
Abstract
It is well known that ionizing radiations induce a marked downregulation of antigen-dependent and natural immunity for a prolonged period of time. This is due, at least in part, to radiation-induced apoptosis of different lymphocyte subpopulations, including natural killer (NK) cells. Aim of this study was to investigate the capability of Beta Interferon (β-IFN) and Interleukin-2 (IL2), alone or in combination, to restore the functional activity of the natural immune system. Mononuclear cells (MNCs) obtained from intact or in vitro irradiated human peripheral blood were treated in vitro with β-IFN immediately before or at the end of the 4-day treatment with IL2. Time-course analysis was performed on the NK activity, the total number and the apoptotic fraction of CD16+ and CD56+ cells, the 2 main NK effector cell subpopulations. The results indicate that radiation-induced impairment of natural cytotoxicity of MNC could be successfully antagonized by the β-IFN+IL2 combination, mainly when exposure to β-IFN preceded IL2 treatment. This radioprotective effect is paralleled by lower levels of radiation-induced apoptosis and increased expression of the antiapoptotic Bcl-2 protein. Since natural immunity can play a significant role in antitumor host's resistance, these results could provide the rational basis for a cytokine-based pharmacological strategy able to restore immune responsiveness and to afford possible therapeutic benefits in cancer patients undergoing radiotherapy.
Introduction
Different processes appear to be involved in the cellular response to IR, for example, generation and scavenging of free radicals, DNA duplication and repair, apoptosis, and inflammation (Sharma and others 2010; Thoms and Bristow 2010). In particular, repair of DNA damage after IR is believed to be a major protective mechanism since nuclear DNA is the most susceptible target in living cells (Wu and others 2011).
Two main compartments of the immune system, that is, antigen-dependent (adaptive) and antigen-independent (innate or natural) immunity are endowed with different, but frequently integrated functions in host's defenses against foreign invaders or malignant cells. Natural Killer (NK) cells play a major role in innate immunity by killing target cells directly or indirectly by releasing a variety of cytokines and chemokines. NK cells lyse a wide range of tumor cell types without presensitization and restriction by the major histocompatibility complex, do not express T-cell antigen receptors, CD3 T cell markers, or surface immunoglobulin B cell receptors. They are phenotypically characterized as CD56+ (a surface antigen present at low or at high levels) and CD16+ (FcγRIII receptor), and constitutively express p75 chain of the Interleukin-2 (IL2) receptor (De Maria and others 2011; Vivier and others 2011). The incubation of peripheral blood lymphocytes with IL2 induces rapidly a selective proliferation and increase of NK activity (Dunne and others 2001) followed by generation and proliferation of an activated NK cell population [i.e., lymphokine-activated killer (LAK) cells]. For their immunological characteristics, NK and LAK cells appear to play a notable role in antiviral response and tumor control (Sinkovics and Horwath 2005; Fuggetta and others 2008; Suck and Koh 2010). Radiotherapy, which is widely utilized in cancer treatment causes severe damage to hematopoietic and lymphoid tissues and results in a marked immunodepression for a prolonged period of time (Ashwell and others 1988; Field and Becker 1989; Park and others 2011). In particular, it is well documented that γ-radiations impair host's NK activity in a dose-dependent manner (Hochman and others 1978; Blomgren and others 1980; Blomgren and others 1982; Zarcone and others 1989). In addition, decline of NK activity and decrease in the percentage of NK cells in the peripheral blood has been found in cancer patients receiving radiotherapy (McGinnes and others 1987).
Previous reports have shown that the exposure of mononuclear cells (MNCs) to IR is followed by a time-dependent decrease of NK activity in vitro (Zarcone and others 1989) and a marked induction of apoptosis (Delic and others 1993). The decline of NK activity was found to be correlated with a reduction of the number of CD16- and CD56-positive cells and of the percentage of the binding of effector cells to their targets (Fuggetta and others 1998).
Moreover, it has been recently demonstrated that exposure to IR induces long-term expression of senescence markers in vivo, thus showing that irradiation can be followed by persistent damage to different host's tissues, including components of the immune system (Le and others 2010). This is consistent with the commonly accepted opinion that radiotherapy downregulates immune responses. Surprisingly however, little information is presently available on possible pharmacological strategies able to restore immune responsiveness in irradiated subjects.
Among biological response modifiers, interferons (IFNs) represent a group of cytokines with a long history as immunotherapeutic agents employed in the treatment of various solid tumors and hematological malignancies. The use of IFNs in cancer therapy was based on their antiproliferative and immunomodulatory effects. More recently, IFNs have been found to possess cytotoxic and antiangiogenic properties (Bracarda and others 2010; Commins and others 2010; Moschella and others 2010). Therefore, combined use of radiotherapy and IFNs could be considered of potential value in the management of various types of neoplastic diseases. Actually, preclinical studies in vivo and in vitro have demonstrated that IFN modifies the radiation effects, enhancing the killing activity of IR on tumor cells and increasing immune responsiveness (Ortaldo and Mc Coy 1980). In particular, it is well known that IFNs are potent activators of NK cell function in vitro and in vivo (Silva and others 1980).
In previous studies, we have demonstrated that in vitro treatment with beta IFN (β
Materials and Methods
Cell lines
The NK- and LAK-susceptible erythroleukemia cell line, K562, and the NK-resistant and LAK-sensitive Burkitt lymphoma cell line, DAUDI, were obtained from ATCC. The EBV-transformed NK-resistant and LAK-sensitive B lymphoblastoid cell line BSM was described by Christmas and Moore (Christmas and Moore 1987). All cell lines were maintained in suspension culture in RPMI-1640 containing 10% fetal calf serum (FCS; Gibco Lab.) and 2 mM l-Glutamine (Flow Laboratories) hereafter referred to as the “Complete Medium” (CM) and subcultured 3 times weekly.
