
Abstract
Toll-like receptors (TLRs) play an important role in the induction of host responses to pathogens. Interactions between TLRs and their ligands result in the production of cytokines that modulate the adaptive immune response through polarizing CD4+ T cells into either T-helper (TH)1 or TH2 phenotypes. In this regard, TLR2 and TLR5 ligands have been shown to induce responses in mammals that are biased toward TH1 or TH2 phenotypes. However, whether a similar phenomenon occurs in chickens remains to be elucidated. To this end, chicken splenocytes were stimulated with the TLR2 ligand Pam3CSK4 and the TLR5 ligand flagellin, and the relative expression of several cytokines and transcription factors was quantified at 1, 3, 8, and 18 h poststimulation. The results suggest that both TLR ligands induce a mixed TH1- and TH2-like response, as characterized by the upregulation of both the TH1-associated cytokine interferon-γ and the TH1-inducing cytokine interleukin (IL)-12, in addition to the TH2-associated cytokine IL-4, and in the case of flagellin, IL-13 as well. Future studies may be aimed at assessing the adjuvant potential of these ligands.
Introduction
T
Interactions between TLRs and their ligands typically result in cellular activation and the production of cytokines. These cytokines include proinflammatory cytokines, which help to establish the early innate response, as well as cytokines that modulate the adaptive immune response through polarizing CD4+ T cells into either T helper (TH) 1 or TH2 phenotypes. In mammals, certain TLRs, such as TLR 7 and 9, induce a TH1-biased response characterized by the production of interferon (IFN)-γ and interleukin (IL)-12 (Netea and others 2005). In contrast, TLR 2 and 5 may induce either a TH1- (Mazzoni and Segal 2004; Vicente-Suarez and others 2009) or a TH2-biased response, as indicated by the upregulation of IL-4, IL-5, and IL-13 (Didierlaurent and others 2004; Dillon and others 2004).
In chickens, the immune-modulating effects of several of the TLRs have been characterized, including TLR3, 4, 7, and 21 (Patel and others 2008; Jenkins and others 2009; St. Paul and others 2011; Villanueva and others 2011). However, few studies have examined the kinetics of responses mediated by TLR2 and 5. It has been established that chicken TLR2 and TLR5 share a similar ligand specificity to their mammalian counterparts (Fukui and others 2001; Keestra and others 2007, 2008). Moreover, TLR2 and 5 have been detected in several cell subsets of chickens, including in heterophils, where they mediate oxidative burst and the production of cytokines (Kogut and others 2005, 2007; He and others 2006). However, the influence of TLR2 and 5 on induction of chicken responses has yet to be elucidated in chickens. The objective of the present study was to determine whether stimulation of chicken TLR2 and 5 could lead to the induction of genes associated with TH1- or TH2-like responses in chickens. Through transcriptional analysis of relevant cytokines and transcription factors, the results of the present study suggest that both TLR ligands can induce a mixed TH1- and TH2-like response.
Materials and Methods
Chickens
Four-week-old broiler chickens (n=6) were procured from the Arkell Poultry Research Center, University of Guelph (Guelph, ON). The chickens were raised in a large cage without any litter. This research was approved by the University of Guelph Animal Care Committee and complied with the guidelines of the Canadian Council on Animal Care.
TLR ligands
The synthetic triacylated lipopeptide Pam3CSK4 (Pam3) and purified flagellin (Fla) isolated from Bacillus subtilis were purchased from Invivogen (San Diego, CA). Both of the ligands were resuspended in sterile endotoxin-free water and diluted to working concentrations in a complete RPMI medium.
