
Abstract
The respiratory syncytial virus (RSV) is a major pathogen of the human species. This pneumovirus is a prominent cause of airway morbidity in children and maintains an excessive hospitalization rate despite decades of research. As involvement of a genetic vulnerability is a possibility supported by recent data, we addressed the question of whether the Mx gene products, the typical target of which consists in single-stranded negative-polarity RNA viruses, could alter the course of pneumovirus-associated disease in vivo. Wild-type and Bos taurus Mx1-expressing transgenic FVB/J mice were inoculated with the mouse counterpart and closest phylogenetic relative of RSV, pneumonia virus of mice. Survival data and follow-up of body weight, histological scores, lung virus spread, and lung viral load unequivocally showed that the viral infection was severely repressed in Mx-transgenic mice, thus suggesting that pneumoviruses belong to the antiviral spectrum of mammalian Mx GTPases. Elucidating the underlying mechanisms at the molecular level could reveal critical information for the development of new anti-RSV molecules.
Introduction
Mx genes encode a subclass of type-I and -III interferon-induced dynamin-like large guanosine triphosphatases (GTPases) called “Mx proteins” of which some mammalian isoforms display antiviral activity. The typical target of antiviral mammalian Mx consists in a single-stranded negative-polarity RNA virus, that is, the exact profile of pneumoviruses.
RSV infection of mice does not result in any measurable degree of morbidity, evokes a mild mononuclear cell infiltration instead of a profuse granulocytic bronchiolitis, does not result in eosinophil recruitment, never progresses to acute respiratory distress syndrome (ARDS), and generates viral titers systematically lower than that inoculated (Chambers and others 1990). Taken together, these characteristics suggest that using the RSV murine model to mimick human disease is rather counterintuitive. Therefore, the putative Mx activity against pneumoviruses in vivo was evaluated in a model using rodent natural pneumovirus pathogen, which is also the closest phylogenetic relative of RSV (Chambers and others 1990), pneumonia virus of mice (PVM). The crucial advantages of this PVM-associated model include the following: (i) clinical picture–morbidity–consistently mimicking that observed in infants with RSV-associated bronchiolitis; (ii) dramatic granulocytic and eosinophilic infiltrations that parallel the pathological changes observed in humans; (iii) clear evidence of widespread viral replication in lung tissue, with incremental recoveries that, at peak, are in excess of 108 PFU/g in response to as few as 30 PFU in the inoculum; and (iv) clear progression to ARDS as reported for ∼3% infants with RSV bronchiolitis (Haataja and Hallman 2002).
As the murine endogenous Mx1 and Mx2 fail to convey such protection in mice (D. Desmecht, personal observations), we used the Bos taurus Mx1-transgenic mouse line that was recently made available. In this transgenic line, bovine Mx1 (boMx1) is expressed to a near-physiological level under the control of its natural inducible promoter and short- and long-range regulatory elements (Garigliany and others 2009). Using this model mouse, we show for the first time that pneumoviruses belong to the antiviral spectrum of mammalian Mx GTPases.
Materials and Methods
Design
Thirty-nine mice from each of 2 lines (wild-type FVB/J and transgenic FVB/J•Bos-Mx1/ML549) were inoculated with PVM and 5 mice of each line were mock-infected. Mortality and body weight loss were monitored daily in 15 mice from each line, starting 1 day before inoculation and going on up to 15 days postinoculation (pi). At selected time intervals, a subset of each line-specific cohort was euthanized for histologic examination and lung virus quantitation.
Mice, virus and inoculation
The experiments were conducted with specific pathogen free 12-week-old female mice obtained either from Charles River Laboratories (wild-type) or from the University animal facility (transgenic). This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and with the official Belgian guidelines (
PVM strain J3666 (kindly supplied by A. Easton) was first passed in 10-week-old BALB/c mice and then grown once onto BS-C-1 cells to produce the stock solution. The stock solution was then diluted to 10−5 in modified Eagle medium, aliquoted, and stored at −80°C to serve as inoculum. Randomly selected aliquots yielded highly reproducible titers on BS-C-1 cells, amounting to ∼2.105 PFU/mL. The inoculation procedure consisted in slowly instilling 50 μL of the viral suspension (i.e., ∼103 PFU) into the nostrils of the anesthetized mouse maintained in a vertical position.