Irradiation of peripheral blood and separation of MNCs
Peripheral blood obtained from healthy adult volunteers after signed consent, and admixed with ethylenediaminetetraacetic acid to prevent coagulation, was exposed in vitro to 20 Gy using a Cesium-137 irradiator (Gamma Cell 1000; mod A, AECL).
MNCs from intact or irradiated peripheral blood were separated on a Ficoll-Hypaque gradient (Pharmacia) and hereafter called MNC and MNC/ir, respectively, as illustrated in Table 1. MNC and MNC/ir collected from the interface were washed twice with RPMI-1640 (Gibco Lab.), resuspended, and cultured in the CM.
Day 0 is considered the day of experiment start.
WB, whole blood. Anticoagulated whole blood was irradiated before MNC separation.
LAK/ir (treatment code TC-4) indicates LAK cells from nonirradiated MNCs, subjected to 20 Gy γ-irradiation. These cells were utilized for cytotoxicity assay within 2 h after irradiation.
Test cells were treated with β
Test cells were treated with IL2 from day 0 through day 4, and then treated with β
LAK, lymphokine-activated killer; MNC, mononuclear cell.
Cytokines
Recombinant human IL2 was kindly provided by Roche, and natural fibroblast β
Treatment of MNC with cytokines alone or in combination
All in vitro treatment schedules utilized in the present study are summarized in Table 1.
On day 0, MNC and MNC/ir were suspended at the final concentration of 2×106 cells/ml and cultured in the CM alone or in the presence of 500 UI/mL of IL2 at 37°C in a 5% CO2 humidified atmosphere for further 4 days. IL2-stimulated effector cells (called hereafter LAK and MNC/ir/IL2, respectively, as reported in Table 1, under TC-2 and TC-3 code, respectively) were recovered, counted, washed twice, and used for cytotoxicity assays and cytofluorimetric analysis.
Moreover, LAK cells obtained from intact MNCs were suspended in the CM at the concentration of 2×106 cells/ml and irradiated with 20 Gy. Shortly after irradiation, LAK cells, called hereafter as LAK/ir (Table 1, TC-4) were used as effector cells against DAUDI, BSM, and K562 cell lines in a standard 51Cr-release assay.
Combined treatments with β
Cell enumeration and viability of all MNCs treated (see Table 1) has been performed by trypan blue dye exclusion.
Cytotoxicity assay
The cytotoxicity activity of effector cells was determined using a standard 51Cr-release assay (Alvino and others 1999). Experimental results were expressed in terms of % specific lysis, or in terms of number of target cells lysed (i.e., killed) by a fixed number of effector cells, as follows:
were KC(m) is the number of target cells killed by m effector cells; T is the number of target cells present in each well (i.e., 2×103 cells); En is the number of effector cells present in each well at the selected E:T ratio, and n% is the specific lysis produced theoretically by En effector cells. The n% is extrapolated from the best fit curve obtained by plotting the different percentages of specific lysis versus ln of the number effector cell/well. In the present study, KC(m) was calculated for m=106 [i.e., KC (106)] and En=2×105 cells (i.e., E:T cell ratio of 100:1) (Alvino and others 1999).
Immunofluorescence staining and cytometric analysis
The surface phenotypes of MNC and MNC/ir, nontreated or incubated with β
MNC/irr exposed to the 2 cytokines were also tested for Bcl-2 expression. Cultured cells were washed twice in PBS, supplemented with 0.1% bovine serum albumin, and 0.02% sodium azide (PBS-A). Cells (1×106) were then fixed in 1% formaldehyde in PBS at 4°C for 15 min and permeabilized in 70% ethanol at −20°C for 1 h. Cells were then suspended in 50 μL of PBS-A containing mAb recognizing Bcl-2, clone 124 (DAKO A/S, Denmark), washed, and labeled with rabbit anti-mouse immunoglobulins, F(ab')2- FITC (DAKO, code F0313). FITC-conjugated mouse IgG2 was used as a negative control. Analysis was performed using a FACScan flow cytometer (Becton-Dickinson).
Flow cytometry was also used to detect apoptosis in MNCs and MNC/ir (Nicoletti and others 1991), nontreated or incubated with the cytokines. Cells were then washed in PBS and fixed with 50% acetone:methanol 1:4 in PBS at 4°C for at least 18 h. After 2 washes in PBS, the cells were suspended in PBS (1×106cells/mL) containing 50 μg/mL of propidium iodide (PI; Sigma) and 100 KU/mL of RNase A (Sigma) and incubated in the dark at room temperature for 30 min. The DNA content per cell was evaluated by PI fluorescence measured on a linear scale using the FACScan flow cytometer (Becton-Dickinson) with an argon ion laser emitting at 488 nm. Data collection of 10,000 cells/sample was gated utilizing forward light scatter and side light scatter to exclude cell debris and aggregates. All data were recorded using CellQuest software (Becton-Dickinson). Apoptotic cells were determined by their hypochromic, sub-G1 staining profiles (Nicoletti and others 1991).