Splenocyte isolation and stimulation
Spleens were collected from 6 chickens and subsequently minced and filtered through a 40-μm nylon cell strainer to obtain a single-cell suspension. The suspension was overlaid onto a Histopaque-1077 (Sigma-Aldrich, Oakville, ON) gradient and centrifuged at 400 g for 30 min, and the mononuclear splenocytes were harvested from the plasma–Histopaque interface and washed 3 times in RPMI-1640 (Invitrogen, Burlington, ON) supplemented with 10% heat-inactivated fetal bovine serum, 200 U/mL penicillin, 80 μg/mL streptomycin, 25 mg gentamicin, 10 mM HEPES buffer, 50 μM β-mercaptoethanol, and 2 mM
RNA extraction and cDNA synthesis
Total RNA was extracted from splenocytes using TRIzol® (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol and treated with DNA Free® (Ambion, Austin, TX) DNAse. Subsequently, 500 ng of purified RNA was reverse transcribed to cDNA using the Superscript® II First Strand Synthesis kit (Invitrogen) and oligo-dT primers, according to the manufacturer's recommended protocol. The resulting cDNA was subsequently diluted 1:10 in DEPC-treated water.
Real-time polymerase chain reaction
Real-time reverse transcription–polymerase chain reaction (RT-PCR) was used to measure transcripts for cytokines and other molecules associated with TH1 or TH2 responses. Since little is known about the phenomenon of TH1/TH2 polarization in chickens, and given the fact that we did not determine the cellular source of cytokines produced, hereafter, we refer to the observed cytokine expression results as indicatives of TH1-like or TH2-like responses. Quantitative real-time PCR using SYBR Green was performed on diluted cDNA using LightCycler® 480 II (Roche Diagnostics GmbH, Mannheim, Germany) as previously described (Villanueva and others 2011). Briefly, each reaction involved a preincubation step at 95°C for 10 min, followed by 45 cycles of 95°C for 10 min, 55°C–64°C (TA as per primer) for 5 s, and elongation at 72°C for 10 s. Subsequent melt curve analysis was performed by heating to 95°C for 10 s, cooling to 65°C for 1 min, and heating to 97°C. Primers were synthesized by Sigma-Aldrich-Canada (Oakville, ON), and their specific sequences and accession numbers are outlined in Table 1.
IL, interleukin; IFN, interferon; Stat4, signal transducer and activator of transcription 4; Gata3, GATA-binding protein 3; MyD88, myeloid differentiation factor 88; F, forward; R, reverse.
Data analysis
Relative expression levels of all genes were calculated relative to the housekeeping gene β-actin using LightCycler® 480 Software (Roche Diagnostics GmbH), based on the formula developed by Pfaffl (2001). Data represent mean±standard error. In each treatment or control group, there were 6 biological replicates. Statistical significance between treatment groups and the medium control group was calculated using a paired student's t-test and was considered statistically significant if *P≤0.05 and **P≤0.01.
Results
To assess the ability of flagellin and Pam3CSK4 to induce cytokines associated with TH1-like responses or their induction, we measured IFN-γ and IL-12 expression at various time points post-treatment. Treatment with both the low dose (0.1 μg/mL) and high dose (1.0 μg/mL) of Pam3CSK4 upregulated IFN-γ transcripts at 1 h poststimulation (P≤0.05) and continued to increase in a dose-dependent manner, with transcripts in the high-dose-treated group being significantly upregulated at both 8 and 18 h post-treatment (P≤0.05) (Fig. 1A). Treatment with the low dose (0.1 μg/mL) of flagellin significantly upregulated IFN-γ transcripts at 1 h poststimulation (P≤0.05), while treatment with the high dose (1.0 μg/mL) upregulated transcripts at 8 h poststimulation (P≤0.05) (Fig. 1A). Furthermore, treatment with the low dose of Pam3 significantly upregulated IL-12 transcripts at 1 and 3 h poststimulation (P≤0.01), while the high dose upregulated IL-12 transcripts at both 3 (P≤0.01) and 18 h (P≤0.05) poststimulation (Fig. 1B). With respect to flagellin, IL-12 transcripts were upregulated solely in response to the high-dose treatment at 8 h poststimulation (P≤0.05) (Fig. 1B).