Assessment of lung lesions and virus spread
At selected time intervals (4, 5, 6, and 7 days pi), 3 mice were overdosed with sodium pentobarbital and were exsanguinated by cutting the renal artery. The lungs were inflated with formaldehyde 4% in phosphate-buffered saline (PBS) for 24 h and then processed as for routine paraffin-embedded block preparation. Five micrometer-thick sections were stained with hematoxylin-eosin and were independently examined by 2 individuals who were never aware of the strain considered, lesions being graded “0” (normal), “1” (scattered single inflammatory cells), “2” (multifocal foci of clustered inflammatory cells), or “3” (multifocal confluent clusters of inflammatory cells). Spread of PVM antigens was sought by fluorescence microscopy, after dewaxed sections of the same blocks were sequentially blocked with bovine serum albumin (BSA) 1% in PBS (45 min at 37°C), incubated with in-house anti-PVM polyclonal rabbit antiserum (diluted 1/50, 45 min at 37°C), and alexa-488 conjugated antirabbit IgGs antibodies (Molecular Probes; diluted 1/1000, 45 min at 37°C).
Assessment of lung virus yields
At selected time intervals (4, 5, 6, and 7 days pi), 3 mice were euthanized for lung virus titers quantitation by real-time quantitative polymerase chain reaction (RT-qPCR). The lungs were weighed, homogenized in ice-cold BSA 1% in PBS, and clarified (1,000 g for 10 min). Viral RNA was extracted using Nucleospin® RNA Virus columns according to the User manual (Macherey Nagel). Homogenates were treated with Fermentas DNase I and an aliquot of each RNA extract (100 ng RNA) was then reverse-transcribed using commercial high capacity cDNA reverse transcription kit (Applied Biosystems), and RT-PCR was conducted using the following PVM SH gene primers set: 5'-GCC GTC ATC AAC ACA GTG TGT-3' and 5'-GCC TGA TGT GGC AGT GCT-3', with Sybr green PCR Master Mix (Applied Biosystem) and using a StepOne plus cycler (Applied Biosystems) with a classical protocol (40 cycles, 60°C). A DNA polynucleotide corresponding to the target sequence was synthesized and a standard curve for qPCR was constructed on the basis of 6 successive dilutions of the stock solution.
Statistical analysis