The effect of the 2 cytokines on the production of γ
Western blot analysis
MNCs obtained from whole blood irradiated with 20 Gy (i.e., MNC/ir) were exposed to the cytokines under investigation for 24 h, washed in PBS, centrifuged, and pelleted. The cell pellets were suspended in the lysis buffer (50 mM Tris-HCl ph 7.5, 1 mM phenyl methylsulfoniyl peroxide-2 mM EGTA-10mMDTT-5 mg/mL of aprotinin, 200 mg/mL of leupeptin-0.1%-Triton X-100) kept on ice for 30 min, sonicated 5 s at 4°C, and centrifuged for 5 min at 15,000 g at 4°C in an Eppendorf microcentrifuge. All reagents were obtained from Sigma. Protein concentrations were determined using the Bio-Rad protein reagent with bovine serum albumin as the standard protein. Thereafter, proteins were separated on 12% SDS(w/v) polyacrylamide gel and transferred to a nitrocellulose filter, using a Biorad electrophoretic blotting apparatus (BioRad). Transfer was carried out at 25 V, at 4°C for 14 h. After transfer, membranes were incubated with 3% (w/v) dry milk (Bio Rad) in TBS (20 mM Tris -HCl ph7.5; 0.9 NaCl) with gentle agitation for 1 h. The membrane was then incubated at room temperature with mouse anti-human Bcl-2 (DAKO, 1:1000) in TBS containing 0.05% Tween 20 (TBST) for 2 h. Thereafter, the