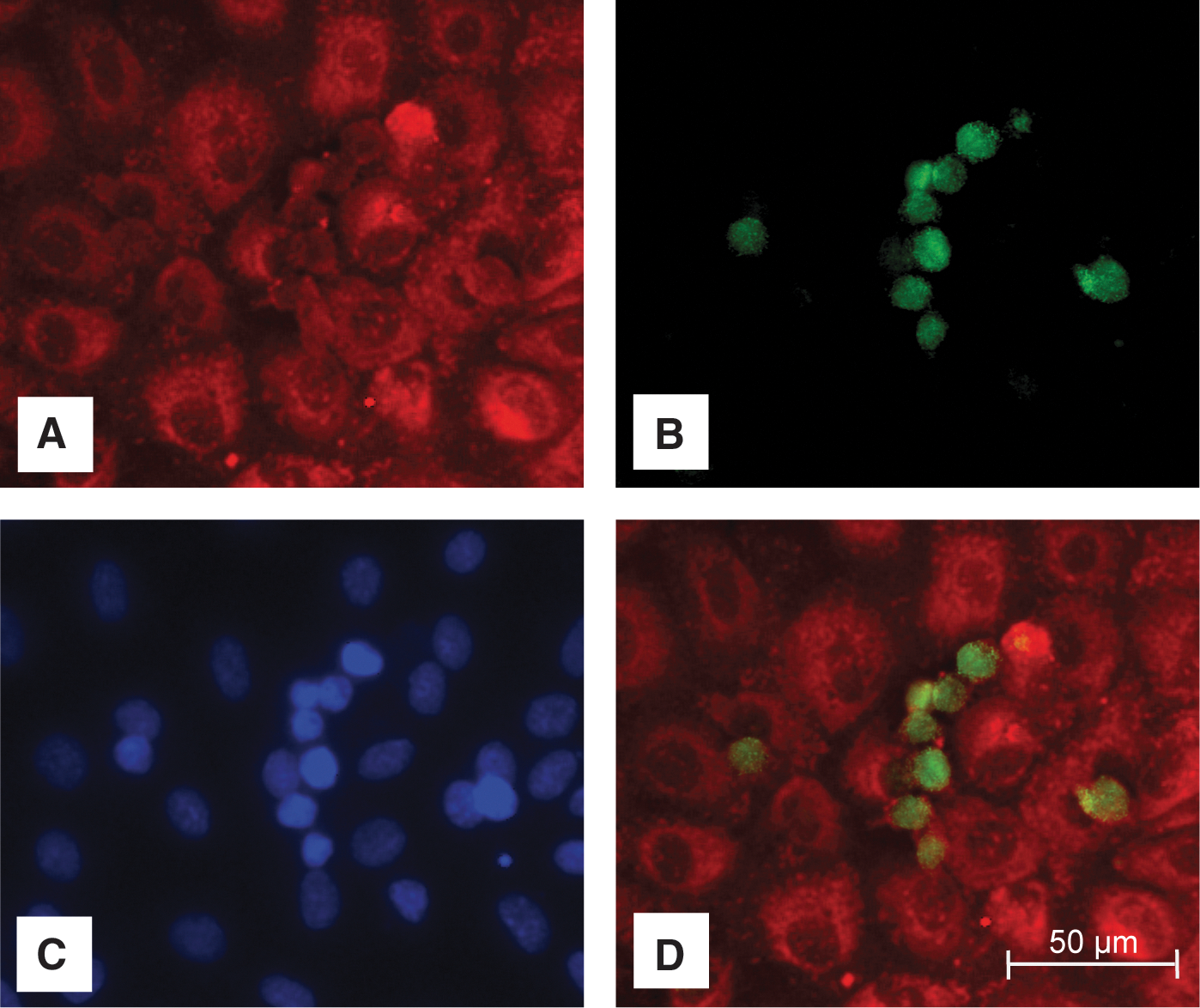
Endothelial cells and macrophages were stained with cell-specific antibodies and analyzed by immunofluorescence microscopy for cell differentiation (Fig. 2). HUVEC (2×105) and THP-1 cells (2×105) were cultivated on cover slips (Thermonox; Nunc) coated with 0.2% gelatin in 24-well plates. The coculture cells were incubated according to the time–temperature protocol previously described (Fig. 1A). We used CD31 (Dako) as a marker for the cytoskeleton of both cell types. THP-1 cells were stained using surface protein CD11b antibody directed against activated monocytes (Biomol). Nuclei were stained with DAPI (Pierce Biotechnology). Cells were fixed with 4% paraformaldehyde (Herbeta) and fixed with a fluorescence mounting medium (Dako). We used an Axio Imager M1 (Zeiss) for immunofluorescence microscopy.
