
Abstract
The aim of this study was to investigate the effects of interferon (IFN)-γ on experimental tooth movement in mice using a murine experimental tooth movement model. An Ni-Ti closed-coil spring was inserted between the upper-anterior alveolar bones and the upper-left first molars in mice. We evaluated the relationship between local Ifn-γ mRNA levels and orthodontic tooth movement. In other experiments, IFN-γ was injected adjacent to each first molar every other day during tooth movement. After 12 days, the amount of tooth movement was measured. Tartrate-resistant acid phosphatase (TRAP)-positive cells at the pressure side of each experimental tooth were counted as osteoclasts. Local Ifn-γ mRNA expression increased with orthodontic tooth movement. The number of TRAP-positive cells increased on the pressure side of the first molar. In contrast, the degree of tooth movement and the number of TRAP-positive cells on the pressure side in IFN-γ-injected mice were less than those of control mice. IFN-γ was induced in experimental tooth movement, and could inhibit mechanical force-loaded osteoclastogenesis and tooth movement. These results suggest that IFN-γ might be useful in controlling orthodontic tooth movement because of its inhibitory action on excessive osteoclastogenesis during this movement.
Introduction
O
Orthodontic tooth movement occurs through repeated alveolar bone resorption by osteoclasts on the pressure side and new bone formation by osteoblasts on the tension side. Several studies have shown that orthodontic forces induce expression of TNF-α and suggest an important role of TNF-α in orthodontic tooth movement (Lowney and others 1995; Başaran and others 2006; Yoshimatsu and others 2006; Andrade and others 2007; Kitaura and others 2008).
Interferon (IFN)-γ also regulates the differentiation and function of bone-related cells. IFNs are crucial to the regulation of a wide variety of innate and adaptive immune responses (Kadowaki and others 2000). IFN-γ mainly contributes to T-cell-mediated regulation of the immune response and has been shown to stimulate antiviral and antibacterial activity (Billiau and others 1998). IFN-γ is secreted by T-helper 1 (Th1) cells, cytotoxic T-cells, dendritic cells, and natural killer cells (Dunn and others 2006), and has been characterized as activating macrophages by upregulating nitric oxide and major histocompatibility complex expression (Billiau and others 1998). However, the role of IFN-γ in pathological bone diseases is controversial. IFN-γ inhibits RANKL signaling by degrading TRAF6 (Takayanagi and others 2000), and IFN-γ-deficient mice show enhanced bone resorption with collagen-induced arthritis (Vermeire and others 1997). These results suggest that IFN-γ inhibits osteoclast formation. IFN-γ also inhibits TNF-α-induced osteoclast formation in vitro and in vivo (Kohara and others 2011). In contrast, recent studies have indicated that IFN-γ indirectly stimulates osteoclast formation via antigen-driven T-cell activation (Gao and others 2007). Thus, the role of IFN-γ in osteoclast formation in vivo is unclear.
In mechanical force-driven tooth movement, cellular responses in alveolar bone, periodontal ligament and other paradental tissues are mediated by interactions between various factors such as cytokines and hormones (Henneman and others 2008; Krishnan and Davidovitch 2009). IFN-γ was expressed on the pressure side of teeth in a rat tooth movement model (Alhashimi and others 2000), and has been evaluated histomorphometrically during orthodontic tooth movement, which increases the trabecular bone volume (BV/VT) (Mermut and others 2007). While these results suggest that IFN-γ plays an important role in orthodontic tooth movement, there are few studies of the effect of IFN-γ on tooth movement. Therefore, in this study, we have investigated the effect of IFN-γ in a mouse model of mechanical-loaded tooth movement.
Materials and Methods
Experimental animals
Eight-week-old C57BL/6 mice were obtained from Japan SLC, Inc. (Shizuoka, Japan). All mice were bred in our animal facilities, and received a sterilized standard diet containing 1% calcium (Oriental Yeast, Tokyo, Japan). During the experiment, the mice were kept in cages in a room maintained at 21°C −24°C with a 12 h/12 h light/dark cycle. Experiments were undertaken in accordance with the 1989 Guidelines for Animal Experimentation, Nagasaki University.