membrane was washed with TBST and incubated with the alkaline phosphatase-coupled secondary antibody diluted 1:7500 in TBST for 1 h. The band was visualized using the Photobed (Promega Biotec) reagents, according to the procedures provided by the manufacturer.
Statistical analysis
Differences in cytolytic effects produced by effector cells in various experimental conditions were evaluated taking into account the percentage of specific cytotoxicity at all E/T ratios. Therefore, p-values were calculated using covariance analysis performed on the regression of the percentage of specific 51Cr release over the ln of the number of effector cells/well. All data relative to cell-mediated cytolysis are expressed in terms of mean KC(106) values without conventional standard error (SE) or standard deviation of the mean. Actually, no statistical analysis can be performed using SE or standard deviation, which are not suitable for covariance analysis of regression lines (Alvino and others 1999).
Where the results are expressed in terms of means±SE, statistical significance was determined using the Student's t-test analysis.
Results
Effect of IL-2 and β-IFN on the functional activity and number of cells exposed to γ-rays
Control MNCs or MNC/ir obtained from peripheral blood exposed to γ-rays, were treated with 500 IU/mL of IL-2 for 4 days (LAK generation) and tested for cytotoxicity against DAUDI, BSM, or K562 cell lines. The results of a representative experiment (Fig. 1) expressed in terms of percent-specific lysis or of KC(106) (see legend of Fig. 1), show that pre-exposure to γ-rays reduces significantly the cytotoxic activity of effector cells. This inhibition, ranging from 40% to 75%, was detectable both against a NK-resistant DAUDI and BSM target cell lines, and against NK-susceptible K562 target cells. We also examined whether similar effects could be obtained when the γ-ray exposure was performed after LAK generation, that is, after IL2 treatment. When MNCs were first treated with IL2 for 4 days and then irradiated with γ-rays (i.e., LAK/ir), they displayed a reduced cytotoxic activity against DAUDI and BSM, but retained almost entirely the lytic activity against the K562 cell line (Fig. 1B).
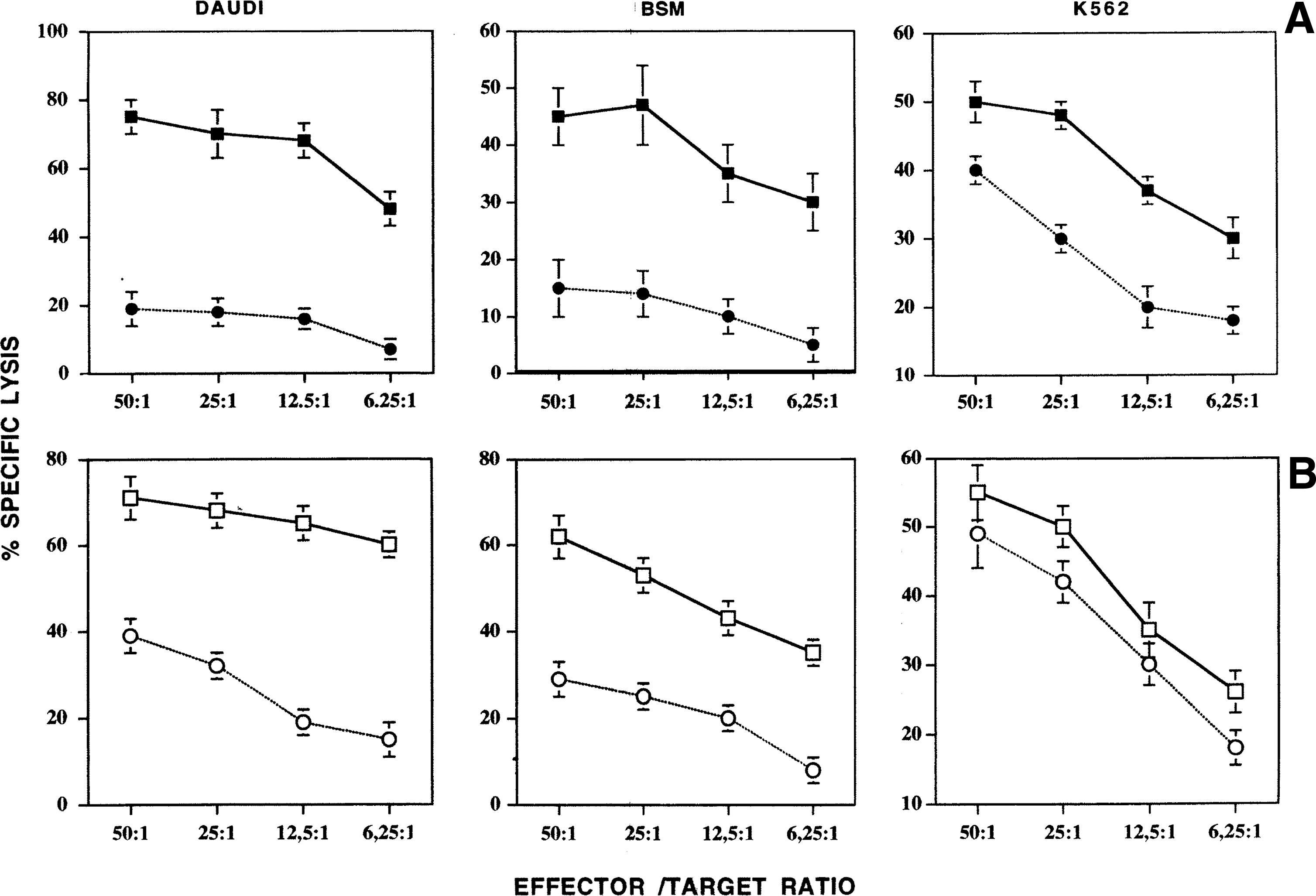
Cytotoxic activity of lymphokine-activated killer (LAK) cells against 3 different target cells. All results are expressed in terms of % specific lysis at different effector/target ratios. Bars represent the SE of the mean. The data relative to