Immunofluorescence microscopy of the coculture.
MTT cell viability assay
The viability of the coculture cells was measured using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Biochemica). The cells were treated in 96-well plates as just described (Fig. 1A). MTT reagent was added at each sampling time and incubated for 2 h at 37°C in a humidified 5% CO2 atmosphere. Acidic isopropanol (0.04 M HCl in absolute isopropanol) was used to dissolve the insoluble purple formazan product into a colored solution. Absorbance was measured at 570 nm using a microplate reader (Multiscan Ascent; Thermo Electron Corporation).
ELISA
For this method, we used 2×105 HUVEC and 2×105 THP-1 cells to analyze IL-6 and IL-8 cytokine secretion in the coculture model. The experimental setting was established as previously described (Fig. 1A). At each sampling time, the supernatants containing floating monocytes as well as dead cells were collected and centrifuged at 2,000 rpm. IL-6 and IL-8 concentrations were measured by ELISA (BD Biosciences) according to the manufacturer's instructions. Absorbance was measured at 450 nm using a microplate reader (Multiskan Ascent).
Clinical setting
Patients
Between May 2008 and November 2009, 20 patients within a median age of 8.3 (range 3.3–38.2) months and median body weight of 7.3 (range 5.2–11.7) kg were enrolled in a prospective clinical study. All the patients were diagnosed with a ventricular septum defect by echocardiographic analysis. Demographic data of the patients are shown in Table 1. In the hypothermic group, 10 infants underwent cardiac surgery at mild hypothermia with a median temperature of 31.8°C (range 30.1°C–33.5°C). In the normothermic group, 10 infants were operated under normothermic CPB with a median temperature of 36.1°C (range of 34.9°C–37.0°C). Surgery was performed with standard techniques. The study was approved by the ethics committee of Charité-Universitätsmedizin Berlin, and informed parental consent was given for all patients.
Clinical data were distinguished due to normothermic and hypothermic group. Age, body weight, CPB time, cross-clamp time, and minimal temperature are given in median and range.
CPB, cardiopulmonary bypass.
Anesthesia and extracorporeal circulation
Anesthetic and extracorporeal circulation techniques were the same in both groups. Midazolam (0.1 mg/kg oral) was administered as premedication. The induction of anesthesia was achieved by a combination of Sufentanil, Etomidat, and Myorelaxanta (Pancuronium) and maintained with Sufentanil and Propofol. Prophylactic antibiotics (cephalosporines) were given during and after surgery. Temperature was monitored through the bladder, esophagus, and intra-arterial lines.
CPB was performed with a mast-mounted pump (Stöckert Instrument), and a Capiox RX05 oxygenator (Terumo) was used for perfusion. The extracorporeal circuit was filled with a standard solution, containing heparin (5,000 IU), tranexamacid (100 mg/mL), and 200 mL Deltajonin. A heparin level of 6 IU/mL was maintained for anticoagulation. The concentration was monitored by Hepcon HMS Plus (Medtronic) and after CPB, antagonized with Protamin. CPB was started when activated clotting time passed 480 s. Cardioplegia was obtained with cold (4°C) Kirsch cardioplegic solution [magnesium-L- hydrogen aspartate (2.575 mg), Procain-HCl (0.3 g), Xillit (4.5 g), in aqua ad injectabilia]. Before the initiation of CPB, the patients were either cooled to a target temperature of 32°C or maintained in a normothermic condition, depending on the randomized grouping. Blood flow was nonpulsatile with a minimum pump index of 1.1±0.18 L/min in the normothermic group and 1.1±0.25 L/min in the hypothermic group. Rewarming in the hypothermic group was initiated after removal of the aortic cross-clamp, with a temperature gradient of 2°C/min. A hematocrit above 25% and a body temperature of 36.5°C were achieved before weaning from CPB. A cellsaver (Electa 5.0; Sorin Group) was used for autotransfusion of the remaining blood during the CPB and for packed erythrocytes, intraoperatively given. Then, a hematocrit of <25% was used as an indication for homologous blood transfusions.
Blood sampling protocol
Blood samples were obtained from the central venous cannulas at the following time points: preoperatively, after the induction of anesthesia (sample 1), immediately after weaning from the CPB and reversing heparin with protamine (sample 2), and 24 h after weaning from the CPB (sample 3) (Fig. 1B). Two venous blood drops were dropped onto a dry filter card of the type, which is usually used for neonatal screening. After drying for 5–6 h, the cards were shrink wrapped and stored at −70°C.