Experimental tooth movement
Under anesthesia, the orthodontic appliance was affixed to the mice using a slight modification of a method described previously (Yoshimatsu and others 2006). An Ni-Ti closed-coil spring (Tomy, Fukushima, Japan) was placed between the upper alveolar bone and upper-left first molar, and fixed with 0.1-mm stainless steel wire. The force level of the coil spring after activation was ∼10 gf (equivalent to 98 mN) according to the manufacturer. Recombinant mouse IFN-γ (R&D Systems, Minneapolis, MN) (0.015–1.5 μg per 20 μL) was injected into a local site adjacent to the upper-left first molar every other day until the end of the experiment. Phosphate-buffered saline (PBS) was used for experimental control.
RNA preparation and quantitative real-time polymerase chain reaction
Total RNA was isolated from the tissues around each upper-left first molar, including alveolar bone, on days 0, 3, 6, and 12 after tooth movement. These samples were frozen in liquid nitrogen, ground using a Multi-Beads Shocker (Yasui Kikai, Osaka, Japan), and then centrifuged in 1 mL of TRIzol reagent (Invitrogen, Carlsbad, CA). cDNA was synthesized from 1 μg samples of total RNA using reverse transcriptase (Invitrogen) and oligo-dT primers (Invitrogen) in a volume of 50 μL. Expression levels of Ifn-γ mRNA were quantified by real-time-based reverse transcription–polymerase chain reaction (RT-PCR) using the Mx3000P real-time PCR system (Stratagene, La Jolla, CA) and SYBR Premix Ex Taq (Takara, Tokyo, Japan). The relative expression of mRNA was normalized against GAPDH mRNA. Samples were preheated at 95°C for 5 min. Each cycle consisted of a denaturation step at 95°C for 30 s, an annealing step at 55°C for 30 s, and an extension step at 72°C for 30 s. The following primers were used: Gapdh: 5′-GGTGGAGCCAAAAGGGTCA-3′ and 5′-GGGGGCTAAGCAGTTGGT-3′; Ifn-γ: 5′-TCAAGTGGCATAGATGTGGAAGAA-3′ and 5′-TGGCTCTGCAGGATTTTCATG-3′.
Measurement of tooth movement
Mice were sacrificed on days 3, 6, 9, and 12, and their cephalic parts removed and fixed in 4% formaldehyde. After removing the maxillae, individual trays were placed on the maxillary teeth to take impressions with dental hydrophilic vinyl polysiloxane impression material (Exafine injection type; GC Co., Tokyo, Japan). The degree of tooth movement was evaluated by measuring the distance between the first and second molars (first- and second-molar distance) in these impressions under a stereoscopic microscope (VH-7000; Keyence, Osaka, Japan).
Histological preparation
Histological sections of maxillae taken from each mouse were fixed in 4% formaldehyde overnight, decalcified with 10% EDTA (pH 7.4) in PBS, dehydrated for 10 days at 4°C, embedded in paraffin, and sectioned at 4 μm. In the mouse first molar, the root length between the bifurcation surface and the apex is about 600 μm, which suggests that the area within the 300 μm of root from the bifurcation surface at the mesial side is under pressure during tooth movement. The alveolar crest is about 80 μm from the bifurcation surface. Thus, we prepared 5 horizontal sections from the root at 100, 140, 180, 220, and 260 μm from the bifurcation surface. These sections were used to count osteoclasts after staining for tartrate-resistant acid phosphatase (TRAP) activity and counterstaining with hematoxylin. For TRAP staining, sections were incubated in an acetate buffer (pH 5.0) containing naphthol AS-MX phosphate (Sigma, St. Louis, MO), Fast Red Violet LB Salt (Sigma), and 50 mM sodium tartrate. Mesial-side TRAP-positive cells of the distobuccal root were counted in 5 sections per mouse, and the mean values were calculated.
Statistical analysis
All data are expressed as mean±standard deviation. Student's t-test was used for statistical analyses. Values of P<0.05 were considered statistically significant.