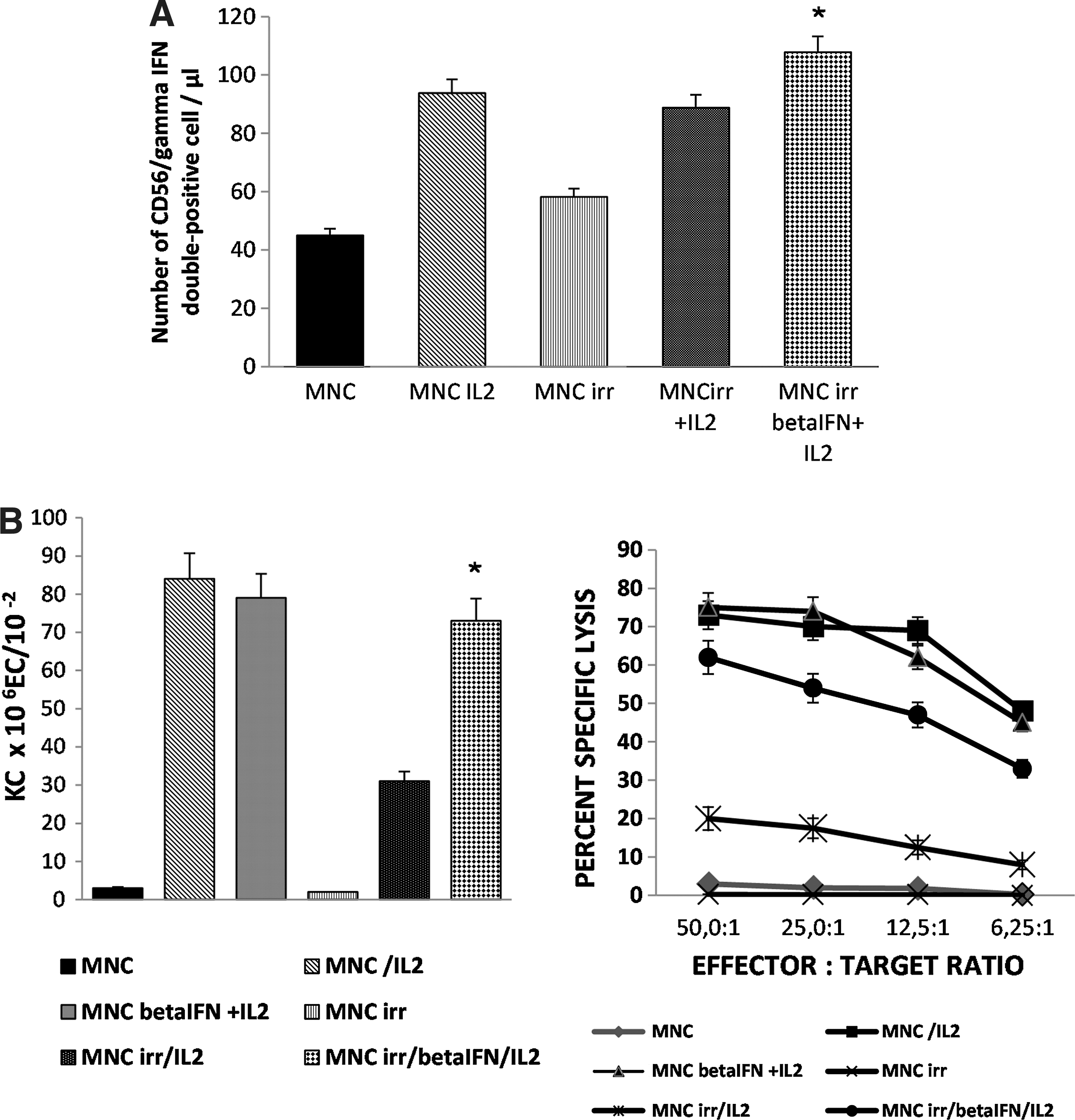
To determine the ability of β
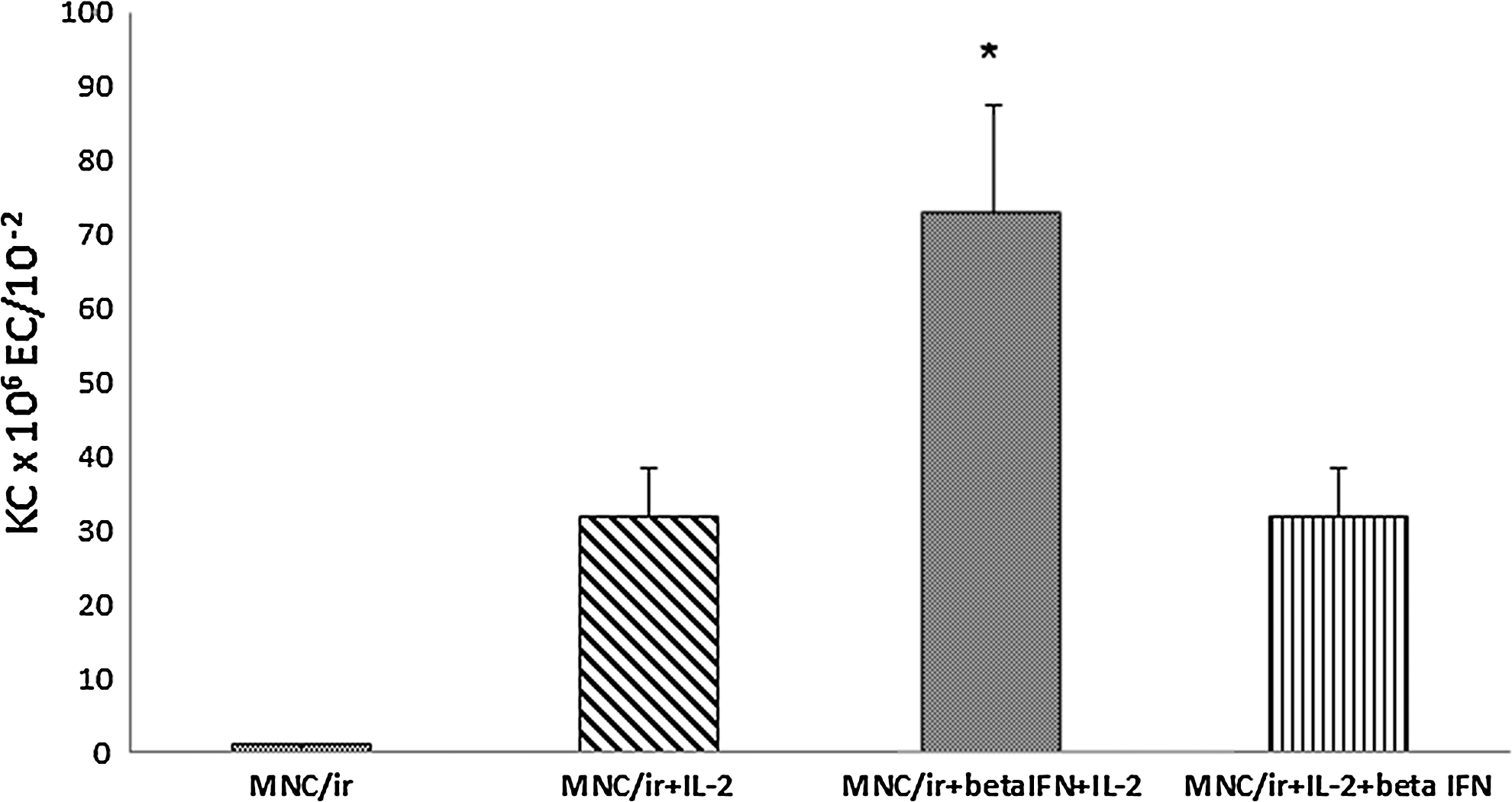
Effect of β
Secretion of γ
Viable (trypan blue exclusion) cell count of intact or irradiated MNCs, nontreated or subjected to cytokine treatment, was performed on day 1 and 4 of culture (Fig. 4). It was found that: a) IR induce a step and progressive decline in the number of MNCs; b) treatment of MNC/ir with IL2 alone did not antagonize the effects of γ-rays on the total number of cells tested on day 1; c) treatment with β
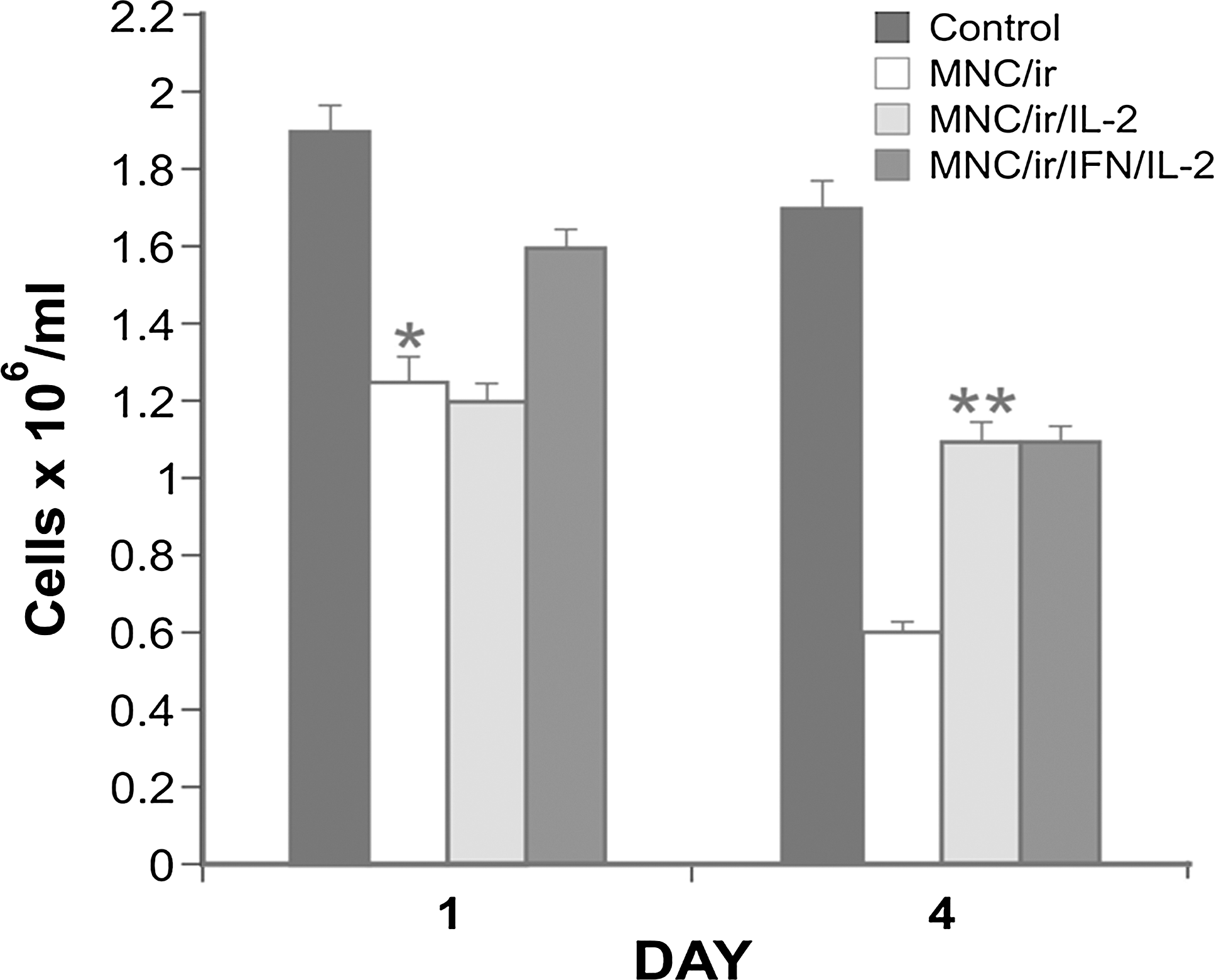
Effect of IFN and IL2, alone or in combination, on the cell number of MNCs collected from intact or irradiated (20 Gy) peripheral blood (treatment schedules are described in Table 1). Results of a representative experiment, that are expressed in terms of geometric mean (each group in triplicate) of viable cell counts. Bars represent the SE of the geometric mean. (*) P<0.05, number of viable cells of MNC/ir versus MNC/ir/IFN/IL2 on day 1. (**) P<0.05, number of viable cells of MNC/ir versus MNC/ir/IL2 and MNC/ir versus on day 4.
Effect of IL2 and β-IFN on γ-ray induced apoptosis and Bcl-2 expression