Extraction of analytes from dried blood spots
Dried blood spots (DBS; approximate drop size of 50 μL) were eluted, and proteins were extracted in a 48-well plate using 200 μL PBS. DBS were incubated for 1 h at room temperature and 650 rpm on a shaker (Eppendorf). Extracted proteins and the filter paper were transferred to an Ultrafree-MC filter unit, pore size 0.65 μm (Millipore), and briefly spun down without any loss of relevant analytes (data not shown). Residual cell debris was removed by another centrifugation step at 3,000 g for 10 min at room temperature.
Multiplexed sandwich immunoassays—Luminex xMAP technology
The Luminex xMAP technology is a bead-based immunoassay that uses color-coded microspheres as a solid support matrix. Each distinct bead set can be coupled with a different capture antibody and pooled for sample incubation. The bound analytes were detected using a fluorescence-based reporter system. The signal intensities were measured in a flow cytometer, which quantified the amount of captured analytes on each individual bead. This set-up allowed for the simultaneous measurements of analytes requiring minimal amounts of sample material (Joos and others 2002; Dunbar and others 2003). Analytes were investigated for IL-6 and IL-8 using a human cytokine/chemokine Milliplex Kit (MPXHCYTO-60K; Millipore). Measurements were conducted in duplicates using 25 μL per well of the DBS extracts. Assays were performed according to the manufacturer's protocol and measured using a Luminex 100 system (Luminex Corp.).
Statistical analysis
GraphPad Prism 4 and SPSS software were used for statistical analysis. Normal distribution was not available when the data were tested. Therefore, nonparametric statistics were used. All values are given as median. Differences between the groups were analyzed by Mann–Whitney U test, and P values of <0.05 were considered statistically significant.
Results
Morphology of the coculture model
Immunocytochemistry was performed to distinguish endothelial cells from macrophages in the direct coculture model (Fig. 2). The cytoskeleton of both cell types was stained with CD31 (Fig. 2A). The macrophages were determined by CD11b staining (Fig. 2B). Nuclei were labeled with DAPI (Fig. 2C). An overlay of CD31 and CD11b illustrates the direct coculture of endothelial cells and macrophages (Fig. 2D), consisting of a confluent monolayer of endothelial cells overlaid with adherent monocytes.
Preserved cell viability of unstimulated and TNF-α stimulated macrophages and endothelial cells after 24 h
We investigated the vitality of the coculture cells 24 h after the experimental start by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. No significant differences were observed between the 4 test groups and the unstimulated control at 37°C (Table 2).
No significant differences were observed between the 4 groups.
TNF-α, tumor necrosis factor-α.
IL-6 and IL-8 cytokine secretion of the coculture model increases over time and is temperature independent after 24 h
In unstimulated cells, there was an increase of IL-6 and IL-8 release after 24 h as a result of the interaction of the immunomodulatory cells. No difference was seen between the normothermic and hypothermic groups (Fig. 3A, B). For IL-6: 3.43 pg/mL versus 1.55 pg/mL after 2 h; 35.33 pg/mL versus 36.62 pg/mL after 24 h; for IL-8: 42.46 pg/mL versus 31.94 pg/mL after 2 h; 578.3 pg/mL versus 514.8 pg/mL after 24 h.

Cytokine secretion of interleukin (IL)-6 and IL-8, measured from the coculture model
We also observed a significant increase of IL-6 and IL-8 concentrations over time in the TNF-α stimulated normothermic and hypothermic groups. The cytokines concentration in the samples that had been taken directly after the experimental start (sample 1) were just above detection level (data not shown). After 24 h (sample 3), the IL-6 secretion in the stimulated hypothermic group (387.6 pg/mL) was equal to that of the stimulated control groups incubated at 37°C (362.4 pg/mL), which indicated that the cytokine secretion was temperature independent at this time point. In fact, 2 h after the experimental start (sample 2), the IL-6 concentration in the hypothermic group was significantly reduced in comparison to the IL-6 concentration of the normothermic group (4.3 pg/mL versus 19.7 pg/mL). There was no difference in the IL-8 concentration of the normothermic versus hypothermic group 2 and 24 h after the experimental start (75.0 pg/mL versus 42.8 pg/mL after 2 h; 12,132.3 pg/mL versus 14,250.0 pg/mL after 24 h) (Fig. 3C, D).