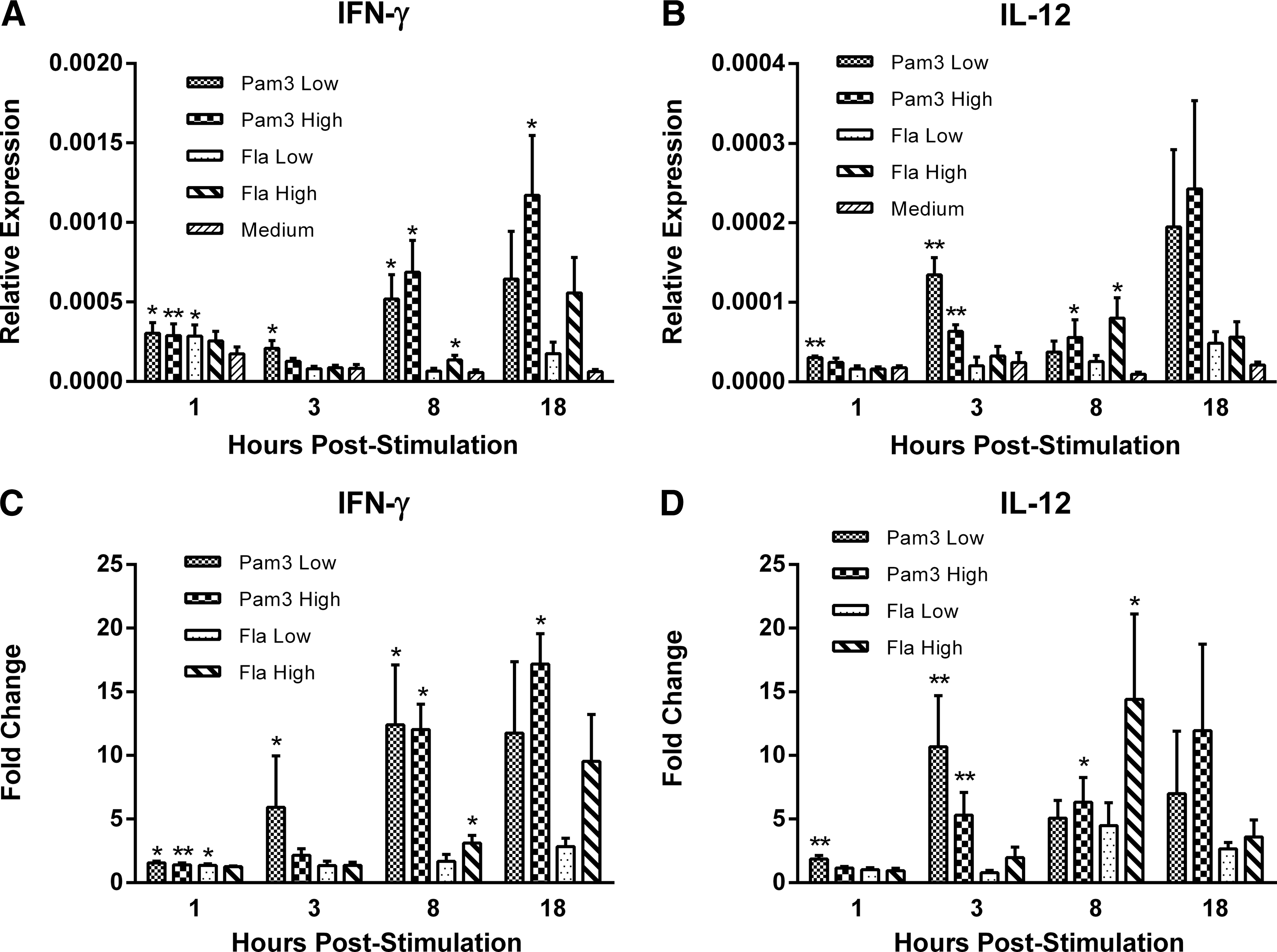
Relative gene expression and fold change of the T-helper 1 (TH1)-associated cytokine interferon (IFN)-γ and the TH1-inducing cytokine interleukin (IL)-12 in splenocytes isolated from 4-week-old broiler chickens at 1, 3, 8, and 18 h post-treatment with low (0.1 μg/mL) and high (1 μg/mL) doses of the toll-like receptor 2 (TLR2) ligand Pam3CSK4 and the TLR5 ligand flagellin. Data represent mean gene expression levels relative to the housekeeping gene β-actin±standard error
To gain a better understanding of the ability of TLR ligands to induce TH2-like responses, we measured expression of IL-4 and IL-13. Treatment with the high dose of Pam3 significantly downregulated IL-4 transcripts at 1 h poststimulation (P≤0.05), while upregulating them at 8 and 18 h poststimulation (P≤0.05) (Fig. 2A). Similarly, treatment with both the low and high dose of flagellin downregulated IL-4 transcripts at 1 h poststimulation (P≤0.05), while transcripts were upregulated at 18 h poststimulation for the high-dose-treated group (P≤0.05) (Fig. 2A). With respect to IL-13, although an upward trend was observed in response to Pam3, there were no statistically significant differences between any of the Pam3 doses and the medium control group at any of the sampling time points (Fig. 2B). This is in contrast to treatment with flagellin, in which IL-13 was initially downregulated at 1 h poststimulation, (P≤0.05), while subsequently being upregulated at both 8 and 18 h poststimulation (P≤0.05) (Fig. 2B).
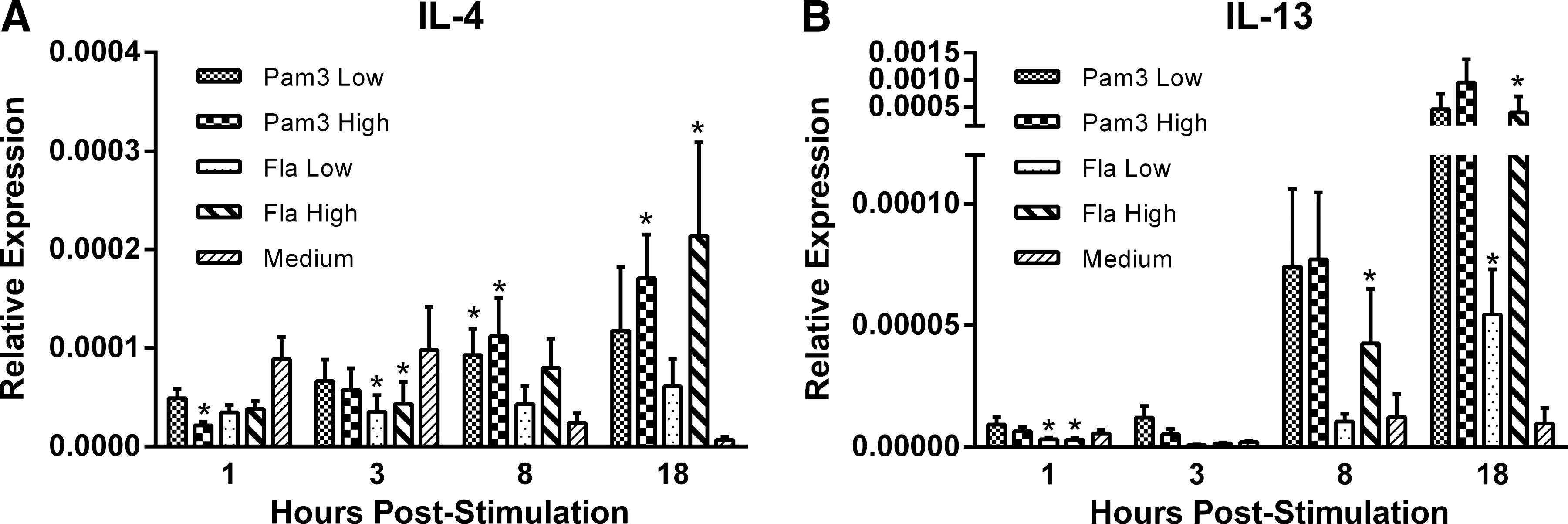