A Kaplan–Meier analysis was conducted to assess survival rates and between-lines comparisons was made by using the Mantel–Haenszel test, with significance threshold set at P<0.0001. Effects of mouse line on histological score and on amount of SH gene copies per 100 ng RNA were analyzed with a Mann–Whitney test, with *P<0.05, **P<0.01, and ***P<0.001. NS, P>0.05.
Results
Survival
With the in vivo study carried out with the 15 mice that were monitored for 15 days, we compared the mortality, median survival duration, and morbidity characteristics of the 2 mouse lines after inoculation of 1,000 PFUs. In wild-type mice, we observed 87% mortality, with an average survival duration of 8 days. Survival (93%) and average duration of survival (>15 days) were significantly better among Mx-transgenic mice (Fig. 1). All mock-infected control mice survived the study.
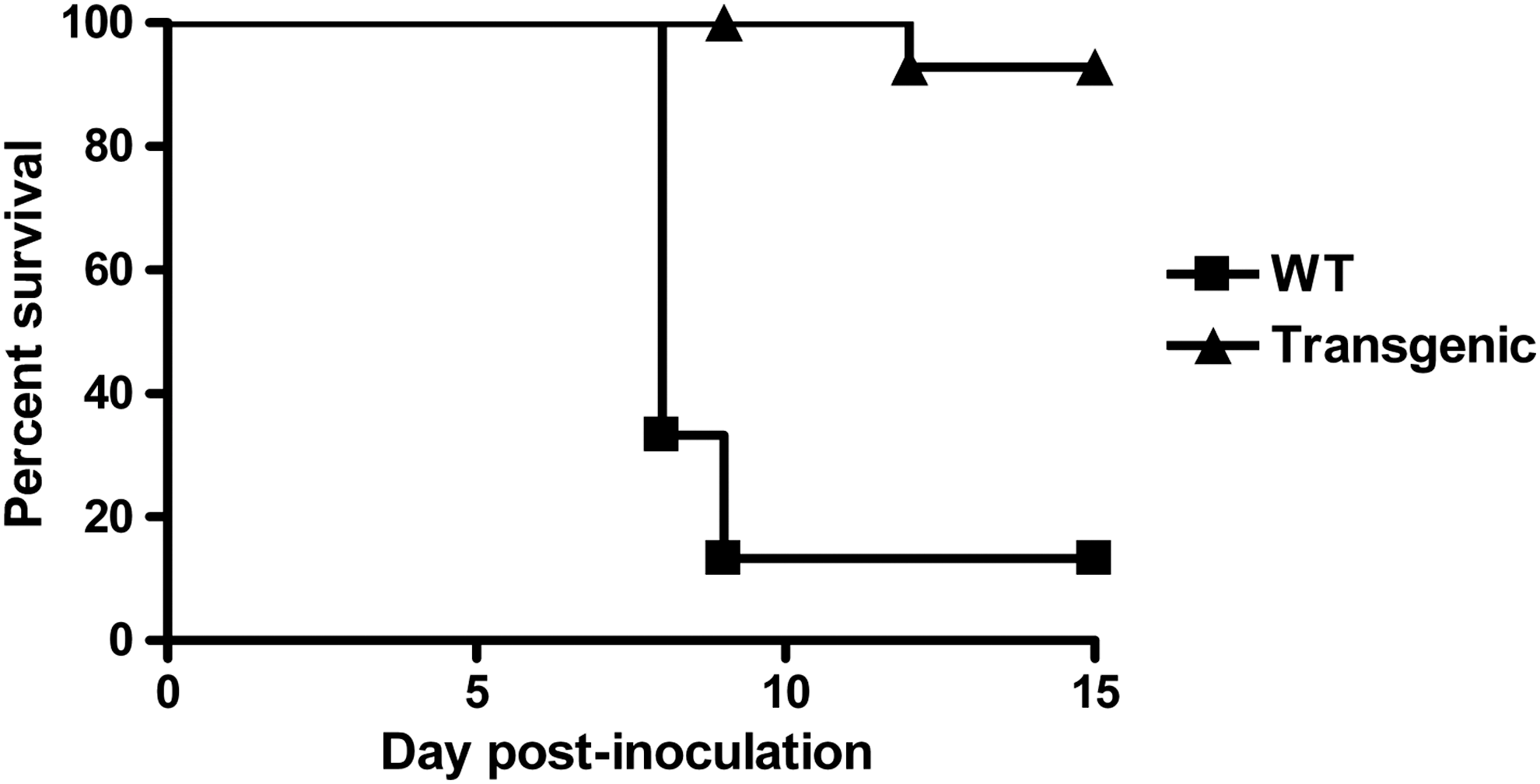
Induction of bovine Mx1 expression by intranasal inoculation of 1,000 PFUs pneumonia virus of mice (PVM) confers dramatic improvement of survival (Kaplan–Meier curve of overall survival). The FVB/J•Bos-Mx1/ML549 Mx-transgenic mouse line (transgenic) is significantly more resistant than wild-type (WT) FVB/J mice (Mantel–Haenszel test, P<0.001). Fifteen mice of each line were enrolled in the study.
Clinical and gross pathological observations
Overall, virus-associated morbidity, body weight loss, and gross lesions caused by inoculation of 1,000 PFUs of PVM were roughly similar for both lines for the first 6 days pi. From day 7 pi onward, body condition very rapidly deteriorated among wild-type mice, whereas that of Mx-transgenic mice remained stable or improved. The pathological processes remained totally asymptomatic for the first 6 days. By day 6 pi, wild-type mice showed general signs such as gradually slower, less frequent, and more erratic spontaneous displacements and a ruffled coat. They abruptly started to exhibit clinical signs of respiratory disease, including respiratory distress, labored breathing, and forced expiration. Autopsies of wild-type mice performed on day 7 pi consistently showed the dark, purplish, bulky, non crepitant, liver-like lungs compatible with a diagnosis of massive pulmonary congestion and consolidation. Isotime lungs from Mx-transgenic mice only showed scattered noncoalescent light red foci reminiscent of multifocal active congestion. Further, wild-type mice showed a ∼70% increase in lung wet weight at endpoint as compared to an increase of about 30% in Mx-transgenic mice (Fig. 2).