Results
Ifn-γ was induced around force-loaded teeth after experimental tooth movement
Twelve days after inserting the appliances, all upper-left first molars had moved mesially (Fig. 1B). We analyzed the expression of Ifn-γ mRNA in tissues around each of these teeth using real-time RT-PCR to follow the time course of changes in expression of Ifn-γ. A significant difference in the level of Ifn-γ mRNA expression was observed only between days 0 and 12 (Fig. 1C).

Interferon (Ifn)-γ induced by mechanical force loading of mouse teeth.
IFN-γ inhibited experimental tooth movement
During the experiment, we injected IFN-γ into a local site adjacent to the upper-left first molar in each mouse. After 12 days of mechanical loading, tooth movement distances in mice injected with PBS were 145±44 μm. In mice administered with IFN-γ, tooth movement distances were significantly less at 56±21 μm (Fig. 2A, B). This effect was dose dependent. The degree of tooth movement in mice injected with 1.5 μg of IFN-γ was significantly lower than that in PBS-injected mice on day 12, and decreased in an IFN-γ dose-dependent manner (Fig. 2C).
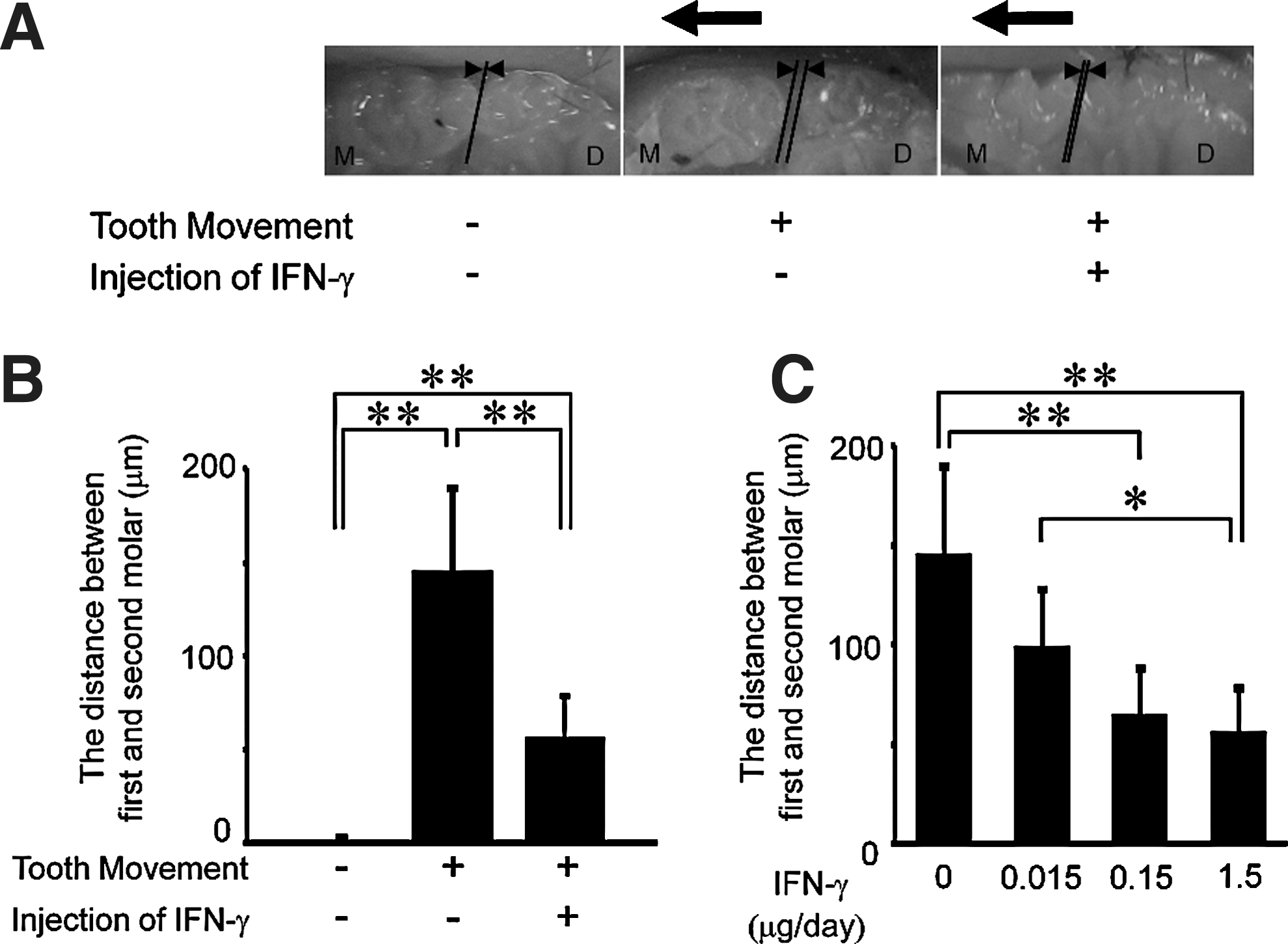
Dose dependence of IFN-γ inhibitory effects on tooth movement.