Relative gene expression of the TH2-associated cytokines IL-4
To gain further insight into the molecular mechanisms governing the responses to Pam3CSK4 and flagellin, gene expression was quantified for the transcription factors, signal transducer and activator of transcription 4 (Stat4), which is upregulated in response to IL-12, thereby suggestive of a TH1-biased response (Wurster and others 2000), and GATA-binding protein 3 (Gata3), which is considered to be a regulator of TH2 cell differentiation (Naito and others 2011), in addition to myeloid differentiation factor 88 (MyD88), which is an adaptor molecule critical to the TLR-signaling cascade (Yamamoto and others 2003). Treatment with both the low and high dose of Pam3 upregulated Stat4 at both 1 and 8 h poststimulation (P≤0.05), while transcripts were upregulated at 18 h post-treatment only in response to the high dose of Pam3 (P≤0.01) (Fig. 3A). Similarly, treatment with the low dose of flagellin upregulated Stat4 transcripts at 1 h poststimulation (P≤0.05), while treatment with the high dose of flagellin upregulated transcripts only at 18 h poststimulation (P≤0.05) (Fig. 3A). In contrast, transcripts for Gata3 were downregulated in response to treatment with either Pam3 or flagellin at both 1 and 3 h post-treatments (P≤0.05) (Fig. 3B). Transcript levels of MyD88 were significantly upregulated at all sampling time points in response to the high-dose Pam3 treatment (P≤0.05), while upregulated only at 8 h poststimulation with the low dose of Pam3 (P≤0.05) (Fig. 3C). However, MyD88 transcripts were upregulated only in response to the high dose of flagellin, and this occurred at both 1 and 3 h poststimulation (P≤0.05) (Fig. 3C).