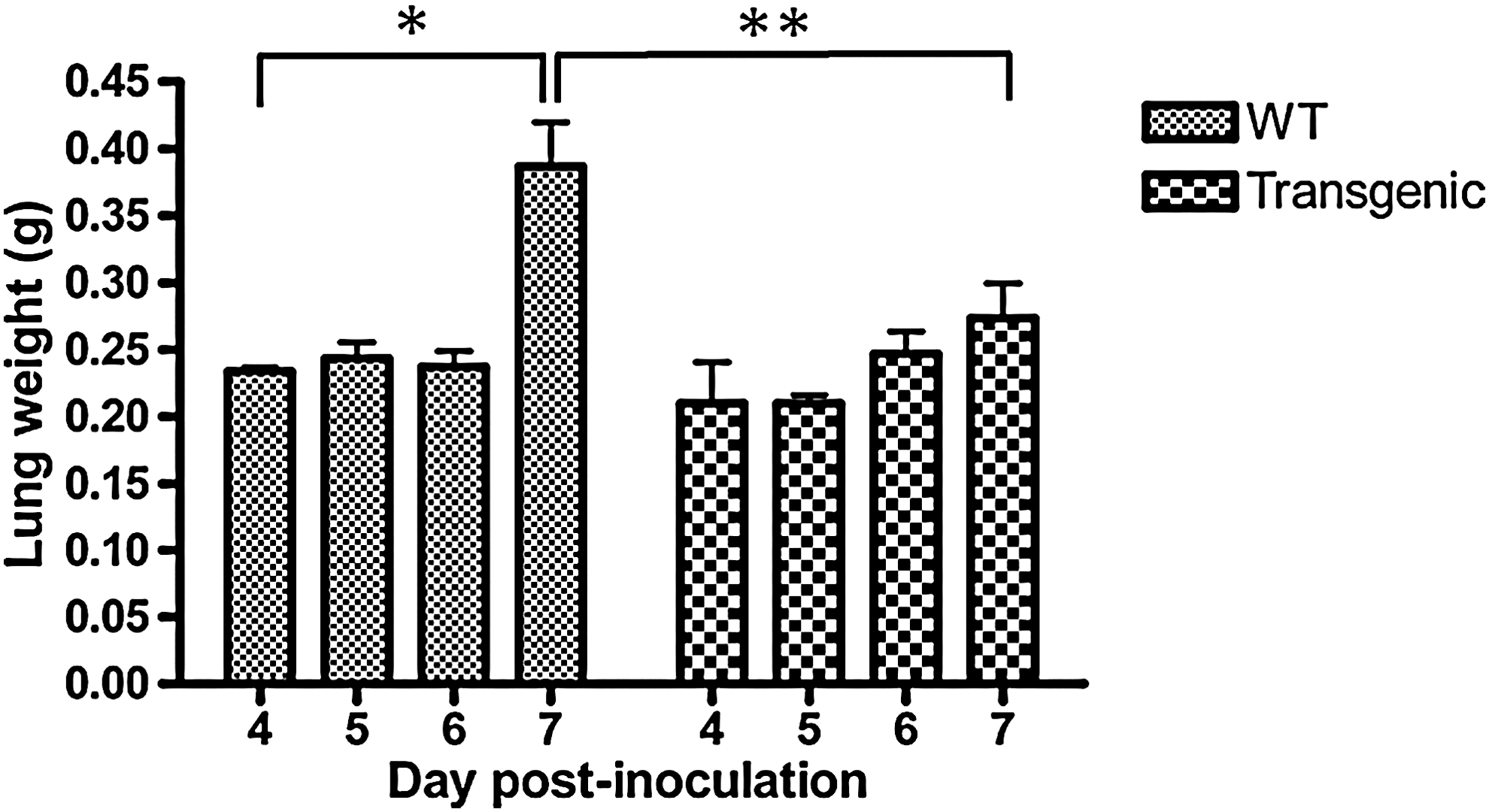
Induction of bovine Mx1 expression by intranasal inoculation of 1,000 PFUs PVM stops the explosive development of lung oedema on day 7 postinoculation (pi). Absolute values of lung wet weight are given as mean±standard deviation (SD) for 6 mice at each time point. Mean significantly different from those of control lungs are indicated (*P<0.05 and **P<0.01).
Histological and virological observations
The density of alveolar inflammatory infiltrates is reported semi-quantitatively (Fig. 3). Two clearly different histological profiles were identified, with a very dense inflammatory exsudate throughout the lungs in wild-type mice whereas only a few foci with few and far between inflammatory cells were seen in Mx-transgenic mice. The spread of the virus throughout the lungs, as judged by immunofluorescence, was dramatically different between lines (Fig. 4). In wild-type mice, scattered foci of PVM antigen-positive bronchiolar epithelial cells, type-1 and -2 pneumocytes, and alveolar macrophages were clearly visible on day 4 pi. Afterward, antigen-positive cells were homogeneously distributed throughout the lungs. Although the types of PVM antigen-positive cells were similar in lungs from Mx-transgenic mice, virus spread was delayed (no positive-cell detected on day 4 pi, observed fluorescence corresponding to red blood cell autofluorescence) and remained drastically confined in a few circumscribed foci. PVM replication data obtained on days 4, 5, 6. and 7 are gathered in Fig. 5. The lung viral load was drastically blunted in Mx-transgenic mice.