In mice injected with PBS, teeth had moved by an average of 49±22 μm on day 3 and continued to move up to day 12, reaching a distance of 145±44 μm from their starting position at this time point. In contrast, in mice injected with IFN-γ, tooth movement increased at day 3 (51±14 μm), but this distance did not increase further up to day 12 (56±21 μm). This difference between the degree of tooth movement in the 2 groups was significant on days 9 and 12 (Fig. 3).
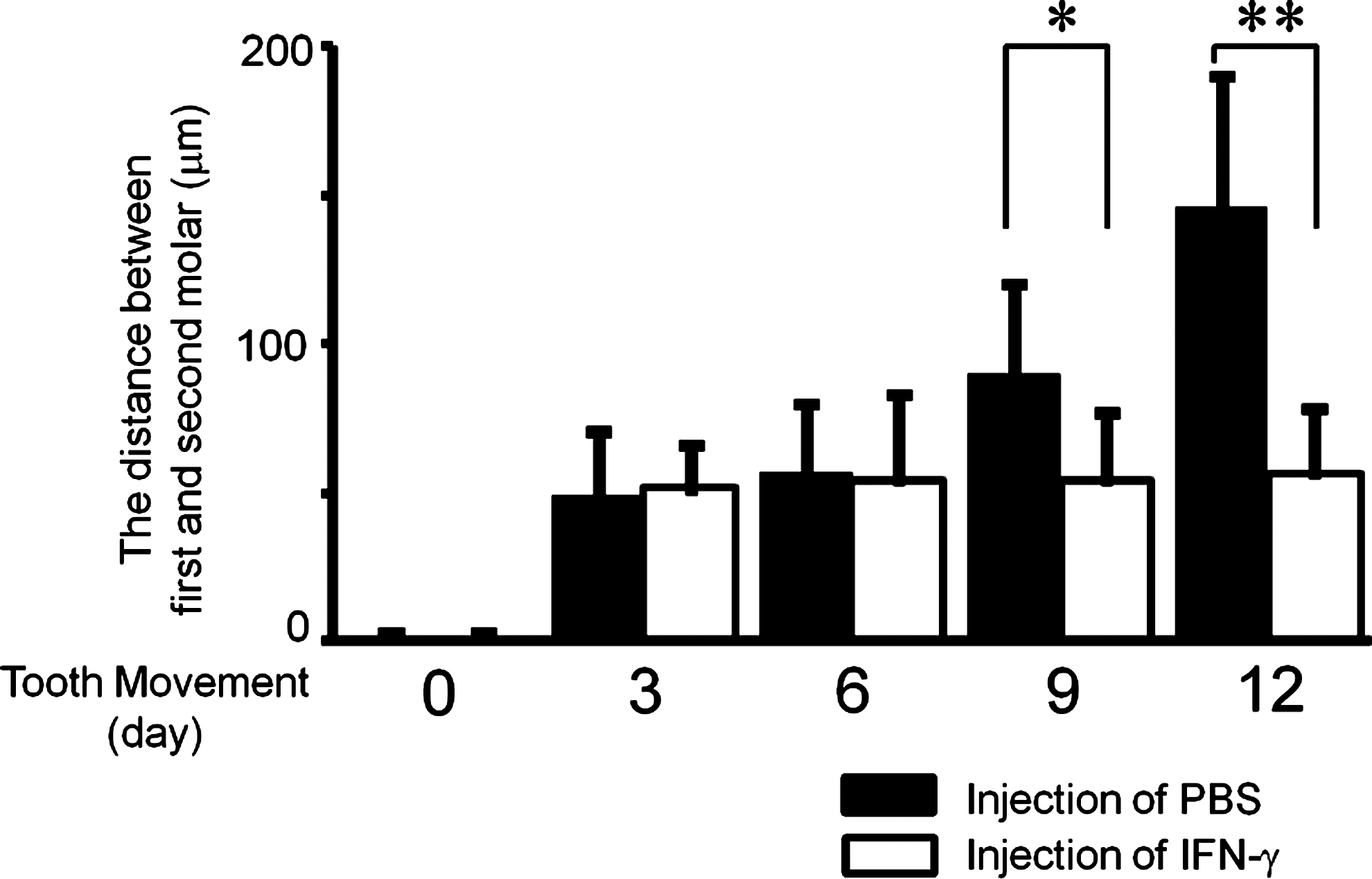
IFN-γ inhibition of experimental tooth movement after 12 days. Distance between the first and second molars before and after tooth movement for 3, 6, 9, or 12 days with administration of PBS (black bars) or 1.5 μg IFN-γ (white bars) every other day. Data are expressed as mean±SD (n=6), *P<0.05 and **P<0.01.
IFN-γ inhibited the number of TRAP-positive cells following experimental tooth movement
To confirm osteoclast formation, we made histological observations of tissues before and after the experimental tooth movement, and with and without IFN-γ injections. Tissue samples were prepared and stained for TRAP. The mesial side of the distobuccal root of each upper-left first molar was examined. Before tooth movement, teeth were positioned roughly in the center of the periodontal membrane. In contrast, after tooth movement, the teeth were forced into close contact with the mesial periodontal membrane, and many TRAP-positive cells were observed at the mesial side of the root surface (ie, in the direction of movement). In IFN-γ-treated mice, the teeth were also located against the mesial periodontal membrane, but the number of TRAP-positive cells was significantly reduced compared with PBS-administered mice (Fig. 4A, B). The number of TRAP-positive cells was decreased in an IFN-γ dose-dependent manner (Fig. 4C). The number of TRAP-positive cells in IFN-γ-treated mice and PBS-injected mice was significantly different on day 12 (Fig. 4D).

IFN-γ inhibition of tartrate-resistant acid phosphatase (TRAP)-positive cell numbers after experimental tooth movement.
Discussion
The present study showed increased expression of Ifn-γ mRNA in response to mechanical force applied during experimental tooth movement. These results are in agreement with a previous histological analysis showing that the expression of the IFN-γ gene and protein was increased by experimental tooth movement in a rat model; IFN-γ-positive cells were apparent 3 days after the initiation of force application, and these elevated levels of IFN-γ were maintained until day 10 (Alhashimi and others 2000). Other studies suggest that early-phase orthodontic tooth movement involves an acute inflammatory response (Storey 1973; Davidovitch and others 1988). In the present study, Ifn-γ mRNA was expressed on day 12 during experimental tooth movement. Thus, it is not clear whether Ifn-γ expression increases only during the early stages of movement; further analysis will clarify this expression pattern.