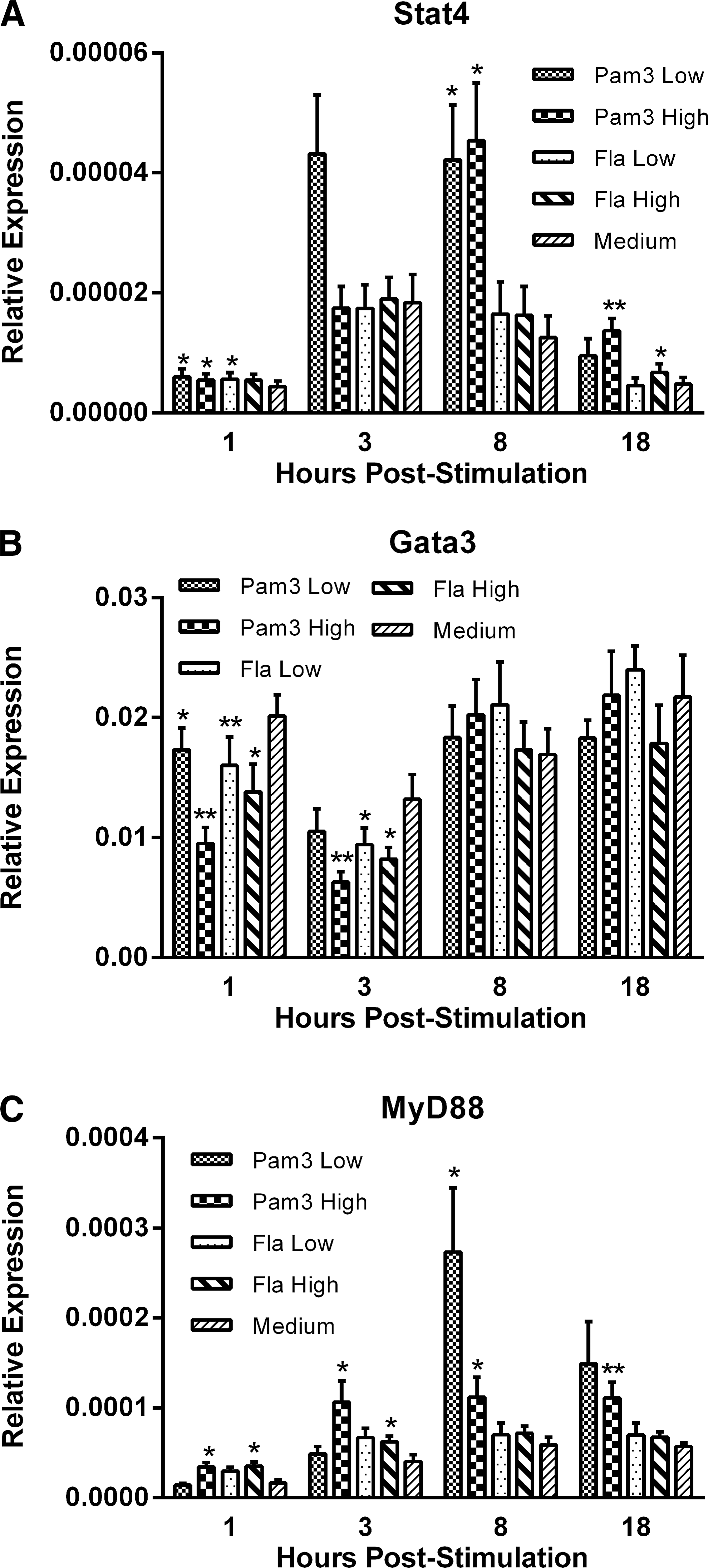
Relative gene expression of the transcription factors, signal transducer and activator of transcription 4 (Stat4)
Discussion
In mammals, several factors are known to influence the polarization of CD4+ T cells, including (1) the dose of the antigen, (2) the strength of the signal received by the T cell receptor (TCR), and (3) the milieu of cytokines present (Murphy and Reiner 2002). With respect to the latter, TLR ligands are known to modulate the composition of the cytokine milieu. For example, treatment with the TLR2 ligand Pam3CSK4 predominantly induces a TH2-biased response in mammals, which may be attributed in part to the TLR2-mediated production of IL-4, IL-5, and IL-13 by B cells (Agrawal and Gupta 2011) and mast cells (Varadaradjalou and others 2003). Indeed, Pam3 may induce a similar response in chickens, as suggested by the upregulation of IL-4 in the present study. However, in contrast to what has been observed in mammals, our results indicated that Pam3 upregulated IL-12, thereby suggesting the induction of a TH1-like response (Agrawal and others 2003). Therefore, we propose that treatment with Pam3 induces a mixed TH1- and TH2-like response in chickens. Interestingly, it has been shown in mice that different TLR2 ligands may differentially alter the kinetics of the host response as evident from the findings that Pam3 and lipoteichoic acids induce distinct proinflammatory responses (Long and others 2009). Studies are currently underway by our group to determine whether a similar phenomenon also occurs in chickens.
In contrast to Pam3CSK4, in vitro studies using mammalian cells have shown that flagellin polarizes the response toward a TH1 phenotype. For example, murine dendritic cells and human peripheral blood mononuclear cells stimulated with flagellin produced IL-12, but failed to upregulate IL-4 or IL-13 (Agrawal and others 2003; Bachmann and others 2006). Furthermore, flagellin was shown to activate human natural killer cells and enhance their IFN-γ production (Chalifour and others 2004). However, the results from the present study indicated a relatively small upregulation of IL-12 and IFN-γ in cells stimulated with flagellin, whereas IL-4 and IL-13 were highly upregulated in these cells. These observations suggest a mixed TH1-/TH2-like response in chicken cells stimulated with flagellin, perhaps with a TH2 bias. The reason for this discrepancy remains to be elucidated, and further studies are warranted. For example, production of IL-12 by chicken dendritic cells stimulated with flagellin may be examined. However, it is also important to note that when flagellin is used as a vaccine adjuvant in mammals, a TH2-biased response may be observed (Didierlaurent and others 2004; Bobat and others 2011). Based on this finding, future studies in chickens may consider exploring the adjuvant potential of flagellin to determine if the mixed TH1-/TH2-like response also occurs in this context.