It is well known that γ-rays can induce apoptotic death in human lymphocytes (Belloni and others 2005; Sharma and others 2010). Therefore, MNC/ir were treated with IL2 for 4 days alone or in combination with β
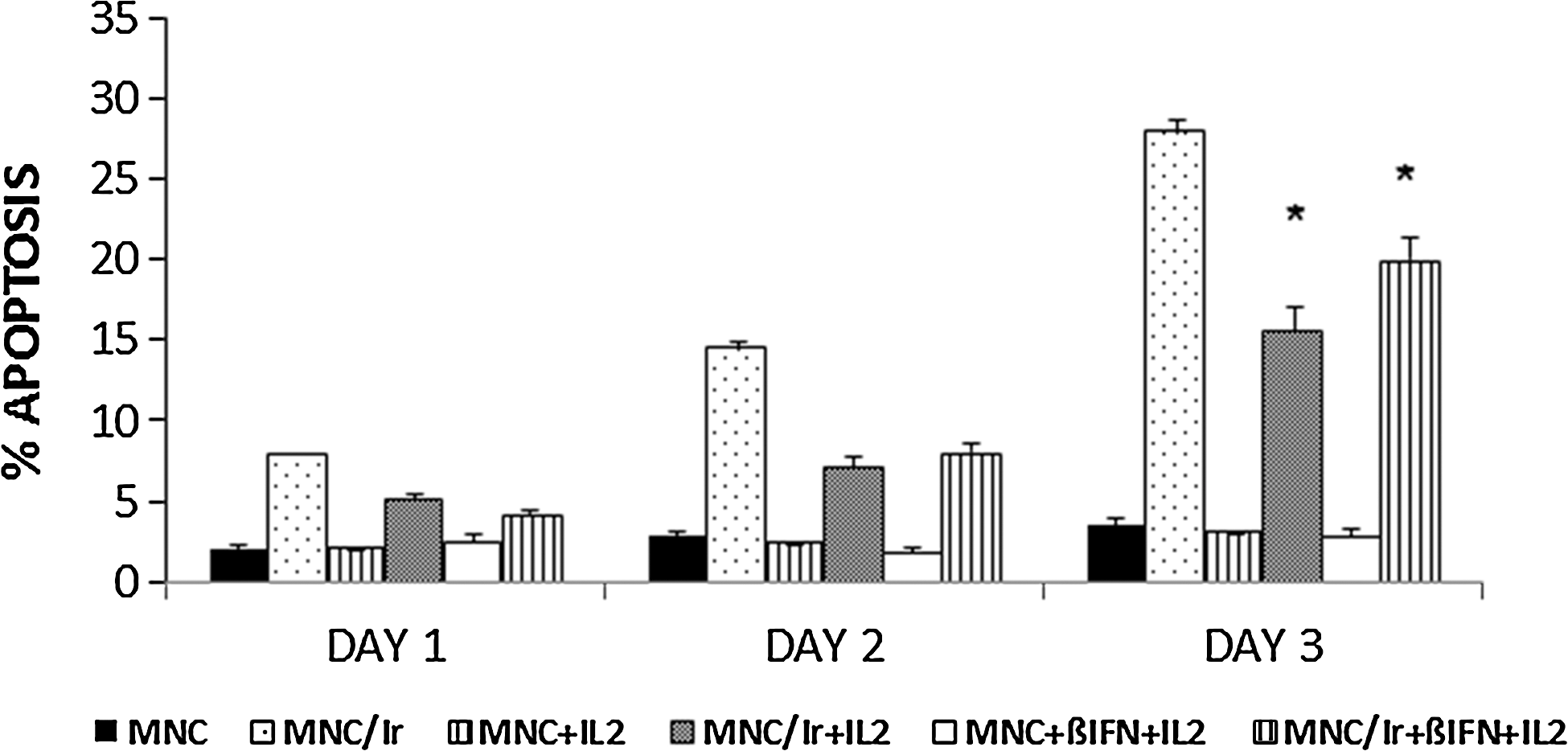
Time-course of apoptosis induction in cultured MNCs collected from intact or from irradiated (20 Gy) blood, and treated with β
Large experimental evidence is now available showing that overexpression of the oncogene Bcl-2 is able to inhibit apoptosis induced by a variety of stimuli. Therefore, Bcl-2 expression was examined to establish whether it could be involved in the rescue effect produced by the combination of β
The levels of the Bcl-2 protein were analyzed in irradiated cells after 24 h of exposure to β
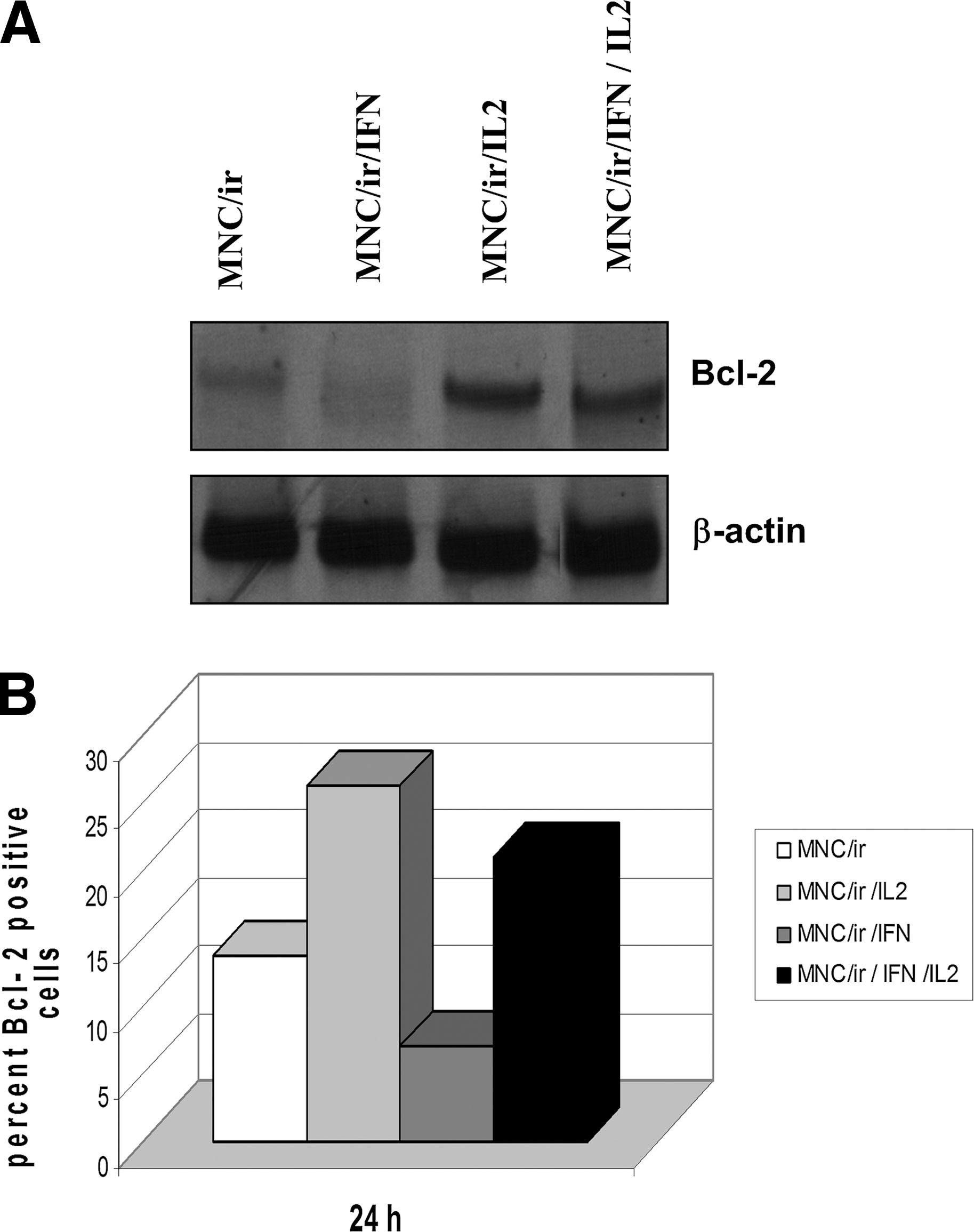
MNC obtained from irradiated (20 Gy) blood, were cultured with medium alone (MNC/ir), with β
Time-course of the number of CD56+ and CD16+ lymphocyte subpopulations after exposure to γ-rays and incubation with the cytokines under investigation
The phenotypes of MNC, untreated or incubated with IL2 (i.e., LAK cells), of MNC/ir, untreated or exposed to IL-2 alone (Table 1, TC-3), or incubated with β
On day 0, MNC were seeded in 24-well plates at the concentration of 2×106 cells/mL, 2 mL/well.