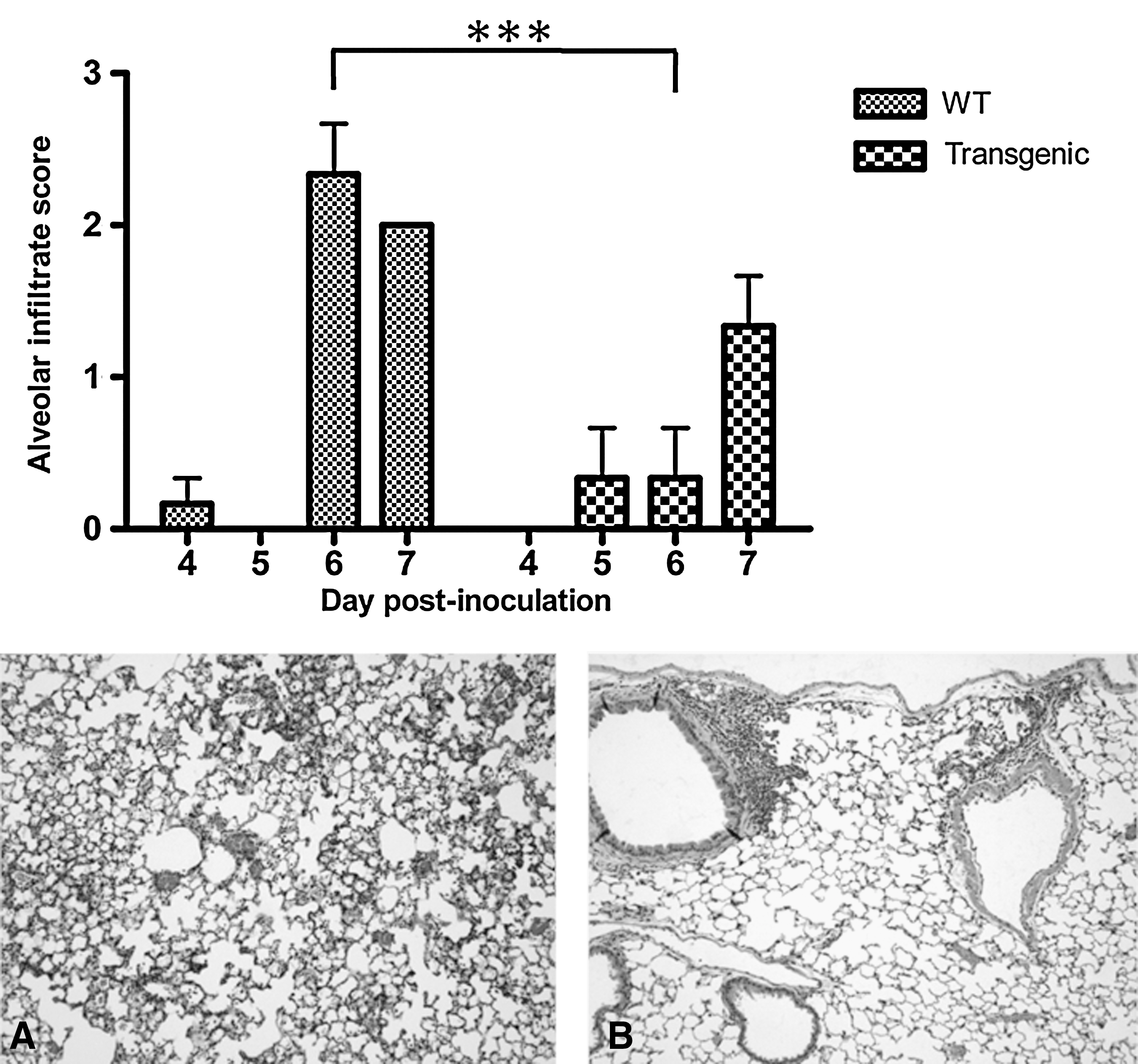
Induction of bovine Mx1 expression by intranasal inoculation of 1,000 PFUs PVM delays and attenuates the lung infiltration by inflammatory cells at said time points. Top: bar graph semi-quantitatively reporting density of alveolar inflammatory infiltrates, with ***P<0.001. Bottom: photomicrographs of hematoxylin and eosin-stained lung sections of PVM-infected mice euthanized on day 6 pi. Dramatically different histopathological signatures are observed, as described in the text (Magnification ×100).