IL-6 and IL-8 concentration in the clinical samples elevates over time and is temperature independent
Blood samples were directly obtained (sample 2) and 24 h (sample 3) after weaning from the CPB (Fig. 1B). We observed an elevation of IL-6 and IL-8 over time in the normothermic and hypothermic group. The levels of both cytokines increased throughout the observation period, as the cytokine concentration in the samples that had been taken preoperatively (sample 1) were just above detection level (data not shown). The observed cytokine release over time was equal in both groups and was, therefore, temperature independent. Thus, median IL-6 secretion values of 26.13 pg/mL (sample 2) increased to 85.8 pg/mL (sample 3) in the clinical group who underwent normothermic cardiac surgery. The patients operated on under hypothermia (32°C) had a median IL-6 secretion value of 21.0 pg/mL (sample 2) and 66.2 pg/mL (sample 3) with a significant increase over time (Fig. 3E). For the IL-8 secretion, we observed similar results as for the IL-6 secretion. Under normothermic cardiac surgery, the measured median IL-8 concentrations were 91.29 pg/mL (sample 2) and 198.15 pg/mL (sample 3), while cardiac surgery at 32°C resulted in IL-8 concentrations of 79.4 pg/mL (sample 2) and 232.1 pg/mL (sample 3) (Fig. 3F).
Discussion
The aim of this study was to establish a coculture model that enables in vitro studies on inflammatory reactions occurring under hypothermia in pediatric open-heart surgery. For validation of this coculture model, we used the clinical data of a prospective pediatric clinical trial to investigate whether our coculture model is comparable in terms of cytokine release (IL-6 and IL-8) to the clinical setting. We investigated that the cytokine concentration of IL-6 and IL-8 increases in the coculture model consisting of endothelial cells and monocytes over time with a similar dynamic compared with IL secretion measured from VSD patients after cardiac surgery under CPB. Moreover, we compared the temperature effect in the cell-culture model with that of the clinical trail. After 24 h, the pro-inflammatory cytokine secretion was observed to be temperature independent in both the coculture model and the clinical data. To our knowledge, this is the first investigation comparing the cytokine release in a coculture model consisting of endothelial cells and monocytes with the IL secretion in the blood of children undergoing cardiac surgery with CPB.
Coculture models are commonly used in experimental studies to investigate the interaction of various cell types under distinct external conditions. Langer and others (2010) described the influence of thrombocytes and its interaction with the endothelium in the context of coronary artery disease. As another example, Eberl and Moser (2009) established coculture models of 2 different T-cell types and monocytes to investigate the acute inflammation in bacterial infection.
Only a few coculture models investigate the cell interaction concerning post-CPB inflammation in pediatric open-heart surgery. Therefore, we established a coculture model of endothelial cells and monocytes. It is well known that the endothelium plays a pivotal role during CPB-induced inflammation, which is characterized by a state of general endothelial activation and partial dysfunction (Beghetti and others 1998). The activated endothelial cells are linked to the inflammation system via protein expression (Boyle and others 1997). The cellular immune system is central to the inflammatory reaction after CPB. It is observed that the CPB-triggered inflammation leads to a spontaneous activation of monocytes and granulocytes. Especially the activated monocytes, via an up-regulation in monocyte chemoattractant factor production, increase cytokine production (Tsuchida and others 1997; Greilich and others 2008). To imitate the CPB-induced inflammation, TNF-α is commonly used to stimulate the endothelium in cell culture models (Diestel and others 2008, 2009; Makó and others 2010). In a randomized controlled trial, Jansen and others (1992) documented that there is an increase in TNF-α concentration after CPB in patients undergoing CPB. Therefore, we used TNF-α as a stimulus of our coculture model. In order to investigate the temperature effect, the interaction of both monocytes and endothelial cells needs to be addressed (Diestel and others 2009). Moreover, only a few studies are available that address the interaction between monocytes and endothelial cells under hypothermic and hypoxic conditions. For example, Wang and others (2008) demonstrated that the adhesion of monocytes to endothelial cells is dependent on early post–hypoxia-hypothermia conditions. As a consequence of these findings, we decided to use endothelial cells and monocytes for our coculture model. Since the diffuse dysfunction of the endothelium observed during CPB-induced inflammation can result in possible changes in cell structure, we chose a direct cell-culture model rather than an indirect model (Beghetti and others 1998).
The cell vitality after 24 h was similar in the hypothermic and the normothermic group (Table 2). We interpreted the equivalent cell vitality at the end of the experimental time as proof of a correctly used TNF-α concentration and amount of monocytes on top of the confluent endothelium monolayer, both of which were retained for our experiments. Immunofluorescence microscopy was applied to differentiate endothelial cells from monocytes. Since some of the monocytes became adherent, we conclude that through the interaction between the cell types, some monocytes were activated and became macrophages (Fig. 2).