Isolated MNCs or MNC/ir (2×106 cells/mL) were cultured in vitro from day 0 through day 4. Treatment with IL2 alone or combined with β
M (SE), mean±standard error (3 observations/group).
For abbreviation identification, see Table 1.
No statistically significant difference was detected between the mean number of CD56+ cells relative to MNC/ir/IL2 group and that relative to MNC/ir/IFN/IL2 group on day 1 or on day 4. Similar results were obtained with CD16+ cells.
%, percent of MNC-positive for the relative marker evaluated by flow cytometry; NT, not tested.
Discussion
The present study has been designed to investigate the potential role of β
In earlier studies (Fuggetta and others 1989, 1998), we found that β
To achieve a more prolonged protective effect against γ-rays, we investigated the effects of β
Several molecules have been involved in the effector function of activated NK cells. Target cell killing is achieved through directed release of lytic granules comprising Granzyme B and perforin [Bryceson and others 2011). Moreover, recognition of target cells is accompanied by the production of chemokines and cytokines, including γ
As mentioned before, time-course analysis of cultured MNCs obtained from irradiated blood shows that rapid decay of the mean number of cells occurred within 4 days of cell culture (Fig. 4). A substantial rescue of cell numbers was found on day 4 in groups treated with IL2 or β
The current literature concerning the effect of IL2 and IFNs in combination with IR on the natural immune system is very limited. Hietanen and others (1995) evaluated in vitro the modulating effect of IFN alpha, beta, and gamma as well as of IL2 on the radiosensitivity of large granular lymphocytes endowed with the NK cell activity. These authors showed that no combination of any type of IFN and IL2 was more radioprotective than IL2 used alone (Hietanen and others 1990, 1995). The reason of the discrepancy between our results and Hietanen's data may be due to the different time intervals elapsing between cell irradiation and cytotoxicity assay as well as to different IL2 and β
In human blood, NK cells can be subdivided into different populations based mainly on the relative expression of the surface markers, CD16 and CD56, which show different functional properties. The CD56+ subset is more immunoregulatory, has little cytolytic activity, but releases high levels of cytokines, whereas the CD16+ subset, which predominates in peripheral blood and inflamed tissues, is characterized by a lower cytokine production, but displays high cytotoxic effector function (Moretta and others 2008; Maghazachi 2010). It has been demonstrated that NK cells belong to highly radiosensitive lymphocyte subpopulation. On the contrary, B and CD8+ T cells show a weaker susceptibility to IR, and CD4+ T cells are relatively radioresistant (Rana and others 1990). However, the radiosensibility of NK cells remains still a matter of debate (Philippé and others 1997; Wilkins and others 2002; Vokurková and others 2006).
In a previous study, we confirmed that decrease of the NK activity is time dependent and could be associated to a significant reduction of the number of CD16+ cells that appeared to be the most sensitive subpopulation to radiation damage (Fuggetta and others 1998). This has been confirmed by the results of the present investigation showing that in case of irradiated blood the number of CD16+ cells decreased severely, whereas a less pronounced cell loss occurred at the level of CD56+ lymphocyte subpopulation on day 4 of culture (Table 2). It must be pointed out that a minimal CD8+/CD56+ NKT cell population has been also indicated as a lymphocyte subset particularly susceptible to low doses of γ-rays (Vokurková and others 2010). In any case, the effects of irradiation on specific NK subpopulations are still an open question and the literature is controversial, and in most cases, available information has been obtained from clinical experimentation (Santin and others 2002; Eric and others 2009).
The mechanisms underlying radiation damage of NK cells are not entirely understood. Numerous studies showed that radiations compromise the integrity and the cytotoxic activity of NK cells both in vivo and in vitro and induce programmed cell death (Philippé and others 1997; Louagie and others 1998; Wilkins and others 2002; Sharma and others 2010). In line with these general observations, the results illustrated in Fig. 5 show that marked and progressive increase in the percentages of apoptotic cells can be found in MNC/ir up to 3 days postirradiation. Moreover, the same figure provides evidence that the treatment with IL2 alone or combined with β
Several authors have shown that IL2 antagonizes the detrimental effects of IR on viability and cytotoxicity of effector lymphocytes, depending on the IL2 concentration, radiation dose, and intervals between irradiation and observation. (Chong and others 1991; Seki and others 1995; Mor and Cohen 1996). This cytokine may play an important role in the regulation of lymphocyte survival in the control of the cell cycle during the immune response both in health and disease. In particular, expression of the Bcl-2 gene promotes cell survival by counteracting apoptosis stimuli, and the mechanism by which IL2 downregulates apoptosis of lymphocytes can be reasonably linked to the induction of Bcl-2 (Mor and Cohen 1996; Adachi and others 1998). Actually, evasion from DNA damage-induced cell death, via overexpression of prosurvival antiapoptotic Bcl-2 family proteins (Wyllie and others 2010) could be considered a key step toward radiation resistance. The results illustrated in Fig. 6 are in line with this hypothesis since Western Blotting and flow cytometry analysis show that enhanced expression of the Bcl-2 protein occurs in IL2-treated MNC/ir. When β
Of particular interest are the results of a preliminary study performed in our laboratory showing that treatment of human melanoma and breast cancer cells with β
In conclusion, we have demonstrated that β
Footnotes
Acknowledgment
This work was partially supported by MIUR Italy, PRIN 2008 n°20089E83YR_005 to MP Fuggetta.
Author Disclosure Statement
No competing financial interests exist.