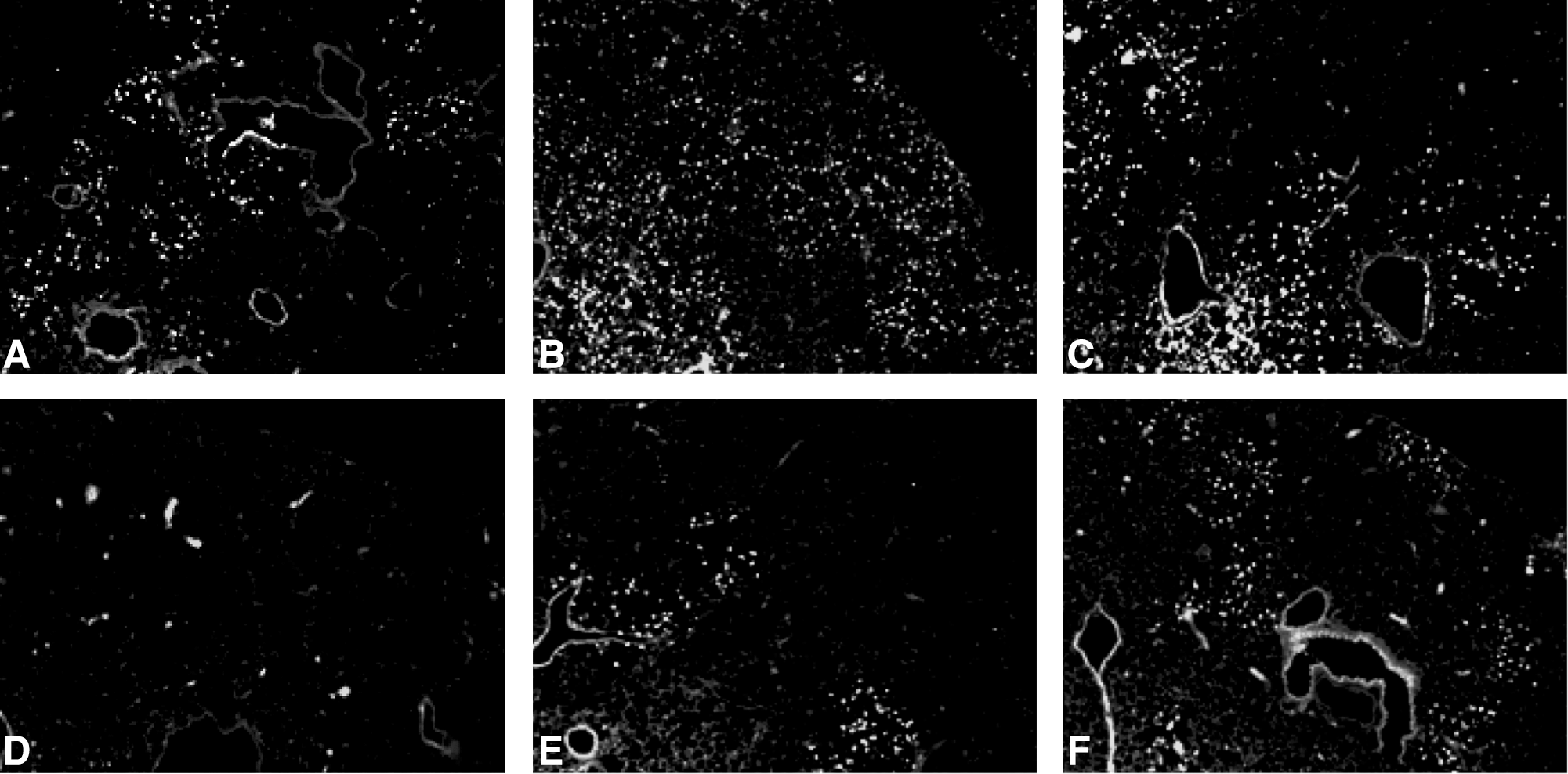
Induction of bovine Mx1 expression by intranasal inoculation of 1,000 PFUs PVM drastically restricts the spread of the virus to some well-demarcated foci. Spread of PVM antigens was sought by fluorescence microscopy after dewaxed sections were sequentially incubated with in-house anti-PVM polyclonal rabbit antiserum and alexa-488 conjugated antirabbit IgGs antibodies. In panel D, some fluorescent spots are visible; they correspond to autofluorescent erythrocytes aggregated in some capillaries.