For validation of the coculture model, we chose 2 ILs (IL-6 and IL-8) closely related to the outcome of the patient. The measured pro-inflammatory concentration of IL-6 is shown to correlate with the mortality and hemodynamic instability of pediatric patients who underwent cardiac surgery with CPB (Oka and others 1992; Hauser and others 1998). The pro-inflammatory chemokine IL-8 is important for chemotaxis and tissue inflammation (Laffey and others 2002). Furthermore, IL-8 secretion is known to increase in patients with systemic inflammatory response syndrome after cardiac surgery.
To compare the cytokine increase observed in our coculture model with clinical data, we enrolled 20 children diagnosed with a ventricular septum defect in a pediatric prospective randomized clinical trial. In order to minimize various factors that can influence the CPB-induced inflammatory response, we included a relatively homogenous group of patients with similar treatment, operation, and duration of CPB. Appachi and others (2007) demonstrated a significantly higher inflammatory reaction during cardiac surgery in neonates with hypoplastic left heart syndrome compared with neonates with transposition of the great arteries. Therefore, we decided to include only patients with the same CHD in order to minimize the cytokine secretion differences caused by the treatment of different CHDs.
In the clinical trial, as well as in our newly established coculture model, we observed an elevation in IL-6 and IL-8 concentrations directly after CPB until 24 h after weaning from the CPB, which is in line with the increase in the coculture model 2 h until 24 h after the experimental start. In our coculture model, we found an elevation over time for both cytokines in the unstimulated groups as well as in the TNF-α-stimulated groups. We interpreted the greater cytokine concentrations as a succeeded TNF-α of the cell-culture model. Therefore, we concluded that the results from our coculture model are comparable with the data observed in our clinical trail.
In a further step, we investigated the influence of hypothermia in both study settings and observed that 24 h after CPB, cytokine secretion occurred in a temperature-independent manner. Hypothermia has been suggested as playing a protective role in reducing acute inflammation, but there are scant data demonstrating a significant response after hypothermic CPB (Appachi and others 2007).
In order to investigate whether the IL secretion in our coculture model is temperature independent, and, thus, prove that the coculture is a reliable model which studies inflammation, we used deep hypothermia (20°C). Interestingly, we observed a significant decrease in IL-6 2 h after the experimental start (t2) in the hypothermic group compared with the normothermic group (37°C) of the coculture model. We assumed that this decrease was due to deep hypothermia used in the coculture model, which is in line with results from former studies of our group and others (Speziale and others 2000; Tassani and others 2002; Schmitt and others 2007; Diestel and others 2010). However, this decrease might not be seen in the clinical trial where only mild hypothermia was used. After 24 h, IL-6 secretion was temperature independent in the coculture model and the clinical investigation. Therefore, we were able to conclude that temperature differences only play a pivotal role in early IL-6 secretion. IL-8 levels do not seem to be influenced by different hypothermic conditions, as observed in the coculture model as well as the clinical data.
Several limitations of this investigation have to be outlined. We investigated an in vitro coculture model mimicking the inflammatory reaction of IL-6 and IL-8 during clinical applications. Inflammation induced by CPB is a complex interaction between large varieties of pathways of the inflammation/hemostatic system, but in the cellular coculture model, we only stimulated the cells with TNF-α. Moreover, similar to monocytes, neutrophils and red blood cells are also critically involved in the initiation and maintenance of inflammation. Additionally, the in vivo situation of inflammation in patients who undergo open-heart surgery is much more complex. However, a cell-culture model is well suited to elucidate the basic effects of hypothermia on the inflammatory response of distinct cell types. In further studies, this coculture model could be used to investigate the intracellular mechanisms of cellular response to hypothermia and to investigate cellular reactions on pharmacological modulation of the inflammatory response after hypothermia. Based on our findings and the proof of concept elucidated in the present study, we suggest that the inflammatory response to hypothermia and rewarming should be analyzed in a model using whole blood component and endothelial cells.
In conclusion, our results demonstrate that the inflammatory reaction (IL-6 and IL-8 release) observed in our coculture model is comparable with the cytokine increase in the blood of children undergoing cardiac surgery with CPB. The cytokine increase starts about the same time and is temperature independent after 24 h in both the in vitro coculture and the randomized prospective clinical study. This suggests that our coculture model could be used for further studies to elucidate the mechanisms of CPB-induced inflammation and to investigate the pharmacological modulation of these reactions.
Footnotes
Acknowledgment
The authors wish to thank Anne Gale for editing the article.
Author Disclosure Statement
No competing financial interests exist.