In this study, the effect of IFN-γ on mechanical tooth movement was investigated in mice. Our orthodontic appliance induced 2 phases of tooth movement, the first phase occurring by day 2, and the second phase occurring between days 8 and 10 (Yoshimatsu and others 2006). This first phase might be related to the finding that the periodontium at the mesial side narrows between days 2 and 8, and the distance of tooth movement increases during this time. Histological sections revealed that extensive alveolar bone resorption occurred on day 8, which may be related to the second phase of tooth movement. In this study, we found that the first phase occurs between days 3 and 6, with the second phase starting on day 9. The first phase does not involve osteoclast formation, but the second phase does involve osteoclast-mediated bone resorption. Because IFN-γ inhibited osteoclast formation, the degree of tooth movement seen in IFN-γ injected mice and PBS-injected mice after 9 days (ie, in the second phase) was significantly different. Ours is the first study showing that IFN-γ can inhibit orthodontic tooth movement. Mermut and others have previously investigated the effects of IFN-γ by histological analysis during experimental tooth movement in rats, and showed that IFN-γ increased the BV/TV and decreased the trabecular separation number during experimental tooth movement (Mermut and others 2007). In their study, 0.01, 0.02, or 0.05 μg of IFN-γ was administered into the mesiobuccal subperiosteal area adjacent to the first molar once a day. The quantity of IFN-γ (1.5 μg/day) in our present study might be sufficient to interfere with osteoclastogenesis and bone remodeling in experimental tooth movement.
In recent years, much importance has been attached to the relationship between cytokines and osteolysis. The course of osteoclast formation can be controlled by cytokines. Interleukin (IL)-6 (Gao and others 1998), IL-17 (Kotake and others 1999), and transforming growth factor-β (Fox and others 2000) induce osteoclast formation and increase bone resorption by osteoclasts. Conversely, IL-4 (Kitaura and others 2003), IL-10 (Owens and others 1996), IL-12 (Horwood and others 2001; Kitaura and others 2002; Nagata and others 2003; Yoshimatsu and others 2009), IL-13 (Onoe and others 1996), IL-18 (Udagawa and others 1997; Kitaura and others 2006; Morita and others 2010), and IFN-γ (Fox and others 2000; Kohara and others 2011) inhibit both osteoclast formation and several of their functions. In this study, IFN-γ inhibited osteoclast formation at the site where pressure builds up during experimental tooth movement, as shown previously by immunohistochemical staining in a rat model (Alhashimi and others 2000). Taken together, these results suggest that IFN-γ might control excessive osteoclastogenesis during orthodontic tooth movement.
It has been reported that IFN-γ regulates the differentiation and functionality of bone-related cells. However, its role in bone pathology is controversial. IFN-γ has been shown to inhibit RANKL signaling (Takayanagi and others 2000), and IFN-γ-deficient mice show enhanced bone resorption with collagen-induced arthritis (Vermeire and others 1997). In contrast, a study has found IFN-γ to be involved in osteoclastogenesis via T-cell activation, with IFN-γ-producing human T-cells directly inducing the differentiation of monocytes into osteoclasts via RANKL expression (Kotake and others 2005). IFN-γ also indirectly stimulates, rather than inhibits, osteoclast formation and bone loss in vivo after ovariectomy by stimulating antigen-driven T-cell activation (Gao and others 2007). However, we previously reported that IFN-γ inhibits osteoclastogenesis by a T-cell-independent mechanism (Kohara and others 2011). The current results suggest that the mechanisms underlying the outcomes observed in this study might therefore be independent of T-cell activation.
TNF-α directly promotes osteoclast differentiation, resulting in accelerated bone resorption in vitro (Kobayashi and others 2000) and in vivo (Kitaura and others 2005). TNF-α-induced osteoclast recruitment is probably central to diseases where the bone is eroded, such as rheumatoid arthritis (Wong and others 2008). TNF-α is expressed in the periodontium on the pressure side during experimental orthodontic tooth movement in vivo (Yoshimatsu and others 2006), but IFN-γ has been found to inhibit TNF-α-induced osteoclastogenesis in vivo and in vitro (Kohara and others 2011). In this study, IFN-γ inhibited osteoclastogenesis during orthodontic tooth movement, suggesting that the experimental tooth movement may cause TNF-α-induced osteoclastogenesis that is then inhibited by IFN-γ.
Our results lead us to conclude that IFN-γ induction is able to inhibit mechanical force-loaded osteoclastogenesis and tooth movement.
Footnotes
Acknowledgments
This work was supported by a Grant for Scientific Research from the Ministry of Education, Science and Culture, Japan, and the President's discretionary fund of Nagasaki University.
Author Disclosure Statement
No competing financial interests exist.