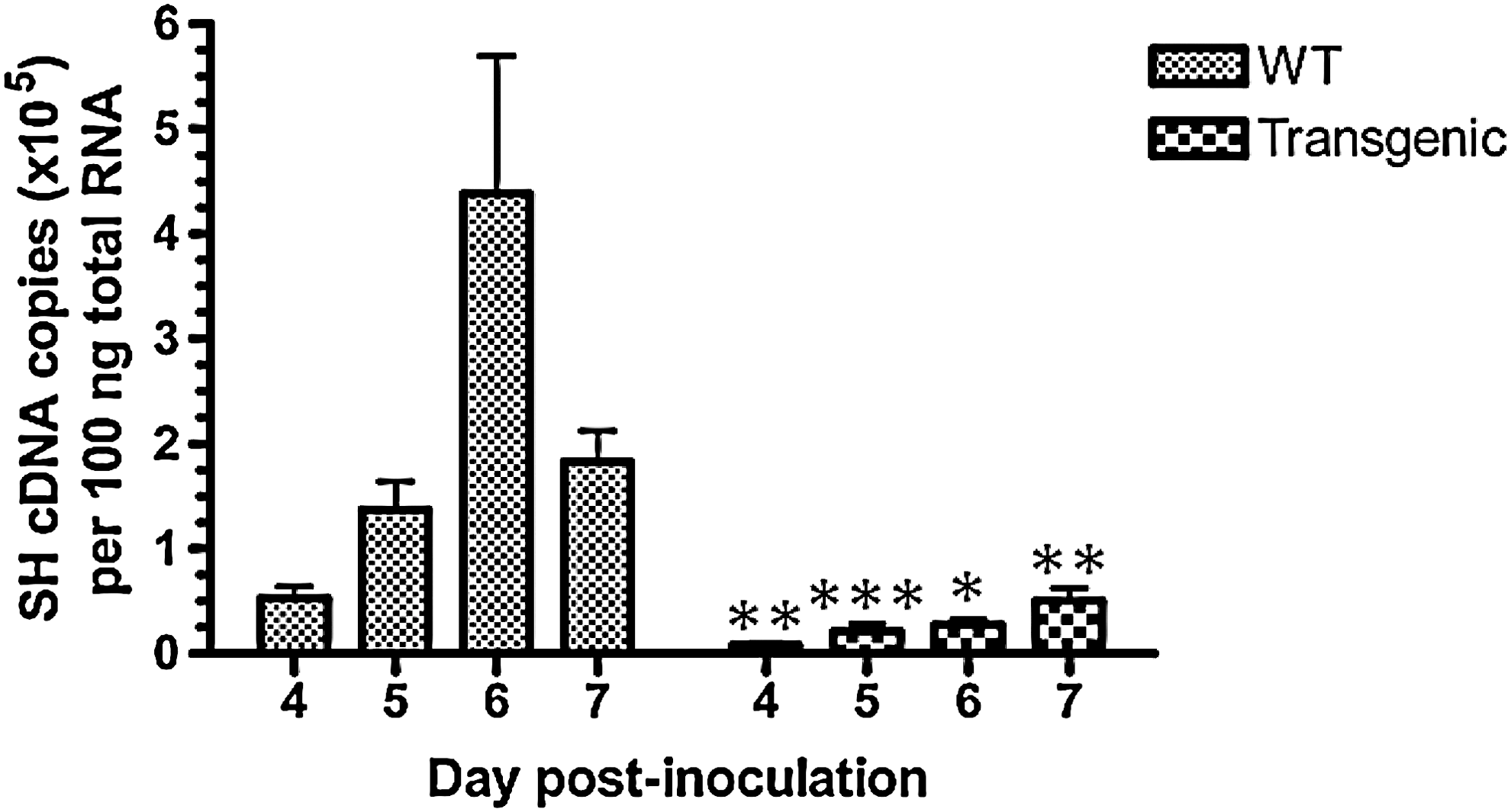
Induction of bovine Mx1 expression by intranasal inoculation of 1,000 PFUs PVM restricts lung virus amplification at said time points. Amounts of SH gene cDNA copies per 100 ng of total RNA are given as mean±SD at each time point. Means significantly different from isotime values measured in the lung of Mx-transgenic mice are indicated (*P<0.05, **P<0.01, and *** P<0.001).
Discussion
Thirty-nine mice from wild-type and Mx-transgenic mouse lines were intranasally inoculated with a standardized suspension containing the murine counterpart of RSV, PVM. The procedure caused a viral respiratory illness, as demonstrated by induced morbidity, gross lesions and lung weight, histology, and viral loads. Significant differences appeared between lines, to the extent that the disease outcome appeared like a categorical variable. Among wild-type mice, the induced respiratory disease became clinically explicit on day 7 pi, was short lasting (48 h), fatal, and consisted in peracute lung congestion, edema and pneumonia with unrestricted viral amplification. Conversely, only non specific clinical signs were recorded among Mx-transgenic mice, with some foci of pneumonia, drastically attenuated virus replication and spread and very little mortality. Expression of boMx1 therefore confers strong resistance against the mouse pneumovirus in vivo.