The phenotype of a TH cell in mammals is defined not only by the cytokine profile it produces, but also by the presence of specific transcription factors, namely T-box transcription factor 21 (T-bet) for TH1 cells and Gata3 for TH2 cells (Naito and others 2011). Unlike Gata3, which has been identified and cloned in chickens (Ishihara and others 1995), to our knowledge, the chicken T-bet sequence has not been published. Therefore, to gain insight into the molecular mechanisms governing the TH1-like response, we quantified the transcript levels of Stat4, which is upregulated in response to IL-12 and is required by TH1 cells to adopt the full TH1 phenotype (Wurster and others 2000; Thieu and others 2008). To this end, only Pam3 strongly induced the upregulation of Stat4 transcripts, thereby supporting the notion of a TH1-like response. Furthermore, although we detected an upregulation of IL-4 and IL-13, suggesting a TH2-like response as well, we observed a downregulation of Gata3 transcripts. While the reason behind this is not known, one possible explanation is that in the absence of TCR signaling, IL-4 is required to upregulate Gata3 in mammals (Ho and others 2009). This is supported by the results from the present study, as the downregulation of IL-4 in the early phases of the response to both TLR ligands correlates with the downregulation of Gata3. In the present study, IL-4 transcripts peaked for both TLR ligand treatments at 18 h post-treatment. Therefore, Gata3 expression peak might have occurred after the 18 h time point. Indeed, a previous timecourse study in mice revealed that Gata3 transcripts were not upregulated until several hours after the peak in IL-4 transcripts, likely corresponding with the peak of IL-4 production at the protein level (Yamane and others 2005). In addition, another potential explanation may be due to the production of transforming growth factor-β by regulatory T cells, as this has been shown to downregulate Gata3 expression in mice (Kim and others 2010).
Although in the present study we have demonstrated that both ligands induced a mixed TH1-/TH2-like response, it is also important to consider many factors that may influence this response. For example, as we did not sample past 18 h poststimulation, it is possible that the immune system has not yet committed to a specific phenotype as the adaptive immune response is on its way to polarization. Furthermore, responses to the TLR2 and TLR5 ligands may have been influenced by previous exposure to environmental PAMPs during the growth of the chicken that may either sensitize or desensitize splenocytes to Pam3 or flagellin. In addition, the results from the present study also indicate a large expression of Gata3 in all samples, including in the medium control, which may be a reflection of previous activation and potential skewing toward a TH2 bias. However, it is important to note that preventing any previous exposure to PAMPs and antigen is unrealistic, as PAMPs found in dust may alter immune response, weight gain, and heart morphology (Lai and others 2009). In addition to environmental PAMPs, the diet of the chicken may also influence antibody responses to vaccination (Parmentier and others 2002) as well as cytokine responses to TLR ligands, such as LPS (Sijben and others 2003). Taken together, future studies maybe aimed at determining whether diet or environmental PAMPs may affect responses to TLR2 and TLR5 ligands.
In conclusion, we have demonstrated that the TLR2 ligand Pam3CSK4 and the TLR5 ligand flagellin induce a mixed TH1-/TH2-like response in chicken splenocytes. Future studies may be aimed at exploring their adjuvant potential.
Footnotes
Acknowledgments
Michael St. Paul is a recipient of an Ontario Graduate Scholarship in Science and Technology. This work was funded by the Natural Sciences and Engineering Research Council of Canada, Poultry Industry Council, and OMAFRA. The authors would like to thank Hamid Haghighi and Leah Read for their assistance.
Author Disclosure Statement
The authors declare no competing financial interests exist.