At first sight, the putative contribution of a cytoplasmic Mx protein with authenticated antiviral activity to innate resistance against viruses of the Paramyxoviridae family appears very plausible since the typical targets of antiviral Mx proteins known so far had previously been shown to be single-stranded negative-sense RNA viruses. For example, human MxA was shown to suppress replication of Orthomyxoviridae (Staeheli and others 1986; Arnheiter and Haller 1988; Pavlovic and others 1990; Garber and others 1991; Haller and others 1995; Thimme and others 1995), Bunyaviridae (Frese and others 1996; Kanerva and others 1996; Hefti and others 1999; Sandrock and others 2001; Andersson and others 2004), Rhabdoviridae (Meier and others 1990; Pavlovic and others 1990), Togaviridae (Landis and others 1998), and Picornaviridae (Chieux and others 2001) in vitro. Regarding members of the Paramyxoviridae family, human MxA, as expected, inhibits human and bovine parainfluenza-3 and measles viruses (Schnorr and others 1993; Schneider-Schaulies and others 1994; Zhao and others 1996). Surprisingly, however, the human pneumovirus, RSV, was shown to escape antiviral mechanisms driven by human MxA in vitro, which is reminiscent of the importance of RSV disease in vivo (Atreya and Kulkarni 1999). Cytoplasmic boMx1 belongs to the phylogenetical subset of Mx GTPases expected to be endowed with antiviral activity (Haller and Kochs 2011) and, as anticipated, it inhibits the replication of members of the Rhabdoviridae (Baise and others 2004; Leroy and others 2006; Garigliany and others 2009) and Orthomyxoviridae families (Garigliany and others 2012). Contrary to human MxA, however, Sendai, bovine and human parainfluenza-3, and measles viruses are resistant to boMx1 (Leroy and others 2005). Further, like human MxA, boMx1 does not alter replication of human and bovine RSV in vitro (Leroy and others 2005). Overall, the data available before this study was started suggested that pneumoviruses were resistant to Mx proteins with authenticated antiviral activity. The unequivocal results gathered here call this view into question. As the Mx-resistance of human (Atreya and Kulkarni 1999) and bovine (Leroy and others 2005) RSVs was demonstrated in vitro, one might argue that this does not automatically mean that the same conclusion would have been reached in vivo. However, if a significant anti-pneumovirus effect was indeed exercised by human MxA or boMx1 in vivo, RSVs would certainly not be considered as a major pathogen in both species. Alternatively, the apparent discrepancy between former and present results could be the consequence of a different cell environment: human glioblastoma or monkey renal epithelial cells for in vitro studies (Atreya and Kulkarni 1999; Leroy and others 2005) and mouse airway/lung epithelial cells here. The fact that the mechanism of human MxA-dependent resistance against measles virus infection was shown to be cell type specific lends credibility to this hypothesis (Schneider-Schaulies and others 1994). Further, the recent demonstration that the molecular scenario underlying Mx antiviral activity might involve specific interactions with some host molecules rather than with viral proteins only argues in the same direction (Wisskirchen and others 2011). Unfortunately, inoculation of human or bovine RSV in mice results in an experimental disease that remains far from recapitulating the corresponding natural diseases (Bui Tran Anh and Desmecht 2001; Bem and others 2011), which prevents future relevant comparisons in vivo. Finally, in spite of being the closest phylogenetic relative of RSV (Chambers and others 1990), PVM itself could display an Mx-susceptible specificity, either in its own proteome or in the host-virus interactome. For example, boMx1 could bind and sequester the PVM nucleoprotein like human MxA does with the nucleoprotein of Thogoto (Kochs and Haller 1999), influenza A (Turan and others 2004), or La Crosse (Kochs and others 2002) viruses. Alternatively, PVM might use an array of host cofactors that is not strictly identical to that recruited by human and bovine RSV, raising the possibility that Mx expression alters a cell pathway that is crucial to PVM, but not to RSV. Interestingly, a similar difference in sensitivity to the antiviral action of Mx proteins between 2 close phylogenetic relatives was recently reported for avian and human influenza A viruses too (Dittmann and others 2008). In this case, the viral nucleoprotein proved to be a determinant of Mx-sensitivity. Further studies analyzing RSV and PVM in vitro with respect to their sensitivities to mammalian Mx proteins are under way to elucidate the determinants of Mx-sensitivity among pneumoviruses.
Footnotes
Acknowledgments
This study was supported by grants provided by the Fonds pour la Recherche Scientifique dans l'Industrie et l'Agriculture and by the Research Council of University of Liège. The authors deeply thank Mickaël Sarlet for his skilled technical assistance and Dr. Benoît De Muynck for his precious contribution to figures realization.
Author Disclosure Statement
No competing financial interests exist.